{kind=link}
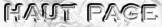
Optimisation des conditions d'expression et purification partielle de la forme mature de la protéine de choc thermique HSP22 recombinante de pois (Pisum sativum) Résultats |
|
Voir un cours sur les protéines de stress LEA et HSP. |
|
1. Courbes de croissance des souches d'Escherichia coli
2. Optimisation des conditions d'expression : température et concentration en IPTG
3. Optimisation des conditions d'expression : effet d'additif(s) dans le milieu de culture |
b. Souche B834(DE3)pLysS
4. Optimisation des conditions d'expression : cinétique de l'effet du PEG ou/et du sorbitol 5. Purification partielle de l'HSP22 recombinante |
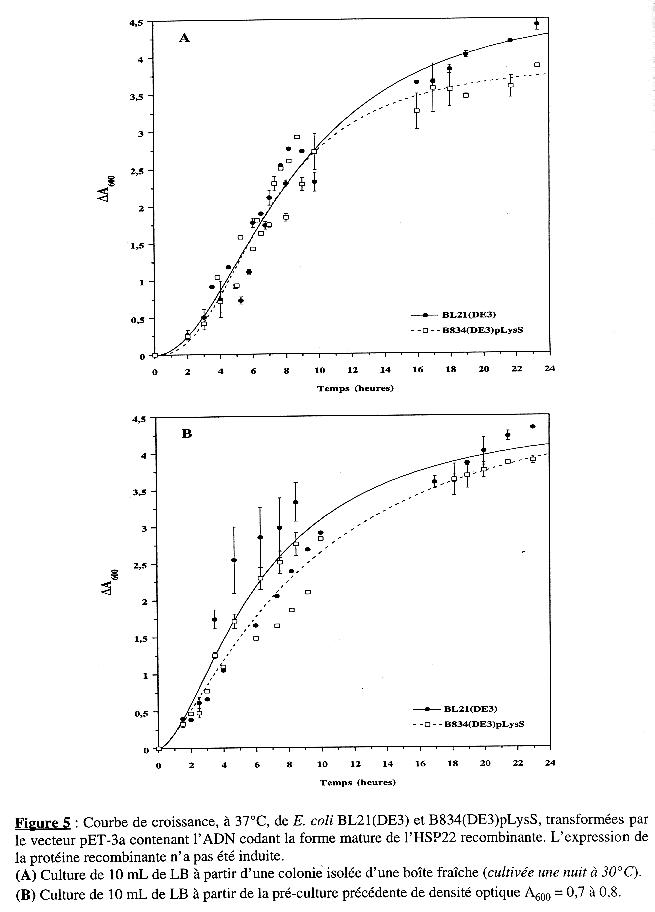
1. Courbes de croissance des souches d'Escherichia coli Les deux souches BL21(DE3) et B834(DE3)pLysS ont été cultivées, sans induction de l'expression, à partir d'une colonie isolée d'une boîte fraîche (cultivée à 30°C) et à partir de cette pré-culture d'absorbance A600 = 0,7 à 0,8. Les courbes de croissance sont présentées dans la Figure 5. Elles ont été analysées avec l'équation suivante : tn Le tableau suivant résume les valeurs des paramètres des courbes de croissance :
Ces résultats indiquent que les deux souches ne se distinguent pas par leur croissance. (Le paramètre n n'a pas de signification biologique, mais il est nécessaire pour lisser les courbes qui sont d'allure sigmoïdale). |
||||||||||||||||||||||||
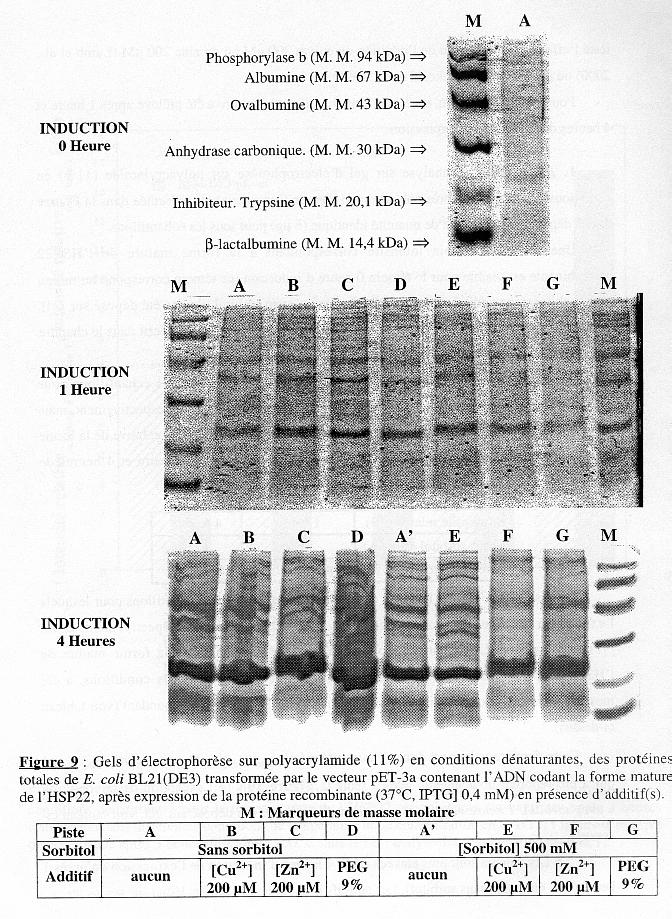
2. Optimisation des conditions d'expression : température et concentration en IPTG L'expression de la forme mature de l'HSP22 recombinante a été induite à 2 températures (37°C et 42°C) et avec 3 concentrations d'IPTG (0,1 mM, 0,4 mM et 1 mM). Pour chaque condition, un échantillon du milieu de culture a été prélevé après 1 heure et 3 heures d'induction de l'expression. Le témoin d'induction 0 heure est un échantillon prélevé juste avant d'induire l'expression. L'analyse sur gel d'électrophorèse en conditions dénaturantes (après coloration au bleu de Coomassie) est présentée dans les Figures 6 & 7 pour BL21(DE3) et pour B834(DE3)pLysS, respectivement. Les dépôts sur gel ont été de volume identique (15 µL) pour tous les échantillons. Pour toutes les expériences, nous avons considéré que la bande de masse molaire (M. M.) apparente = 21,5 kDa correspond à la forme mature de l'HSP22 recombinante parce que :
Pour chaque piste des gels présentés dans les Figures 6 & 7, l'intensité de toutes les bandes et celle de la bande correspondant à la forme mature de l'HSP22 recombinante ont été déterminées avec le logiciel de densitomètrie NIH Image 1.59 (voir l'exemple en "Annexe"). |
|
En ce qui concerne le témoin 0 heure d'induction à 37°C (température contrôle), une bande pour la forme mature de l'HSP22 recombinante (d'intensité relative 8,0 %) est visible dans le cas de BL21(DE3) (piste A du 1er gel, Figure 6), mais pas dans le cas de B834(DE3)pLysS (piste A du 1er gel, Figure 7). Ce résultat est dû à la "fuite" d'expression dans le cas de la souche non co-transformée par le plasmide pLysS (voir le chapitre "Matériels et Méthodes"). L'intensité relative dans les mêmes conditions (37°C, sans IPTG) après 1 heure et 3 heures de culture (piste A du 2 ème et du 3ème gel des Figures 6 et 7) sont respectivement :
La valeur obtenue après 1 heure de culture pour BL21(DE3) n'est pas interprétable. Les valeurs obtenues pour B834(DE3)pLysS indiquent que la présence du plasmide pLysS n'empêche pas complètement cette "fuite", mais que cette dernière est nettement moindre en sa présence et qu'elle ne devient significative que plus tardivement. |
b. Différentes conditions d'induction Du fait de la valeur ambigue obtenue après 1 heure de culture en absence d'induction pour BL21(DE3), nous n'avons reporté les résultats obtenus dans différentes conditions d'induction que pour 3 heures. La Figure 8 représente l'intensité relative de la bande pour la forme mature de l'HSP22 recombinante sur-exprimée dans une condition d'induction i moins l'intensité relative de cette même bande sur-exprimée à 37°C sans IPTG (c'est-à-dire 8,4 et 1,3, respectivement). La température La température de 42°C a été testée afin d'induire un stress thermique des souches d'E. coli étudiées et, le cas échéant, la sur-expression d'une HSP endogène. On remarque qu'en absence d'IPTG pour les deux souches (piste C, Figures 6 & 7) et en présence d'IPTG 0,4 mM pour B834(DE3)pLysS (piste D, Figure 7) il y a apparition d'une bande de M. M. apparente équivalente à celle de la forme mature de l'HSP22 recombinante (d'intensité relative 5,7, 3,9 et 7,7, respectivement, Figure 8). La littérature (voir le chapitre "Introduction") et, depuis peu, le séquençage complet du génome de E. coli K-12 (Blattner et al., 1997) indiquent que plusieurs dizaines de protéines de choc thermiques, de tailles très variable, sont synthétisées par cette bactérie. Parmi ces protéines, il en existe au moins 1 d'une taille comparable à la forme mature de l'HSP22 de pois : l'HSP htrC qui contient 179 acides aminés [E. coli K-12, souche MG1655; accession AAC43087] (Blattner et al., 1993). En conséquence, en l'absence d'un crible tel que la révélation de l'HSP22 de pois recombinante avec un anticorps spécifique, on ne peut exclure la possibilité de l'induction par la température d'une HSP endogène de E. coli de même taille que la protéine étudiée. La concentration en IPTG La Figure 8 indique les valeurs suivantes d'intensité relative :
Par la suite, nous avons opté pour les conditions d'induction suivantes : 37°C et [IPTG] 0,4 mM. |
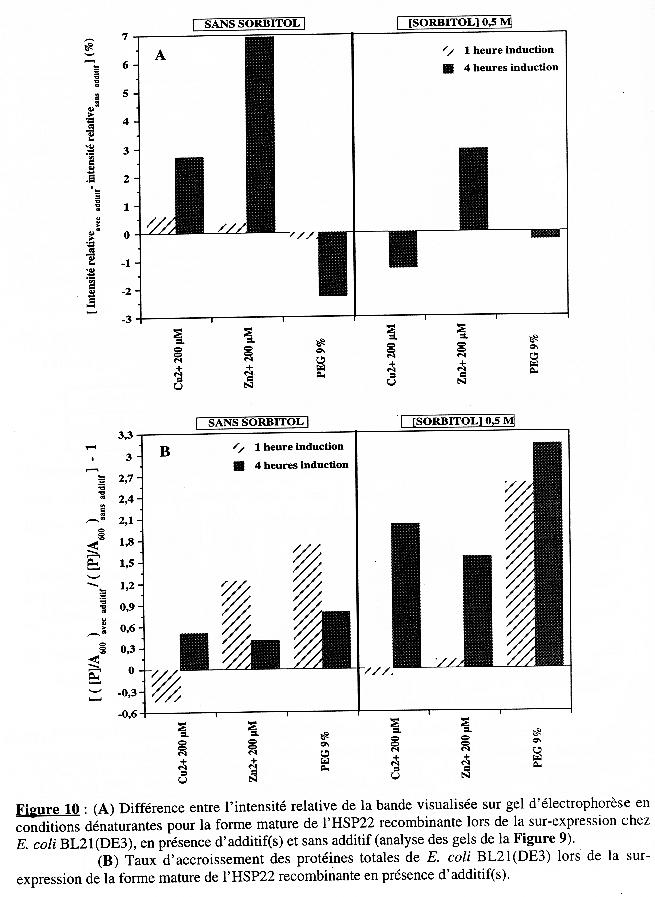
3. Optimisation des conditions d'expression : effet d'additif(s) dans le milieu de culture Les deux souches ont été cultivées dans le milieu de culture contenant ou non du sorbitol 500 mM (Gunasekera & Kemp, 1999). Puis au moment de l'induction, nous avons testé l'effet sur l'expression de l'addition du cuivre 200 µM ou du zinc 200 µM (Lamb et al., 2000) ou du PEG 9% (Umakoshi et al., 1996). Pour chaque condition, un échantillon du milieu de culture a été prélevé après 1 heure et 4 heures d'induction de l'expression. |
|
L'analyse sur gel d'électrophorèse sur polyacrylamide (11%) en conditions dénaturantes (après coloration au bleu de Coomassie) est présentée dans la Figure 9. Les dépôts sur gel ont été de quantité identique (6 µg) pour tous les échantillons. Une bande de faible intensité correspondant à la forme mature de l'HSP22 recombinante est visible pour le témoin 0 heure d'induction (ce témoin correspond au milieu sans sorbitol, sans additif et sans IPTG ; le témoin avec sorbitol n'a pas été déposé sur gel). L'apparition de cette bande en absence d'induction confirme le résultat décrit dans le chapitre précédent. Les pistes A et A' (2ème et 3ème gel) correspondent au dépôt des échantillons pour lesquels l'expression a été induite en absence ou en présence de sorbitol, respectivement, mais sans additif. Ils sont donc les témoins de l'effet de ces additifs. L'intensité relative de la bande correspondant à la forme mature de l'HSP22 recombinante, après 1 heure et 4 heures de culture dans ces conditions, sont respectivement :
Les pistes (B, C, D) et (E, F, G) ) correspondent au dépôt des échantillons pour lesquels l'expression a été induite en absence ou en présence de sorbitol, respectivement, et en présence d'additif.
Elle a été reportée pour chaque additif dans la Figure 10A, à l'exception de la condition "avec sorbitol, 1 heure" puisque cet échantillon n'a pas été déposé sur gel, (tableau ci-dessus).
|
La concentration en protéines totales a été mesurée pour chaque échantillon. Cependant, la présence d'additif et/ou de sorbitol peut modifier la cinétique de croissance des bactéries. Si tel est le cas, les prélèvements étant effectués aux mêmes temps (1 heure et 4 heures), la quantité de bactérie diffère d'un échantillon à l'autre. Pour tenir compte de cette possible différence, nous avons normalisé la concentration en protéines en la divisant par la valeur d'absorbance à 600 nm mesurée pour chaque échantillon au moment du prélèvement, soit ([P] / A600 avec additif). Puis ce rapport a été divisé par celui du témoin correspondant dont les valeurs sont présentées dans le tableau suivant :
L'expression globale est donc [([P] / A600 avec additif) / ([P] / A600 sans additif)] et elle vaut 1 s'il n'y a aucun effet. Afin d'exprimer le taux d'accroissement en protéines totales, nous avons soustrait 1 à l'expression précédente et reporté les valeurs obtenues pour chaque additif (Figure 10B).
Conclusion Pour une condition donnée, une valeur forte observée dans la Figure 10A et une valeur faible observée dans la Figure 10B indiquent que l'augmentation de la quantité de protéines lors de l'expression est due essentiellement à la synthèse de la forme mature de l'HSP22. Cela semble le cas pour :
|
|
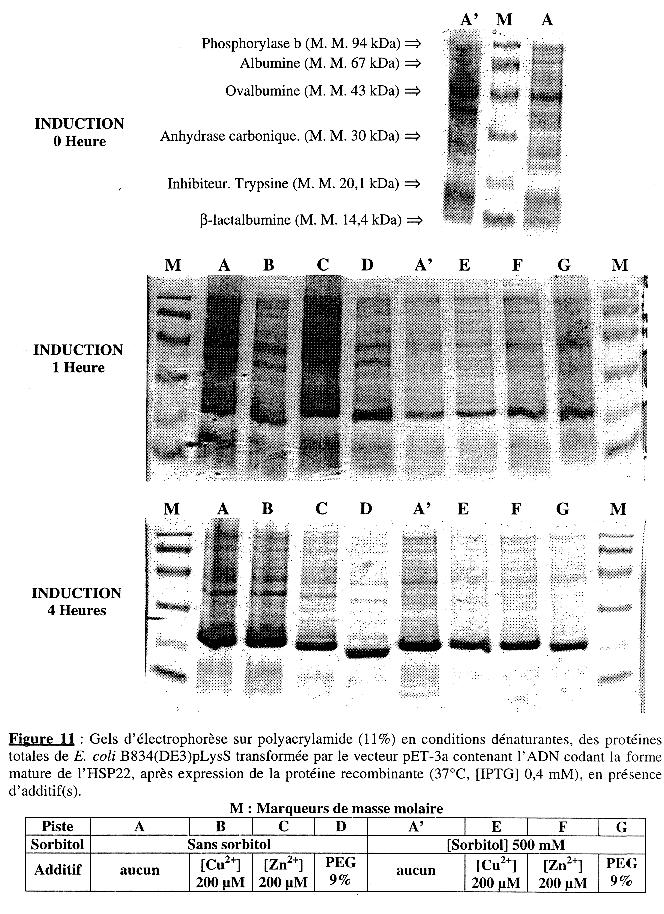
L'analyse sur gel d'électrophorèse en conditions dénaturantes est présentée dans la Figure 11. Les dépôts sur gel ont été de quantité identique (30 µg) pour tous les échantillons. Les pistes A et A' (2ème et 3ème gel) correspondent au dépôt des échantillons pour lesquels l'expression a été induite en absence ou en présence de sorbitol, respectivement, mais sans additif. Ils sont donc les témoins de l'effet de ces additifs. L'intensité relative de la bande correspondant à la forme mature de l'HSP22 recombinante, après 1 heure et 4 heures de culture dans ces conditions, sont respectivement :
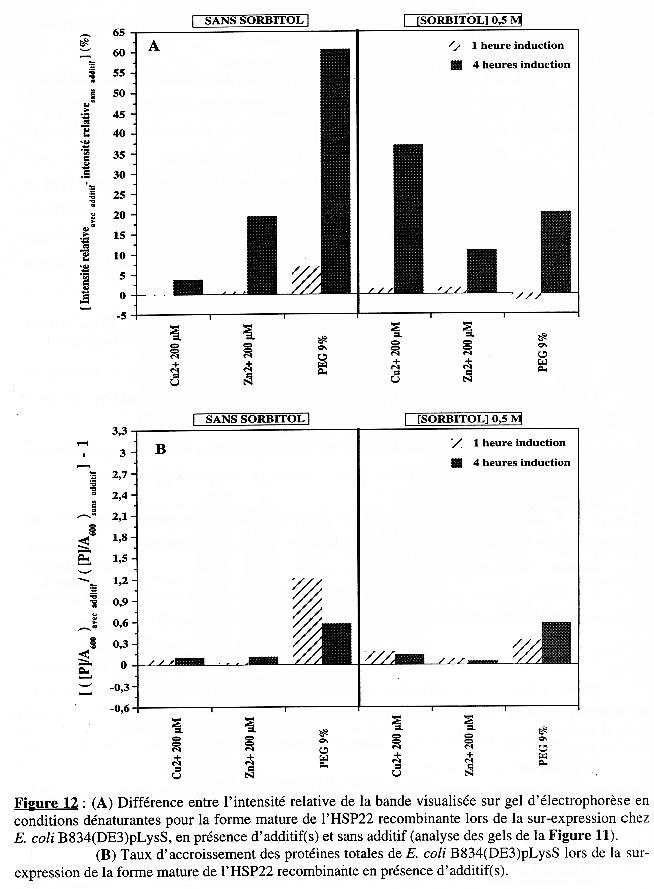
Puis la valeur obtenue pour : (intensité relativeavec additif - intensité relativesans additif) a été reportée pour chaque additif dans la Figure 12A.
|
En ce qui concerne les protéines totales, les valeurs obtenues pour les échantillons sans additif sont respectivement :
La valeur obtenue pour [([P] / A600 avec additif) / ([P] / A600 sans additif)] - 1 a été reportée pour chaque additif dans la Figure 12B. Le taux d'accroissement est moindre pour B834(DE3)pLysS que pour BL21(DE3) (voir Figure 10B). Hormis en présence de PEG, il semble même peu significatif, que ce soit après 1 heure ou 4 heures d'induction de l'expression et ce, en absence ou en présence de sorbitol. Conclusion Si l'on tient compte d'une valeur forte dans la Figure 12A et conjointement d'une valeur faible dans la Figure 12B, on s'aperçoit que :
sont des conditions qui induisent une augmentation de la synthèse spécifique de la forme mature de l'HSP22 recombinante chez B834(DE3)pLysS. |
|
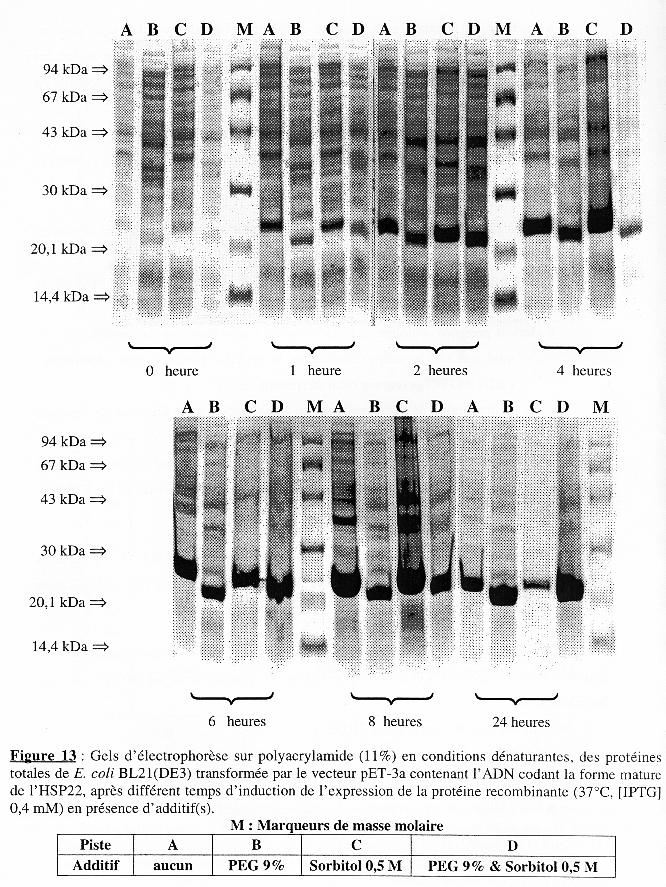
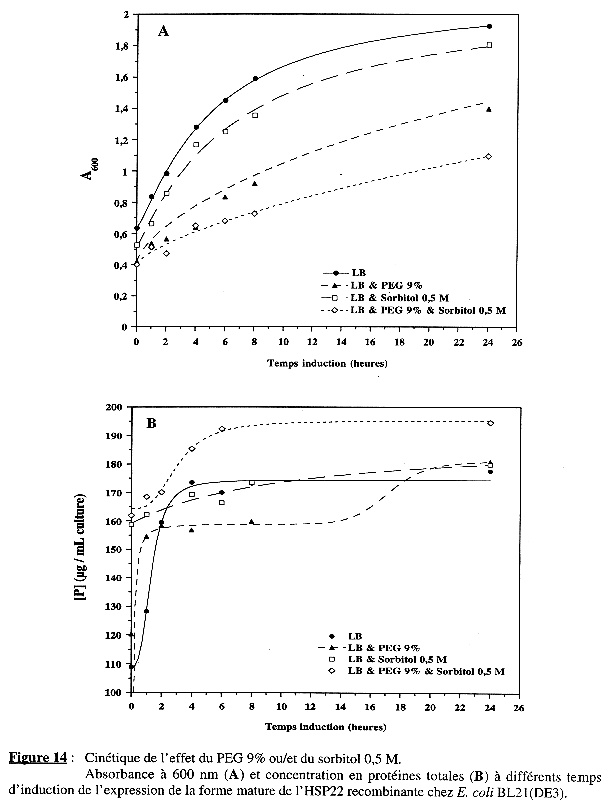
L'analyse sur gel d'électrophorèse sur polyacrylamide (11%) en conditions dénaturantes est présentée dans la Figure 13. Les dépôts sur gel ont été de quantité identique (50 µg) pour tous les échantillons. Quelle que soit la condition, l'intensité apparente de la bande correspondant à la forme mature de l'HSP22 recombinante augmente avec le temps d'induction de l'expression. La Figure 14A montre que la présence d'additif(s) ralentit la croissance des bactéries pendant l'induction de l'expression. Le ralentissement est plus accentué en présence de PEG 9% qu'en présence de sorbitol 0,5 M et l'effet des deux additifs est cumulatif. La Figure 14B montre la variation des protéines totales et le tableau suivant en résume les principales caractéristiques :
|
|
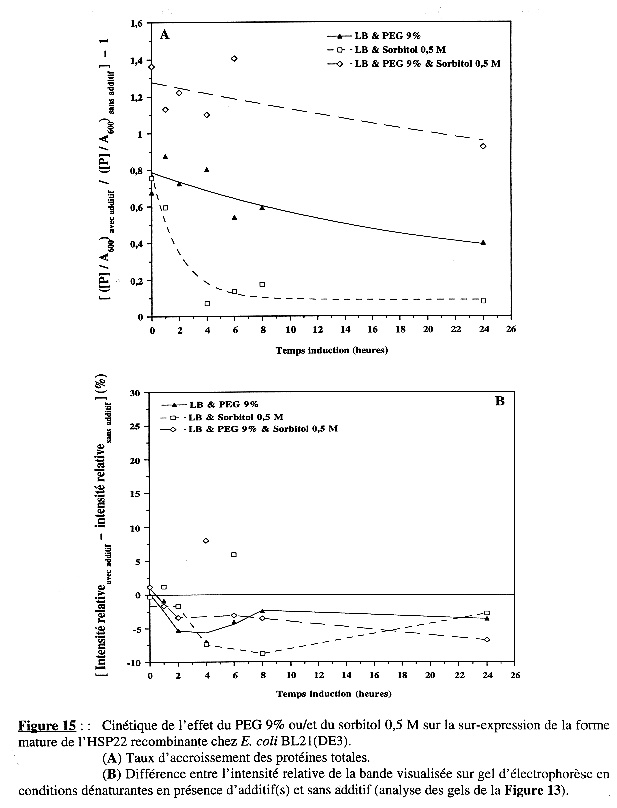
La Figure 15A représente la variation du taux d'accroissement des protéines totales en fonction du temps d'induction de l'expression :
La Figure 15B indique, globalement, que le PEG 9% ou le sorbitol 0,5 M ou les deux ensemble n'ont pas d'effet sur la synthèse spécifique de la forme mature de l'HSP22 recombinante sur-exprimée chez BL21(DE3). Conclusion L'addition du PEG 9% seul ou avec le sorbitol 0,5 M augmente la synthèse nette des protéines dans les premières heures d'induction chez BL21(DE3), mais n'augmente pas celle de la forme mature de l'HSP22 recombinante. |
|
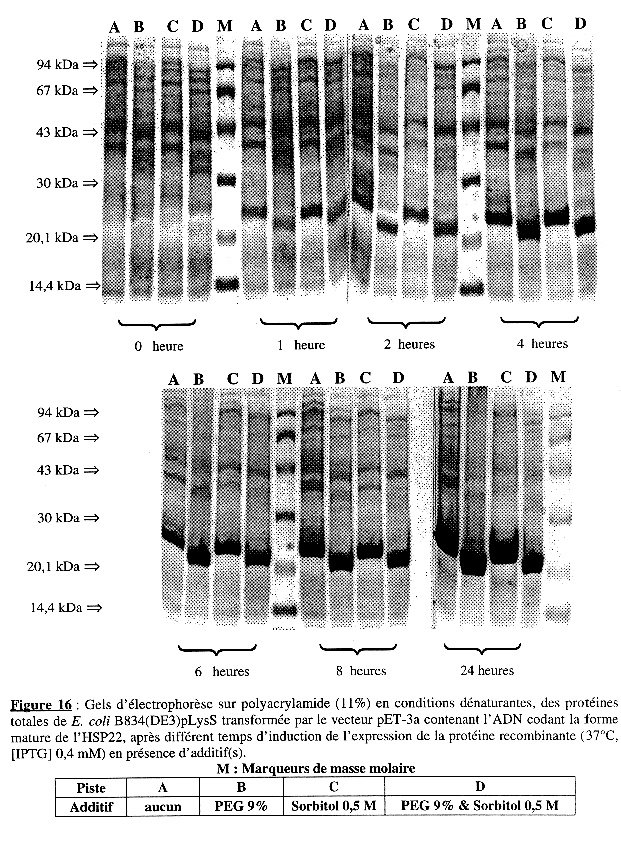
L'analyse sur gel d'électrophorèse en conditions dénaturantes est présentée dans la Figure 16. Les dépôts sur gel ont été de quantité identique (50 µg) pour tous les échantillons. De la même manière que pour BL21(DE3) :
|
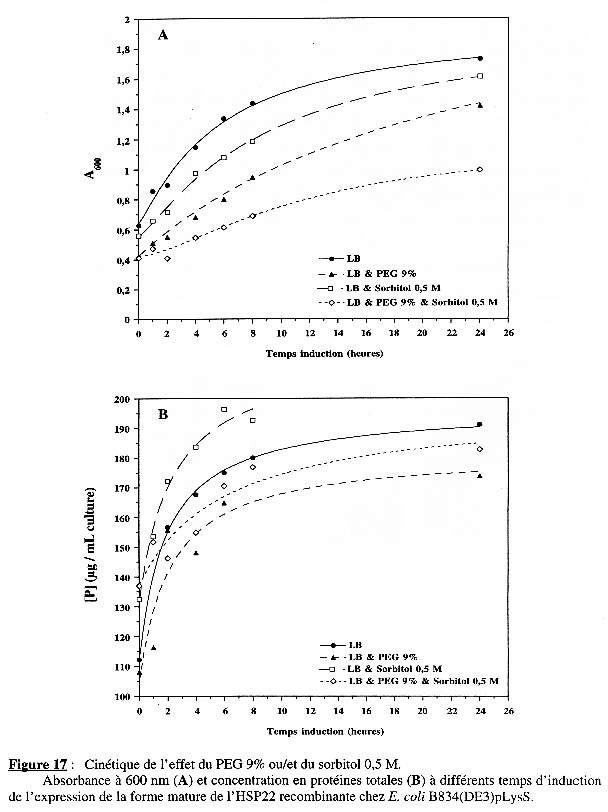
La Figure 17B et le tableau suivant décrivent l'effet des additifs sur la variation des protéines totales :
A la différence de BL21(DE3) :
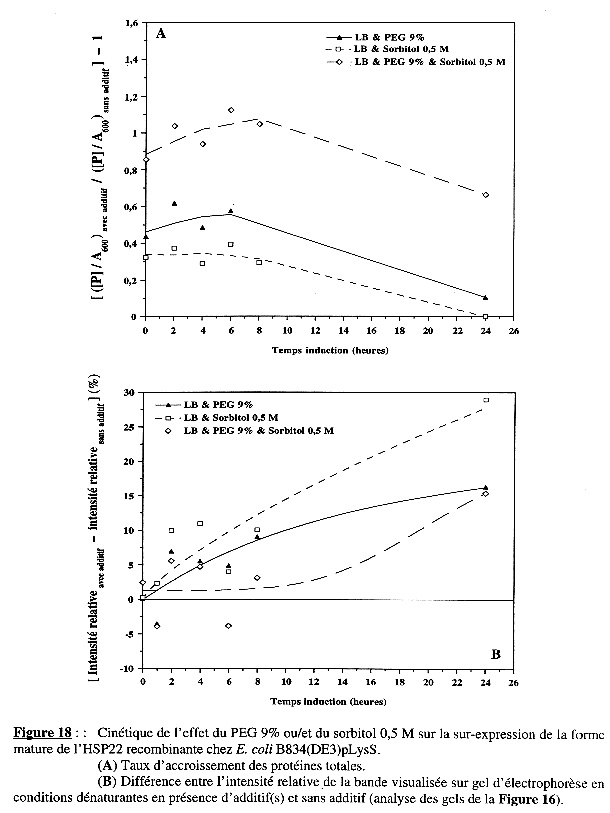
La Figure 18A représente la variation du taux d'accroissement des protéines totales en fonction du temps d'induction de l'expression : hormis la cinétique du phénomène observé (une augmentation durant les 6 premières heures d'induction puis une diminution de la synthèse nette des protéines), les effets des additifs seuls et ensemble sont les mêmes que pour BL21(DE3). |
La Figure 18B montre les effets des additifs sur la synthèse spécifique de l'HSP22 recombinante.
Si l'on fait le parallèle entre les résultats présentés dans les Figures 18A & 18B pour le PEG 9% seul et le sorbitol 0,5 M seul, il est tentant de conclure que, pendant les 6 premières heures d'induction, l'accroissement des protéines totales résulte essentiellement de la synthèse de la forme mature de l'HSP22 recombinante. |
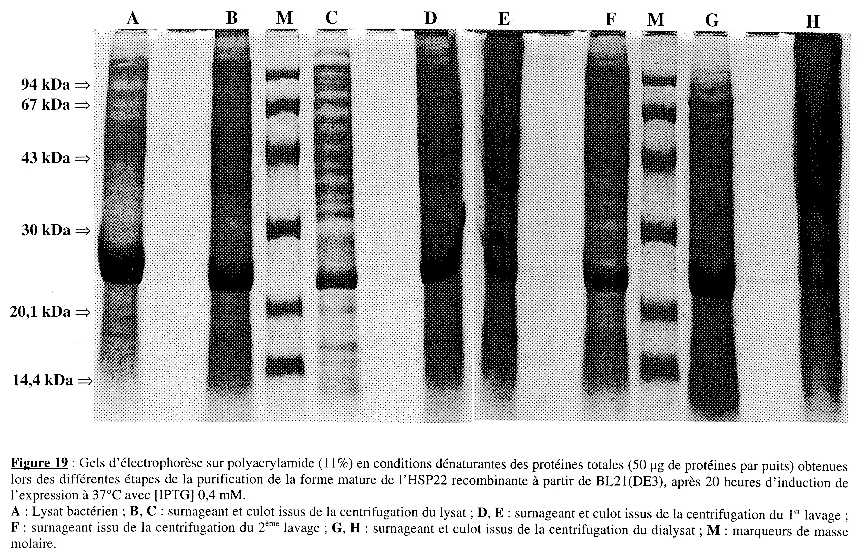
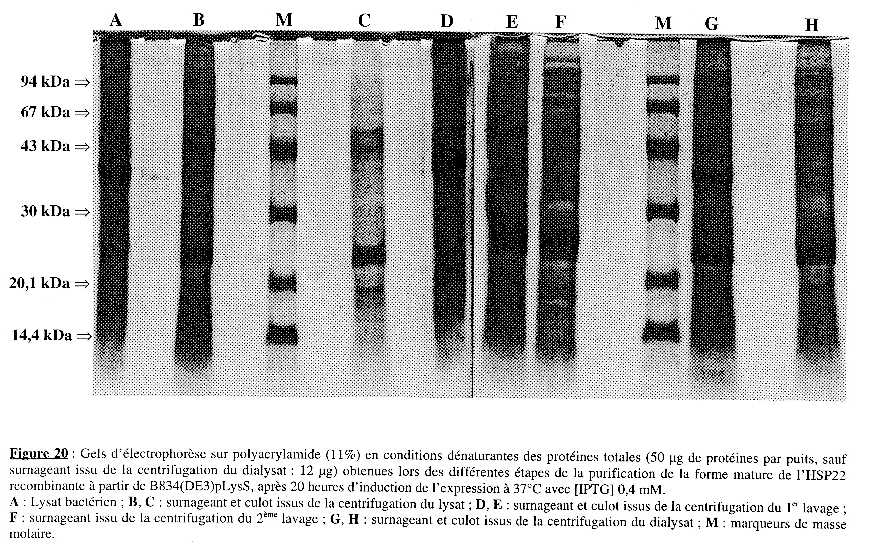
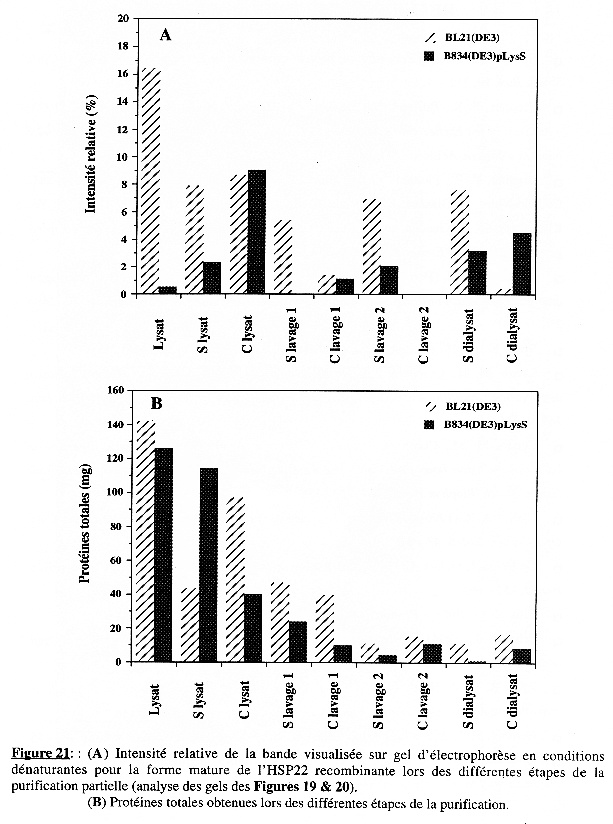
5. Purification partielle de l'HSP22 recombinante Une première purification partielle (résultats non présentés) à partir du culot de bactéries de 100 mL de culture (pour chaque souche), après 20 heures d'induction de l'expression, a permis de " cerner " les conditions requises pour chaque étape (volume pour reprendre les culots, vitesse et temps de centrifugation). La seconde purification partielle a été effectuée à partir de 250 mL de culture (pour chaque souche) à 37°C, après 20 heures d'induction de l'expression par [IPTG] 0,4 mM. Les cultures ont été centrifugées 15 min à 6000 g. Le culot a été lavé par 15 mL de LB frais, centrifugé 15 min à 6000 g et repris dans 7 mL de tampon de lyse (lysat). L'analyse sur gel d'électrophorèse sur polyacrylamide (11%) en conditions dénaturantes des échantillons obtenus à chaque étape est présentée dans les Figures 19 & 20 pour BL21(DE3) et B834(DE3)pLysS, respectivement. Les dépôts sur gel ont été de quantité identique (50 µg) pour tous les échantillons (excepté le surnageant de dialyse pour B834(DE3)pLysS : 12 µg). Les Figures 21A & 21B présentent l'intensité relative de la bande correspondant à la forme mature de l'HSP22 recombinante et la quantité de protéines totales, respectivement, pour chaque étape de la purification. Enfin, le Tableau I résume le protocole suivi, les résultats obtenus à chaque étape et présente le bilan de purification partielle. |
La lyse La quantité de protéines totales contenues dans le lysat est à peu près identique pour les deux souches : 142 mg et 126 mg pour BL21(DE3) et B834(DE3)pLysS, respectivement. Les conditions de lyse sont aussi efficaces dans un cas que dans l'autre. Cependant, le profil électrophorètique révèle une nette différence : la bande correspondant à la forme mature de l'HSP22 recombinante est très visible dans le cas de BL21(DE3) et a une intensité relative d'environ 16%. Dans le cas de B834(DE3)pLysS, elle est peu visible et a une intensité relative d'environ 0,5 %. Après centrifugation 40 min à 15000 g, on obtient le surnageant du lysat et le culot du lysat (qui est repris dans le tampon de lavage 1). Le culot contient environ les deux-tiers des protéines totales [97 mg des (97 + 43,5) mg totaux] dans le cas de BL21(DE3) mais l'intensité relative de la bande correspondant à la forme mature de l'HSP22 recombinante est à peu près la même que dans le surnageant (8,7 % et 7,8 %, respectivement). Dans le cas de B834(DE3)pLysS, le culot contient environ un quart des protéines totales [40 mg des (40 + 114) mg totaux] et l'intensité relative est 4 fois supérieure à celle du surnageant (9,1 % et 2,3 %, respectivement). Puisque c'est le culot qui est traité à chaque étape, il faut envisager une centrifugation plus longue et/ou à vitesse plus élevée dans le cas de BL21(DE3) pour augmenter la quantité de forme mature de l'HSP22 recombinante dans le culot. La contre-partie est d'entraîner la totalité des protéines et de ne pas accroître le degré de pureté à cette étape. |
Préparation des corps d'inclusion (1er lavage) Le culot repris dans le tampon de lavage 1 a été centrifugé 60 min à 21000 g. On obtient le surnageant de lavage 1 et un culot de lavage 1 (repris dans le tampon de lavage 2). Les conditions pour cette étape ne sont pas optimales dans le cas de BL21(DE3) :
Dans le cas de B834(DE3)pLysS, on aboutit aux mêmes conclusions, hormis ce qui concerne l'intensité relative ; cependant, elle est si faible (même nulle pour le surnageant) qu'il est difficile d'en tirer un enseignement. Préparation des corps d'inclusion (2ème lavage) Le culot repris dans le tampon de lavage 2 a été centrifugé 60 min à 21000 g. On obtient le surnageant de lavage 2 et un culot de lavage 2 (repris dans le tampon de dénaturation). Pour les deux souches, la quantité de protéines est cette fois plus élevée dans le culot que dans le surnageant. Il est malgré tout difficile de conclure pour cette étape puisque le culot de lavage 2 n'a pu être déposé sur gel d'électrophorèse du fait de la concentration élevée en chlorure de guanidine qui empêche une migration correcte. Remarque Les corps d'inclusion sont des particules qui contiennent la protéine sur-exprimée sous forme d'agrégats. Il n'est pas certain que les conditions de dosage des protéines par la méthode de Bradford les dénaturent et libèrent ainsi leur contenu protéique. Si les corps d'inclusion restent intègres, on sous-estime la quantité de protéines totales pour les étapes qui précèdent en ne dosant essentiellement que les protéines intrinsèques d'E. coli. |
Dénaturation des agrégats protéiques et renaturation de la protéine sur-exprimée Les corps d'inclusion (dans un premier temps) et les agrégats protéiques qu'ils contiennent (ensuite) sont dénaturés par le chlorure de guanidine (culot de lavage 2 repris dans le tampon de dénaturation) : après solubilisation de la membrane des corps d'inclusion, les chaînes polypeptidiques qui forment les agrégats se séparent les unes des autres. La renaturation est effectuée par dialyse extensive. La dialyse est une méthode simple qui préserve la structure des protéines. Elle présente une caractéristique supplémentaire extrêmement importante : la cinétique d'élimination de l'agent dénaturant est lente, ce qui augmente la probabilité de stabilisation de la structure des différents intermédiaires du repliement qui aboutissent à la forme native donc fonctionnelle d'une protéine. Cependant, certaines molécules ne suivent pas un chemin de repliement correct : l'exposition au solvant (polaire) de surfaces hydrophobes induit la formation de nouveaux agrégats protéiques. Il se forme alors un précipité dans le boudin de dialyse. Le dialysat a donc été centrifugé 10 min à 6000 g et on obtient le culot de dialysat et le surnageant de dialysat. C'est le surnageant qui cette fois nous intéresse puisqu'il contient les molécules solubles, donc celles qui se sont repliées (au moins partiellement) de façon correcte. Le surnageant du dialysat contient moins de protéines totales que le culot mais le résultat obtenu (quantité de protéines solubles, rendement et intensité relative non négligeables) indique que les conditions de renaturation, sans être encore optimales, ne sont pas mauvaises. |
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}