| Les protéines 14.3.3 |
| Tweet |
|
|
1. Présentation générale 2. Structure des protéines 14.3.3 3. Les protéines cibles des protéines 14.3.3 |
4. Exemples de régulation du métabolisme par les protéines 14.3.3 5. Liens Internet et références bibliographiques |
1. Présentation générale Moore et Perez (1967) ont donné leur nom aux protéines 14.3.3 en fonction :
Les protéines 14.3.3 sont ubiquitaires dans toutes les espèces eucaryotes, à l'exception de la protéine 14.3.3 σ qui n'est trouvée que dans un certain type de cellules. Les protéines 14.3.3 forment un groupe de protéines trés conservées (famille PF00244). Rôles des des protéines 14.3.3 La phosphorylation des protéines est un évènement capital dans la signalisation et donc dans la régulation du métabolisme. Les protéines 14.3.3 fixent plusieurs centaines de protéines qui sont phosphorylées sur des Ser ou des Thr au sein de motifs spécifiques. Ces protéines phosphorylées ("phosphopeptide-binding proteins") ou protéines cibles des protéines 14.3.3 interviennent dans un trés grand nombre de processus :
|
|
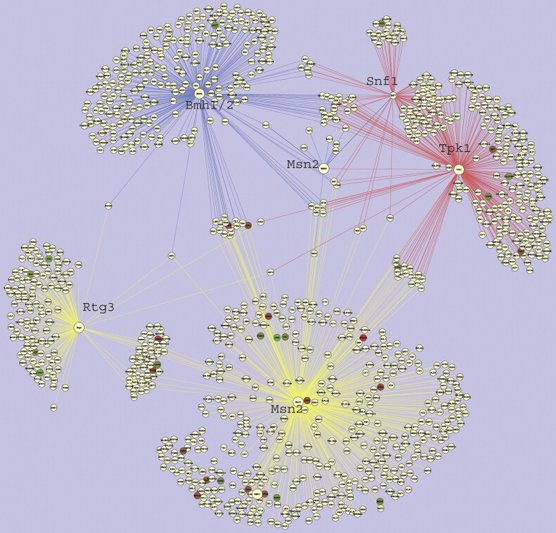
Les protéines 14-3-3 jouent donc un rôle majeur dans les réseaux de régulation cellulaires. Figure ci-dessous, visualisation d'une partie du réseau dans lequel les protéines 14.3.3 de la levure sont impliquées (logiciel "Cytoscape").
Source : van Heusden (2009) Les régions dites nativement désordonnées ou non-structurées ("natively disordered regions") sont des segments de la chaîne polypeptidique d'une protéine qui ne se replient pas dans une structure tridimensionnelle définie. Ces régions sont donc flexibles et jouent des rôles physiologiques clé. Or les phosphoprotéines cibles des protéines 14-3-3 sont riches en acides aminés dits "promoteur de désordre" ("disorder-promoting residues") autour de leurs sites de phosphorylation. Les protéines 14-3-3 interagissent donc avec de multiples cibles qui sont nativement désordonnées ce qui explique leur participation dans de nombreux réseaux de signalisation. |
Les différentes isoformes des protéines 14.3.3 La famille des protéines 14.3.3 est constituée de 7 isoformes (sauf chez les plantes - voir ci-après) : β, γ, ε, σ, ζ, τ et η. Les liens dans le tableau ci-dessous renvoient vers des informations complémentaires pour chaque isoforme.
Exemple de nomenclature : YWHB = "tyrosine 3-monooxygenase / tryptophan 5-monooxygenase activation protein, beta polypeptide" Il existe des protéines 14.3.3 α ("14-3-3 protein beta/alpha") et 14.3.3 δ qui sont les formes phosphorylées des protéines 14.3.3 β et 14.3.3 ζ , respectivement. L'existence d'une trés large famille de gènes codant les protéines 14-3-3 chez les plantes (règne Viridiplantae) traduit leurs rôles essentiels dans la physiologie des plantes. Les protéines 14-3-3 des plantes ont été classées en 4 groupes. On a recensé 13 isoformes de protéines 14-3-3 chez Arabidopsis thaliana. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||
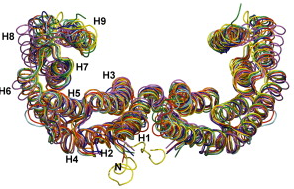
2. Structure des protéines 14.3.3 Les protéines 14.3.3 sont de caractère acide d'une masse molaire d'environ 28 à 33 kDa. Elles s'assemblent en homodimères ou en hétérodimères via des interactions par leurs domaines N-terminaux. Une exception : la protéine 14.3.3 σ ne forme que des homodimères. Il existe un trés haut degré d'identité de séquences en acides aminés (sauf la boucle C-terminale) et donc de conservation de structure entre les 7 isoformes des mammifères comme le montre la superposition tout à fait étonnante des structures des protéines 14.3.3 de l'homme (figure ci-dessous).
Source : Obsil & Obsilova (2011) 14.3.3 β (PDB 2C23) : magenta - 14.3.3 ε (PDB 2BR9) : orange - 14.3.3 γ (PDB 2B05) : rouge - 14.3.3 ζ (PDB 1QJB) vert - 14.3.3 η (PDB 2C74) : cyan - 14.3.3 τ (PDB 2BTP) : jaune - 14.3.3 σ (PDB 1YWT) : bleu |
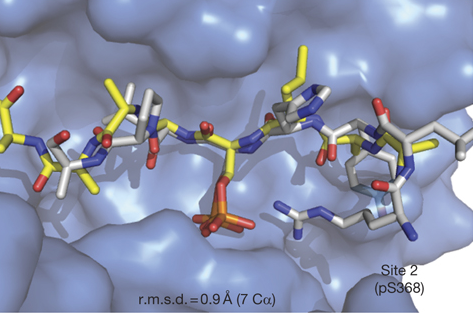
Visualisation de la protéine 14.3.3 ε humaine à une résolution de 1,75 Å Code PDB : 2BR9 Remarque : la protéine 14.3.3 (sous forme de monomère) est complexée à un hexapeptide (en bleu) avec une phosphosérine. On notera le contenu en hélices α antiparallèles.
|
Le motif "Tetratrico Peptide Repeat" - (TPR) est un motif structural dégénéré de 34 acides aminés qui forme 2 hélices α antiparallèles et sert de module pour l'interaction protéine - protéine. Un trés grand nombre de protéines impliquées dans les voies de signalisation via ce genre d'interactions contiennent plusieurs motifs TPR en tandem : cette super-structure confère à ces protéines une forme qui permet "d'enrober" les protéines cibles avec lesquelles elles interagissent. Chaque momomère des protéines 14-3-3 contient un ensemble de 9 hélices alpha antiparallèles (H1 à H9 - figure ci-dessus) qui forment une surface concave en forme de "L. Au sein de cette surface est dessiné une sorte de "sillon" de caractère amphiphile puisqu'il est constitué :
Ce "sillon" amphiphile (figure ci-dessous) d'environ 35 Å de largeur et 20 Å de profondeur, constitue le site de fixation des protéines cibles.
Source : Kostelecky et al. (2009) Les acides aminés qui forment les surfaces d'interactions entre monomères au sein des dimères sont trés conservés entre les différentes isoformes et dans tous les règnes (animaux, plantes, levure). En revanche, les séquences en acides aminés de la boucle C-terminale sont trés variables et cette partie des protéines 14-3-3 est structuralement désordonnée. Les protéines 14.3.3 ζ sont des isoformes tronquées dans cette région et elles fixent mieux certaines protéines cibles : bien que l'on en connaisse pas la structure, il a été suggéré que cette partie C-terminale se comporterait comme un "suppresseur" de liaisons / fixation non spécifiques avec des ligands inappropriés. L'isoforme ζ présente une autre particularité : la surface d'interaction entre les monomères des protéines 14.3.3 ζ contient 3 ponts salins dont un seul est trouvé dans toutes les autres isoformes de protéines 14.3.3. Les protéines 14.3.3 sont elles même phosphorylées sur de nombreux résidus. |
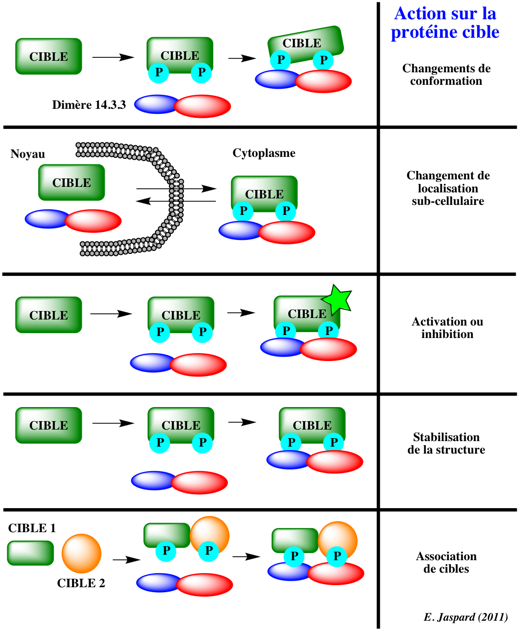
3. Les protéines cibles des protéines 14.3.3 Chaque monomère contient un site de fixation du ligand indépendant : le dimère peut donc interagir avec deux protéines cibles simultanément. Des centaines de protéines signal (protéines cibles) ont été recensées comme ligands des protéines 14-3-3. Exemples de protéines (liste non exhaustive) avec lesquelles la protéine 14.3.3 tau (τ) interagit : CDK16, SSH1, CDKN1B, GAB2, MEF2D, "Bcl-2-associated X protein", "Bcl-2-associated death promoter", CRTC2, C-Raf, histone déacétylase 5, NRIP1, protéine kinase D1, protéine kinase Mζ, UCP3, PFKFB2, télomerase reverse transcriptase, ... Cette aptitude des protéines 14.3.3 à se fixer à autant de protéines cibles rappelle celle de la calmoduline. La fixation protéines 14-3-3 à leurs protéines cibles induit :
Adaptée de Gökirmak et al. (2010) Parmi les changements de localisation sub-cellulaire des protéines cibles, on peut mentionner :
|
Motifs phosphorylés des protéines cibles reconnus par les protéines 14.3.3 Les protéines 14-3-3 reconnaissent des séquences consensus spécifiques contenant la Ser ou la Thr phosphorylée. Elles reconnaissent 2 types de séquences consensus et dans la littérature on trouve les expressions "mode I" et "mode II". |
| mode I | mode II | mode III |
| R(S/Φ)X(pS/pT)XP | RX(Φ/S)X(pS/pT)XP | L'acide aminé phosphorylé est l'avant dernier de la chaine polypeptidique : (pS/pT)X-COOH |
| R(S/Φ)[basique]pSXP | RX[Φ/S][basique]pSXP | |
| RXX(pS/pT)XP chez les plantes : LX(R/K)SX(pS/pT)XP |
--------- | |
| R[SFYW]XpSXP | RX[SYFWTQAD]Xp(S/T)X[PLM] | |
| phosphopeptides synthétiques : RSX(pS)XP | phosphopeptides synthétiques : RX(F/Y)X(pS)XP | |
Nomenclature : "/" = "ou"; "Φ" = aromatique; "pS/pT" = Ser/Thr phosphorylée; "X" = n'importe quel acide aminé. |
||
Les séquences consensus décrites dans le tableau ci-dessus dépendent du type d'étude bioinformatique (recherche de motifs par alignement de séquences et/ou comparaison de structures tri-dimensionnelles). Celà dépend donc aussi du nombre de séquences et de structures disponibles de protéines 14.3.3 (nombre qui augmente avec le temps). Les premières structures cristallines de complexes [protéines 14-3-3 / phosphopeptides synthétiques] ont montré que :
Il existe aussi un motif de reconnaisance mode III où l'acide aminé phosphorylé est l'avant dernier (C-term -1). L'exemple le mieux caractérisé d'un tel motif est la pompe à protons de la membrane plasmique des plantes : un dimère de protéine 14-3-3 se fixe à l'extrémité phosphorylée du côté cytoplasmique de 2 sous-unités adjacentes de la pompe, de sorte que 3 dimères de protéines 14-3-3 stabilisent la forme héxamèrique active de cette pompe. |
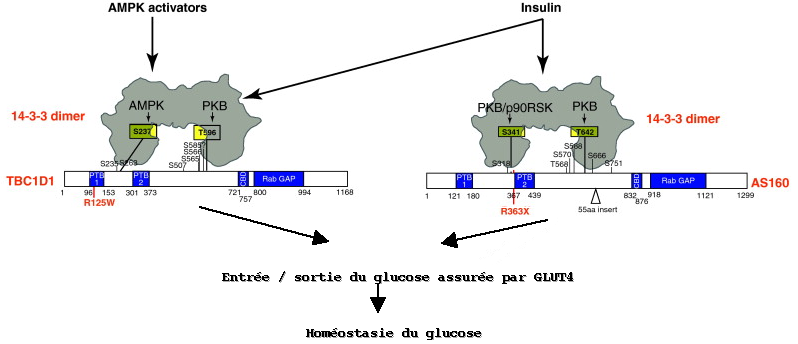
4. Exemples de régulation du métabolisme par les protéines 14.3.3 a. Médiation de l'action de l'insuline par les protéines 14.3.3 L'étude du phosphoprotéome (ensemble des protéines phosphorylées) a montré qu'il est probable que les protéines 14-3-3 interagissent avec les protéines [GTPase - activatrices de Rab] AS160 et TBC1D1 pour réguler les besoins en glucose dans certains tissus en réponse à l'insuline et à un stress énergétique. Dans une cellule non stimulée, GLUT4 (transporteur du glucose) est localisé dans des vésicules de stockage qui doivent fusionner avec la membrane plasmique avant que le glucose ne pénètre dans la cellule. Dans les muscles squelettiques et cardiaque, l'entrée/sortie du glucose assurée par GLUT4 est activée par la contraction, qui à son tour active des protéines kinases, incluant la protéine kinase activée par l'AMP (AMPK) qui est activée lors d'un abaissement de la concentration en ATP. L'entrée/sortie du glucose assurée par GLUT4 recquiert que les protéines [Rab liées au GTP] soient sous forme active. Or les protéines TBC1D1 et AS160 induisent l'hydrolyse du GTP par Rab. Il semble que ces complexes [Rab/TBC1D1] et [Rab/AS160] soient inactivés par phosphorylation via l'insuline et l'AMPK, ce qui maintient le GTP fixé sur Rab et facilite l'entrée/sortie du glucose assurée par GLUT4. Les protéines TBC1D1 et AS160 ont des structures proches et possèdent deux sites de fixation aux protéines 14-3-3 et un autre domaine d'interactions PTB1 et PTB2 (qui contient des phosphotyrosines). Figure ci-dessous, schéma hypothètique des mécanismes de régulation complémentaires de l'homéostasie du glucose par les protéines TBC1D1 et AS160 dans les muscles squelettiques :
Source : Chen et al. (2011) Figure ci-dessus :
Les deux hypothèses faites dans ce modèle sont :
Cette hypothèse ne prend pas en compte les autres signaux perçus par TBC1D1 et AS160 via des sites de phosphorylation trés proches des sites de fixation des protéines 14-3-3. |
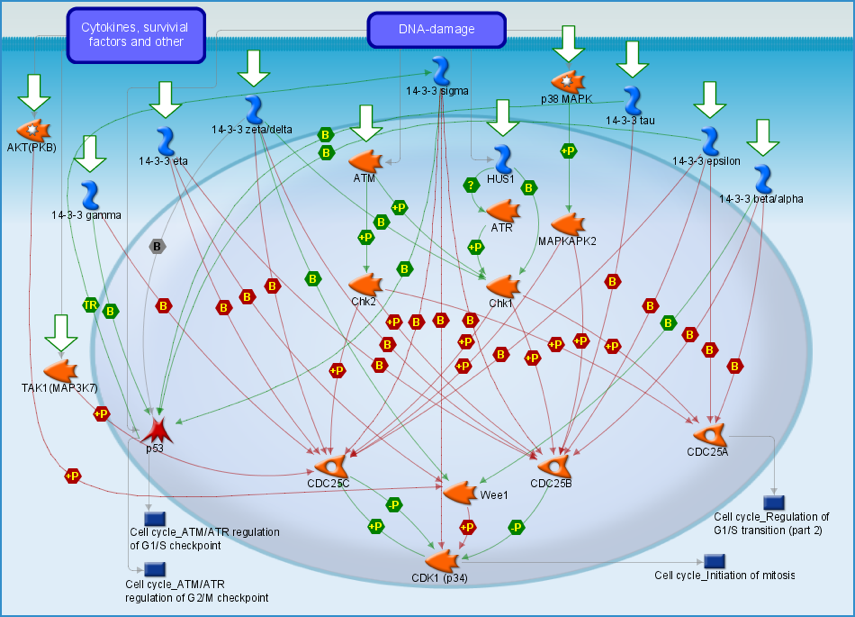
b. Régulation du cycle cellulaire par les protéines 14.3.3 Les protéines 14.3.3 régulent le cycle cellullaire et préviennent l'apoptose en contrôlant la distribution nucléaire et cytoplasmique des molécules signal avec lesquelles elles interagissent.
Source : Life Science Research Les protéines 14.3.3 assurent des fonctions cruciales pendant la division cellulaire. En effet, divers mécanismes impliquant des complexes [protéines 14-3-3 / protéines cibles] permettent que la mitose ne soit pas prématurément activée avant que ne soit finie la réplication de l'ADN au cours de l'interphase. Les protéines 14.3.3 régulent le cycle cellulaire via la protéine kinase "checkpoint kinase 1" (Chk1) et en phosphorylant les phosphatases de la division cellulaire CDC25 ("cell division cycle 25"), la tyrosine kinase Wee1, le suppresseur de tumeur p53 et la "cyclin-dependent kinase 1"(CDK1). Les CDC25 sont la cible principale de la régulation par les protéines 14.3.3 : les CDC25 activent les CDK par déphosphorylation, stimulant ainsi la progression du cycle cellulaire. Différentes CDC25 participent aux différentes phases du cycle cellulaire :
L'absence de formes actives de CDC25 résulte en une accumulation de la forme phosphorylée de CDK (forme inactive), incapables de participer à l'initiation de la réplication. De plus, la voie [MAP kinase p38 / MAPKAPK2] ("mitogen-activated protein kinase-activated protein kinase 2") et TAK1 participent à la phosphorylation de la CDC25B et de la CDC25C. De nombreuses isoformes de protéines 14.3.3 inhibent les CDC25 phosphorylées, et il en résulte probabement une rétention des CDC25 dans le cytoplasme et/ou un accès bloqué des CDK au site catalytique des CDC25. Quand la mitose est activée, la séquestration de la CDC25C dans le cytoplasme par les protéines 14.3.3 est inhibée par une phosphorylation mediée par la CDK1. CDK1 est régulée directement par les isoformes 14-3-3 σ ou via Wee1. Wee1 inhibe la Cdk1 par phosphorylation de la Tyr 15. Wee1 peut être activée par les isoformes 14-3-3 [α / β] et 14-3-3 [δ / ζ ]. Wee1 phosphorylée par la protéine kinase B (AKT) sur la Ser 642 peut-être retenue dans le cytoplasme par les isoformes 14-3-3 η. Les isoformes 14.3.3 γ, τ, ε et γ peuvent activer p53. Mais une déphosphorylation de la Ser 376 de p53, induite par des radiations ionisantes, est nécessaire pour que ce processus ait lieu. |
| 5. Liens Internet et références bibliographiques |
| Moore & Perez (1967) "Specific acidic proteins of the nervous system", FD Carlson. ed. "Physiological and biochemical aspects of nervous integration", Prentice-Hall, Inc, The Marine Biological Laboratory, Woods Hole, MA. pp. 343 - 359 | |
|
Kostelecky et al. (2009) "Recognition of an intra-chain tandem 14-3-3 binding site within PKCε" EMBO reports 10, 983 - 989 van Heusden (2009) "14-3-3 proteins: Insights from genome-wide studies in yeast" Genomics 94, 287 - 293 Gökirmak et al. (2010) "Plant phosphopeptide-binding proteins as signaling mediators" Curr. Opin. Plant Biol. 13, 527 - 532 Chen et al. (2011) "The capture of phosphoproteins by 14-3-3 proteins mediates actions of insulin" Trends Endocrinol. & Metab.22, 429 - 436 |
|
|
Johnson et al. (2010) "Bioinformatic and experimental survey of 14-3-3-binding sites" Biochem J. 427, 69 - 78 Obsil & Obsilova (2011) "Structural basis of 14-3-3 protein functions" Semin. Cell. Dev. Biol. (sous-presse) Denison et al. (2011) "14-3-3 proteins in plant physiology" Semin. Cell. Dev. Biol. (sous-presse) |
|
|
Life Science Research : plusieurs centaines de voies de signalisation et autres outils. PINA : "Protein Interaction Network Analysis platform" |
|
![]()