| L'acétylation |
| Tweet |
|
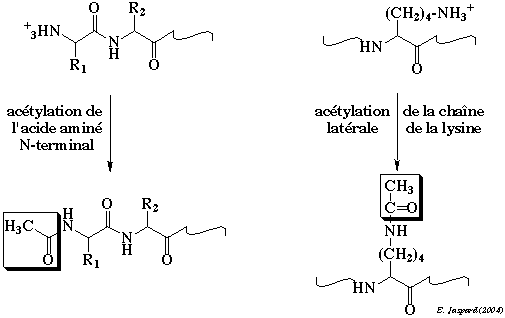
L'acétylation est l'addition d'un groupement CH3-C=O sur les résidus lysine en position N-terminale ou au sein de la chaîne polypeptidique.
La réaction est catalysée par des acétyltransférases. Exemples : la CREB-binding protein (CBP) - [CREB : "cAMP response element binding protein"] et la protéine p300 qui agissent sur les histones. Cette modification post-traductionnelle neutralise la charge positive et modifie la taille de la chaîne latérale des résidus lysine ce qui induit un changement de conformation des protéines modifiées et le mode d'interaction avec leur molécules cibles, en particulier l'ADN qui est chargé négativement. L'acétylation est une modification réversible et le clivage est catalysée par des désacétylases comme les sirtuines. |
|
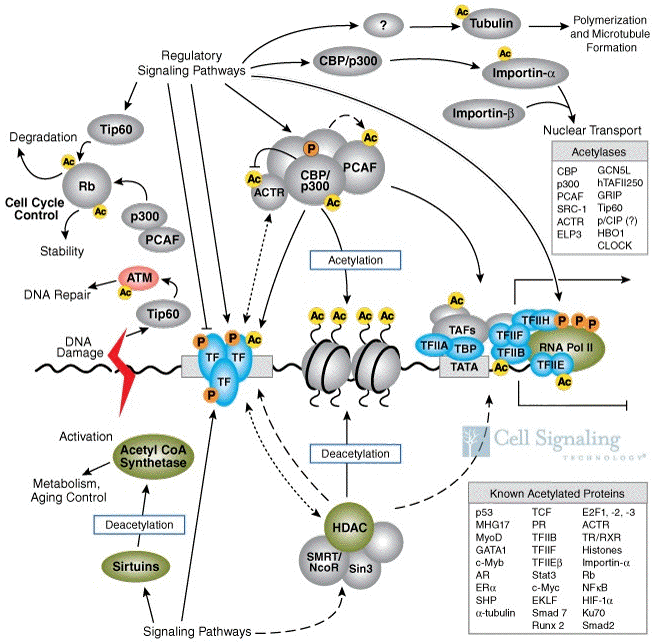
L'acétylation et la désacétylation ont une incidence sur la structure de la chromatine et donc jouent un rôle capital dans la régulation de la transcription des gènes. En effet, des complexes d'acétylation (exemple : [CBP/p300 - PCAF] dans la figure ci-dessous) ou de désacétylation (exemples : Sin3, SMRT/NcoR) sont recrutés par les facteurs de transcription liés à l'ADN ("TF") en réponse à certains signaux ("Regulatory Signaling Pathways").
L'acétylation des histones stimule la transcription :
Source : "Cell signaling" La désacétylation réprime la transcription via des mécanismes inverses. Au niveau de l'organisme, l'acétylation joue un rôle important dans l'immunité, le rhythme circadien et la construction de la mémoire. Sirtuines : famille d'histones désacetylases qui recquièrent le NAD+ comme co-enzyme. |
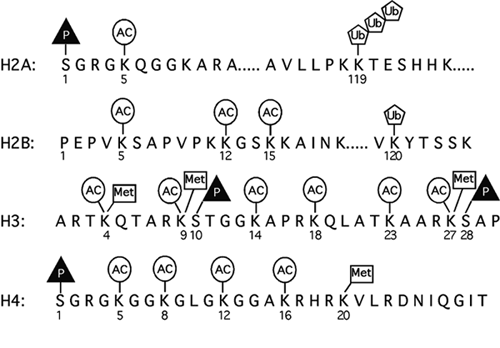
L'acétylation des histones dans le cytoplasme aide à l'assemblage de la chromatine, une fois que les histones ont rejoint le noyau. En effet, l'acétylation des lysines localisées dans la partie N-terminale des histones H3 et H4 empêche l'interaction de cette région avec l'ADN. Celui-ci est donc empaqueté de façon moins serrée par les histones. Les lysines des histones peuvent être modifiées de différentes manières (figure ci-contre).
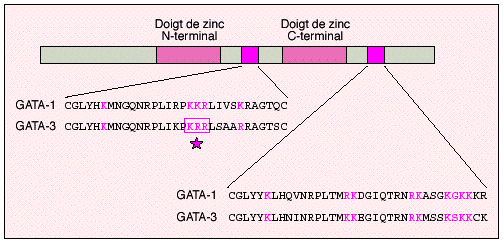
Source : Lee & Young (2000) Ac = acétylation ; Met = méthylation ; Ub = ubiquitination ; P = phosphorylation Deux facteurs de transcription de la famille GATA, les protéines GATA-1 et GATA-3, (impliquées dans la différenciation érythroide et lymphoide T, respectivement) sont aussi acétylés par CBP et p300.
Source : Coulombel (2001)
|
| Références bibliographiques |
|
Dali-Youcef et al. (2007) "Sirtuins: the 'magnificent seven', function, metabolism and longevity" Ann. Med. 39, 335 - 345 Hadnagy et al. (2008) "Histone tail modifications and noncanonical functions of histones: perspectives in cancer epigenetics" Mol. Cancer Ther. 7, 740 - 748 Olsson et al. (2007) "How important are post-translational modifications in p53 for selectivity in target-gene transcription and tumour suppression ?" Cell Death Differ. 14, 1561 - 1575 Tyteca et al. (2006) "To die or not to die: a HAT trick" Mol. Cell 24, 807 - 808 Yang & Seto (2007) "HATs and HDACs: from structure, function and regulation to novel strategies for therapy and prevention" Oncogene 26, 5310 - 5318 Lee & Young (2000) Ann. Rev. Genet. 34, 77 - 137 Coulombel, L. (2001) médecine/sciences 17, 222 - 223 |
----- ----- |
![]()