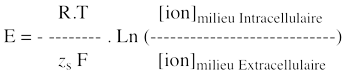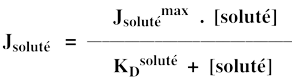| Tranports membranaires |
| Tweet |
|
Voir le cours sur les transports membranaires. Voir l'énoncé des exercices. |
Exercice N°1 : transport passif - equation de Ussing-Teorell Si la valeur théorique calculée avec l'équation de Ussing-Teorell est égale à la valeur de f (le quotient des flux transmembranaires mesurés expérimentalement avec des isotopes), le transport est passif. Si l'isotope I n’est pas chargée, le gradient de potentiel chimique ΔμI dépend de la différence de potentiel chimique transmembranaire de I : - ΔμI = RT ln (CExtracellulaireI / CIntracellulaireI) CIntracellulaireI et CExtracellulaireI sont les concentrations de I respectivement dans les compartiments intracellulaire et extracellulaire. Equation de Ussing-Teorell : f = exp (- ΔμI / RT)
=> - ΔμI = RT ln f = RT ln (CExtracellulaireI / CIntracellulaireI) => f = CExtracellulaireI / CIntracellulaireI Conclusion : la valeur théorique (CExtracellulaireI / CIntracellulaireI = 4) est quasiment identique au rapport des flux mesurés expérimentalement (f = 4,03), donc le transport est passif. |
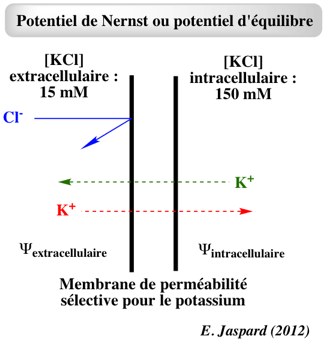
Exercice N°2 α. Rappel sur le potentiel d'équilibre pour un ion ou potentiel de Nernst E pour un ion Le milieux extracellulaire et le milieux intracellulaire sont séparés par une membrane. Ils contiennent KCl et la concentration dans le milieu intracellulaire est plus élevée.
Si la membrane n'est perméable qu'à K+, cet ion traverse la membrane du milieu intracellulaire (concentration plus élevée) vers le milieu extracellulaire (concentration plus faible). L'ion Cl- ne peut pas traverser la membrane.
Très rapidement, un équilibre s'établit où la différence de potentiel électrique déplace K+ vers le milieu intracellulaire à la même vitesse que la vitesse de diffusion de K+ vers le milieu extracellulaire en raison du gradient de concentration. Quand il n'y a plus de flux net de l'ion, il s'établit un équilibre thermodynamique. La différence de potentiel électrique à laquelle cet équilibre s'établit est appelée potentiel d'équilibre ou potentiel de Nernst (souvent noté E) pour l'ion considéré :
Remarque : cette expression est équivalente à ΔΨM = ΨI - ΨE = - [RT / zs . F] . ln (CsI / CsE) La valeur du potentiel d'équilibre d'un ion dépend donc du gradient de concentration de cet ion au travers de la membrane :
|
β. Correction de l'exercice N°2
Par conséquent, après un déplacement microscopique de K+, on arrive presque instantanément à une situation d’équilibre qui se traduit par : ΔG' ≈ ΔμK+ElectroChimique = RT ln ([K+]intracellulaire / [K+]extracellulaire) + (zK+ . F . EK+) = 0 zK+ . F . EK+ = - RT ln ([K+]Intracellulaire / [K+]Extracellulaire) Potentiel d’équilibre ou potentiel de Nernst : EK+ = - (RT / zK+ . F) ln ([K+]Intracellulaire / [K+]Extracellulaire) = - 58 mV |
Exercice N°3 : Potentiel d’équilibre ou potentiel de Nernst a) Le mouvement d’un ion à travers une membrane perméable est dû :
La différence de potentiel électrochimique de l’ion K+ est donné par la relation de Nernst : ΔG' ≈ µAK+ - µBK+ = RT . ln ([K+]compartiment A / [ K+]compartiment B) + [zK+ . F . (EA - EB)K+] À l’équilibre électrochimique, cette différence de potentiel électrochimique est nulle. Donc : (EA - EB)K+ = - (RT / zK+ . F ) . ln ([K+]compartiment A / [ K+]compartiment B) (EA - EB)K+ = - (8,315 x 310,3 / 1 x 96485) . ln (0,1 / 0,01) = - 62 mV b) À l’équilibre, le flux net de l’ion K+ est nul. |
Exercice N°4 : Potentiel d’équilibre ou potentiel de Nernst 1er cas
2ème cas
3ème cas
Conclusion sur la participation des ions à l’établissement d’un potentiel transmembranaire :
|
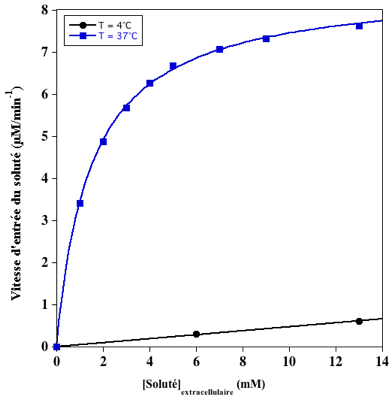
Exercice N°5 : Diffusion simple vs. transport facilité - Transport actif secondaire 1. Interprétation des résultats
Cellules incubées à 4°C :
Cellules incubées à 37°C :
Jsolutémax est le flux maximal d'entrée du soluté dans la cellule (asymptote de l'hyperbole) : Jsolutémax ≈ 8 µM.min-1.
|
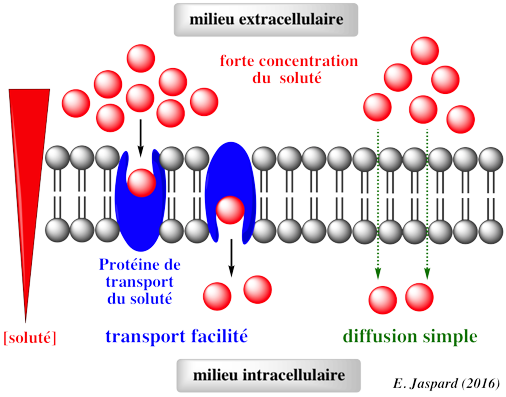
2. Illustration des processus
Exemple de transport facilité : le glucose par un transporteur de la famille GLUT.
|
3. Energie libre de Gibbs associée au transport facilité du glucose ΔG' = RT . Ln (CGlucoseIntracellulaire / CGlucoseExtracellulaire) = 1,987 . 310,3 . ln (0.5 10-3 / 5 10-3) = - 1.42 kcal.mol-1
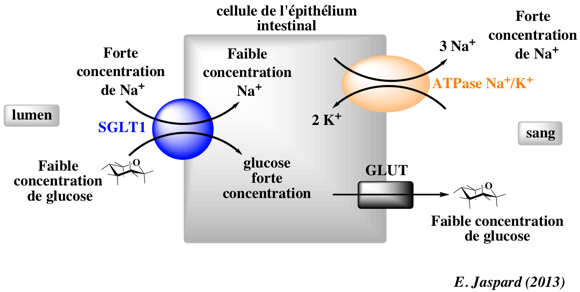
4. Energie libre de Gibbs nécessaire au transport actif du glucose Il faut un système qui pompe le glucose du liquide interstitiel dans les tubules ([glucose]tubule = 0,005 mM) puis le transporte au travers de la membrane plasmique jusqu'au sang ([glucose]sang = 5 mM). Le gradient de concentration au travers duquel le glucose doit être pompé est donc : 0,005 mM => 5 mM. Il faut un système de transport actif qui compense une énergie minimale : ΔG' = 1,987 . 310,3 . ln (5 10-3 / 0.005 10-3) = + 4.26 kcal.mol-1 5. Cette énergie est apportée par un système de transport actif secondaire :
6. Les 2 sources d'énergie pour déplacer une mole de glucose émanent de la composante chimique (gradient de concentration) et de la composante électrique (différence de potentiel électrique : ΔΨM = - 70 mV) puisque l'ion Na+ est chargé : ΔG' = ΔG'ChimiqueNa+ + ΔG'ElectriqueNa+ = RT . Ln (CNa+Intracellulaire / CNa+Extracellulaire) + ( zNa+ . F . ΔΨM ) ΔG' = [1,987 . 310,3 . ln (10 10-3 / 140 10-3)] + [+1 . 23060 . (- 70 10-3)] = - 3,25 kcal.mol-1
Conclusion : l'énergie fournie (ΔG' = - 3,25 kcal.mol-1) par le transport d'1 ion Na+ n'est pas suffisante => 2 ions Na+ doivent être transportés pour transporter une molécule de glucose. |
![]()