| Formalisme du modèle allostérique MWC (1965) - 1ère partie |
| Tweet |
|
| Le cours qui suit est inspiré du livre "Enzyme Kinetics" de Irwin H. SEGEL (1993) (Wiley & sons. ed.) |
|
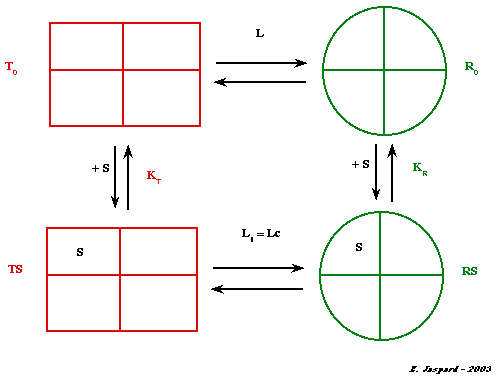
a. Équilibres macroscopiques : constantes Li, KT et KR Fixation de la 1ère molécule de substrat S
|
L T0 <===> R0 |
[T0] L = ----- [R0] |
| KT T0 + S <===> TS |
[T0]
[S] KT = -------- [TS] |
|
| L1 TS <===> RS |
[TS] L1 = ------ [RS] |
|
| KR R0 + S <===> RS |
[R0]
[S] KR = -------- [RS] |
|
On définit un coefficient de fixation NON exclusive, c, comme étant le rapport des constantes de dissociation du substrat de chacun des deux états R et T. En d'autres termes, comme étant le rapport des affinités pour chacun des deux états : |
|||
KR |
[R0]
[S] -------- [RS] c = ---------- => [T0] [S] ------- [TS] |
[R0].[TS] c = ---------- => [T0].[RS] |
1 c = --- . L1 L |
| D'où : L1 = L . c | |||
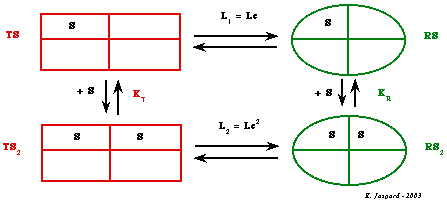
| Fixation
de la 2ème molécule de substrat
|
L1 TS <===> RS |
[TS] L1 = ------ [RS] |
| KT TS + S <===> TS2 |
[TS]
KT = -------- [TS2] |
|
|
L2 TS2 <===> RS2 |
[TS2] L2 = ------- [RS2] |
|
| KR RS + S <===> RS2 |
[RS]
KR = -------- [RS2] |
|
KR |
[RS]
[S] -------- [RS2] c = ---------- => [TS] [S] -------- [TS2] |
[RS].[TS2] c = ----------- => [TS].[RS2] |
1 c = ---- . L2 L1 |
|
D'où : L2 = L . c2 |
|||
|
Fixation de la 3ème et de la 4ème molécule de substrat : |
L3 = L . c3 L4 = L . c4 |
b. Passage aux constantes microscopiques KμT et KμR Considérons la fixation de la 1ère molécule de substrat. D'un point de vue macroscopique, l'équilibre s'écrit : |
|
| KR
R0 + S <===> RS |
[S] [RS] = [R0] . ------ KR |
|
Mais si l'on considère les 4 sites de fixation indépendamment, il existe 4 possibilités de fixer cette 1ère molécule de substrat.
Dès que cette 1ère molécule de substrat est fixée, il n'existe qu'1 possibilité de la dissocier. D'où le facteur statistique permettant de passer des constantes macroscopiques (KT et KR) aux constantes microscopiques (KμT et KμR) : 4 |
|
D'où l'expression de [RS] en fonction de KμR (expression 1) : |
[S] [RS] = [R0] . 4 . ----- KμR |
|
Considérons la fixation de la 2ème molécule de substrat. D'un point de vue macroscopique, l'équilibre s'écrit : |
KR
RS + S <===> RS2 |
[S] [RS2] = [RS] . ----- KR |
|
Si l'on considère les 4 sites de fixation indépendamment : 1 site est dejà occupé donc il existe 3 possibilités de fixer la 2ème molécule de substrat. Dès que cette 2ème molécule de substrat est fixée, il existe 2 possibilités de dissocier une molécule de substrat. |
| D'où : |
3 [S] [RS2] = [RS] . ---- . ----- 2 KμR |
|
En remplaçant [RS] par l'expression 1 : |
[S]
3 [S] [RS2] = [R0] . 4 . ----- . ---- . ----- KμR 2 KμR |
| On obtient (expression 2) : |
[S]2 [RS2] = [R0] . 6 . ------- [KμR]2 |
|
Fixation de la 3ème molécule de substrat : |
KR RS2 + S <===> RS3 |
[S] [RS3] = [RS2] . ------ KR |
|
Fixation de la 4ème molécule de substrat : |
KR RS3 + S <===> RS4 |
[S] [RS4] = [RS3] . ------ KR |
|
Fixation de la 3ème molécule de substrat : 2 sites sont dejà occupés donc il existe 2 possibilités de fixer la 3ème molécule de substrat. Dès que cette 3ème molécule de substrat est fixée, il existe 3 possibilités de dissocier une molécule de substrat. En remplaçant [RS2] par l'expression 2, on obtient (expression 3) : |
[S]3 [RS3] = [R0] . 4 . ------- [KμR]3 |
|
Fixation de la 4ème molécule de substrat : 3 sites sont dejà occupés donc il existe 1 possibilité de fixer la 4ème molécule de substrat. Dès que cette 4ème molécule de substrat est fixée, il existe 4 possibilités de dissocier une molécule de substrat. En remplaçant [RS3] par l'expression 3, on obtient : |
[S]4 [RS4] = [R0] . ------- [KμR]4 |
![]()