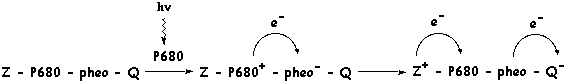| Photosystèmes I et II - Antenne, paire spéciale, séparation de charges |
| Tweet |
|
|
1. Description générale des photosystèmes 2. La chute dans le rouge et la résonance 3. La séparation des charges positive et négative 4. Les donneurs primaires d'électrons |
5. Résumé des évènements au sein du photosystème II (PSII - P680) 6. Le photosystème I (PSI - P700) 7. Complexes CPC ou complexes d'antenne secondaires 8. Liens Internet et références bibliographiques |
|
1. Description générale des photosystèmes Deux types de complexes protéines-pigments appelés respectivement :
représentent les unités fonctionnelles au sein desquelles se déroulent les réactions photoinduites de la photosynthèse. Les 2 photosystèmes sont enfouis sur toute l'épaisseur de la membrane thylacoïde, dans des régions différentes :
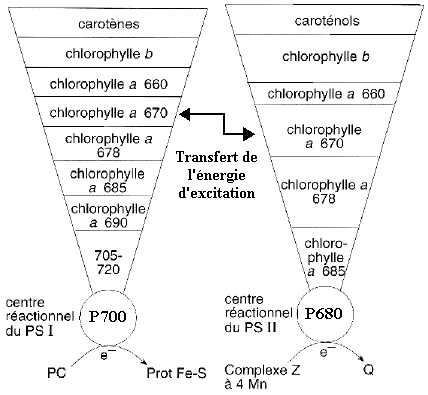
Chaque photosystème comporte un lit de pigments (60 à 2000 molécules selon l'organisme) appelé complexe d'antenne. Chaque photosystème contient un second complexe d'antenne volumineux qui sert de collecteur de lumière : le complexe photocollecteur couplé aux chlorophylles a/b ou complexe CPC ou complexe d'antenne secondaire mobile. La figure ci-dessous indique la distribution des divers pigments (appelés pigments accessoires) dans les complexes d'antenne de chaque photosystème des végétaux verts.
Source : "Physiologie végétale" (1995) - Laval-Martin & Mazliak Les chiffres indiquent la longeur d'onde d'absorbance maximale de chacun des pigments. Ces longueurs d'onde se situent dans le rouge du spectre de la lumière visible. Le coeur du processus photochimique au sein de chaque photosystème est un complexe protéique appelé centre réactionnel qui contient 2 molécules de chlorophylle a formant la paire spéciale. Ces 2 molécules de chlorophylle a se distinguent des autres molécules de chlorophylle car elle sont le foyer vers lequel converge l'énergie d'un photon captée par le complexe d'antenne. Le centre réactionnel d'un photosystème est désigné d'après la longueur d'onde du pic d'absorption de sa paire spéciale :
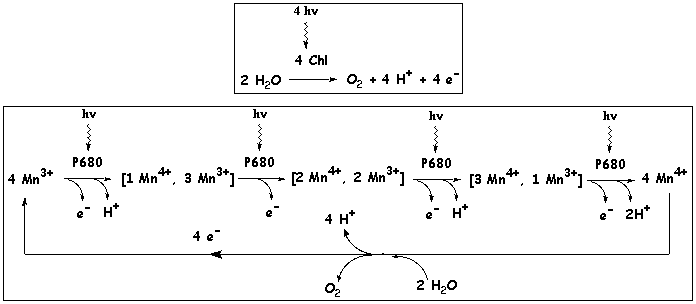
Les donneurs d'électrons sont respectivement : la plastocyanine (PC) et le complexe Z à 4 ions manganèse |
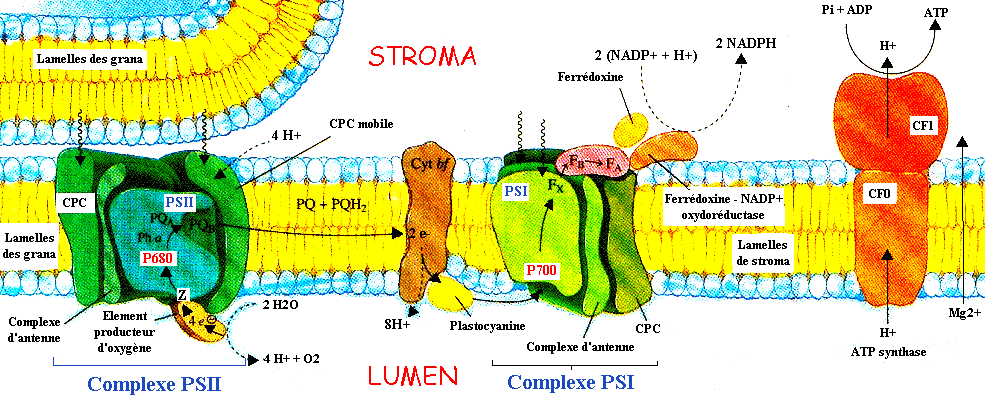
| Les 2 photosystèmes
fonctionnent en série : comme ils se situent dans des régions
différentes de la membrane
thylacoïde, ils sont en relation par l'intermédiaire
de transporteurs
d'électrons particuliers.
La membrane thylacoïde contient d'autres composants actifs de la photosynthèse :
Adapté de : "Principes de Biochimie" Horton et al. (1994) |
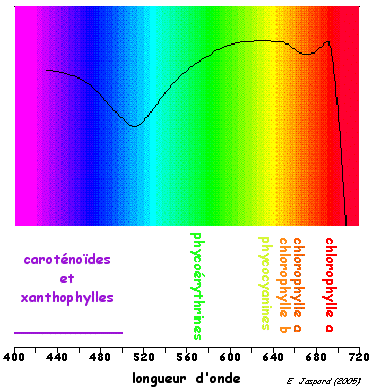
2. La chute dans le rouge et la résonance On observe une chute brutale de la photosynthèse à partir de 680 nm bien que la lumière soit encore fortement absorbable par les chlorophylles. Ce phénomène est appelé "chute dans le rouge" ("red drop"). L'activité photosynthétique peut être restaurée quand les photons de longueur d'onde inférieure à 680 nm sont ajoutés à l'éclairement de 680 nm. L'inefficacité des photons de longueur d'onde supérieure à 680 nm dans le phénomène photosynthétique est dû à l'absence de chlorophylle dans PSII apte à absorber ces longueurs d'onde. Remarques :
Adapté de : "Photosynthesis" |
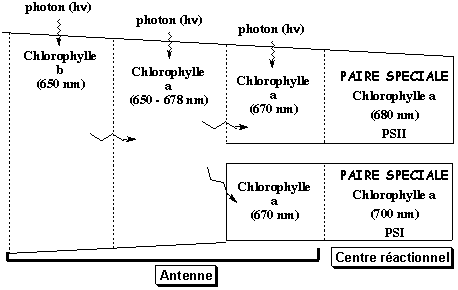
Les complexes d'antenne photosynthétiques ont donc pour rôle d'augmenter la probabilité des réactions photochimiques en "piègeant" l'énergie des photons et en la transmettant de molécule de chlorophylle en molécule de chlorophylle sans perte d'énergie grâce au phénomène de résonance.
Adapté de : "Physiologie végétale" (1995) - Laval-Martin & Mazliak Cette énergie est ainsi canalisée jusqu'à la paire spéciale de chlorophylle a du centre réactionnel.
Ces distances soulignent l'optimisation de leur organisation pour assurer un transfert d'énergie parfait. |
|
3. La séparation des charges positive et négative La chlorophylle peut adopter divers états d'excitation. En ordre d'énergie croissante par rapport à l'état fondamental, ces niveaux sont : le premier état singulet excité / l'état triplet métastable / le second état singulet excité / l'état triplet excité C'est seulement à partir du premier état singulet ou de l'état triplet métastable que le retour de l'électron à l'état fondamental peut fournir une énergie qui permet les réactions photochimiques de la photosynthèse. Ces réactions ne sont possibles que si la différence d'énergie momentanément stabilisée dure plus de 15 10-9 s. Cette stabilisation s'opère par un mécanisme de séparation des charges positive et négative entre :
Ces molécules sont étroitement associées aux chlorophylles a (Chl a) des centres réactionnels P700 et P680. |
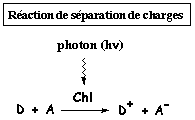
| A l'obscurité, les électrons de D, A et de Chl sont à l'état fondamental et la combinaison peut s'écrire : | D - Chl - A |
| Si, à la lumière un photon est absorbé par Chl, il délocalise l'électron de Chl au premier état singulet excité et la combinaison devient : | D - Chl* - A |
| Instantanément et simultanément, l'électron de D est cédé à Chl* et l'électron de Chl* réduit A. La combinaison devient : | D+ - Chl - A- |
La séparation des charges positive et négative est stabilisée entre D+ et A- pour une durée de 10-10 s à 10-3 s.
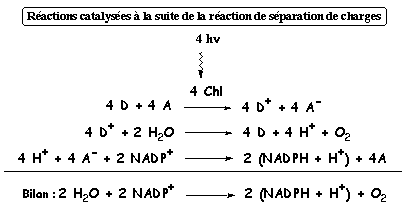
Au cous de ce processus, la Chl n'a été oxydée en Chl* que pendant le temps trés court et habituel de 10-15 s et a joué le rôle de catalyseur pour les réactions mentionnées dans la figure ci-dessous.
Adapté de : "Physiologie végétale" (1995) - Laval-Martin & Mazliak |
|
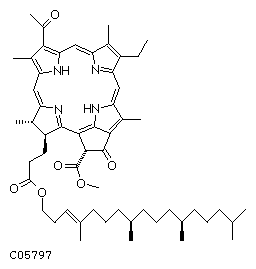
b. La plastoquinone QA Après excitation par un photon, la chlorophylle a transmet immédiatement cette excitation à une phéophytine a (pheo). A la diffèrence de la Chl a qui contient de 2 ions magnésium, la phéophytine a contient 2 protons (figure ci-dessous).
Source : KEGG La phéophytine a cède 1 électron à l'accepteur primaire, la plastoquinone QA (fortement attachée à un polypeptide de PSII) selon le mécanisme décrit dans la figure ci-dessous.
Comme dans le cas de l'ubiquinone des mitochondries, QA est réduite par 2 transferts mono-électroniques successifs. A son tour, QA cède ses électrons à un accepteur secondaire QB, une plastoquinone attachée de façon réversible à un polypeptide de PSII. |
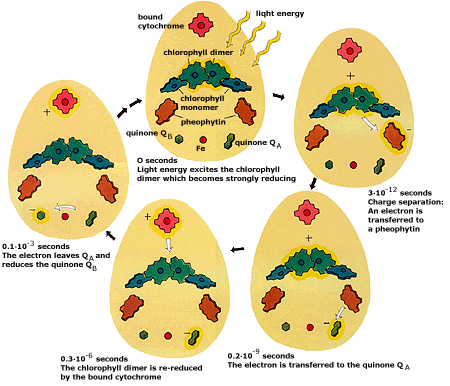
5. Résumé des évènements au sein du photosystème II (PSII - P680) J. Deisenhofer, R. Huber, H. Michel ont reçu le Prix Nobel en 1988 pour la détermination de la structure d'une protéine membranaire aussi complexe que celle du centre réactionnel P680 de la bactérie Rhodopseudomonas virdis. Cette structure a permi d'établir l'ordre des évènements et les temps pour chacun d'entre eux (figure ci-dessous). Elle illustre la notion de stabilisation de la différence d'énergie qui s'opère par le mécanisme de séparation des charges.
|
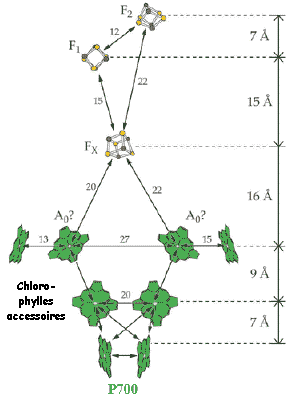
| 6. Le photosystème I (PSI - P700) Le donneur primaire d'électrons de PSI est la plastocyanine, protéine de 11 kDa liée à la membrane et qui contient du cuivre. Elle reçoit les électrons d'un complexe cytochrome b563/f lié à un quinol et qui contient un centre [Fe - S] de type "de Rieske". L'accepteur primaire d'électrons de PSI est un ensemble constitué d'une chlorophylle réduite Chl- et d'un composé X. Ce premier accepteur est suivi du P430, constitué de 2 protéines (FA et FB : centres [Fe-S]). Deux protéines extrinsèques interviennent ensuite dans le transfert final des électrons :
La figure ci-dessous monte un modèle pour l'organisation des transporteurs d'électrons au sein du PSI.
Source : "Photosystem I" - "Plant Physiology Online"
|
|
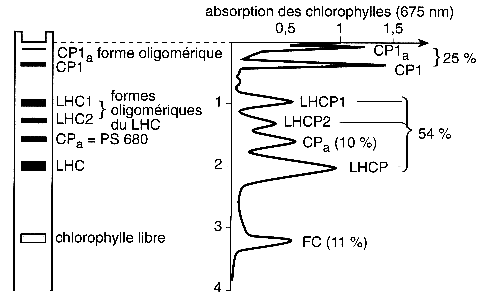
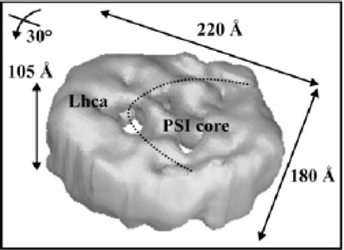
7. Complexes CPC ou complexes d'antenne secondaires Les photosystèmes PSI et PSII contiennent chacun un second complexe d'antenne. Ces complexes d'antennes périphériques appartiennent à la classe des complexes photocollecteurs couplés aux chlorophylles a/b ("Light Harvesting Complexes I and II" : LHCI pour PSI et LHCII pour PSII). Ces complexes servent de collecteur de lumière pour chaque photosystème : les plantes peuvent ainsi s'adapter aux changements de caractéristiques du spectre de la lumière absorbée en bougeant ces antennes mobiles du PSII au PSI (ce que l'on appelle "l'état de transition" - voir la figure 1 de Haldrup et al. (2001). La partie gauche de la figure ci-dessous montre la séparation des LHCI et LHCII par électrophorèse sur gel de polyacrylamide en conditions dénaturantes. A droite sont indiqués les pics d'absorbance à 675 nm de ces complexes et leur pourcentage en chlorophylles.
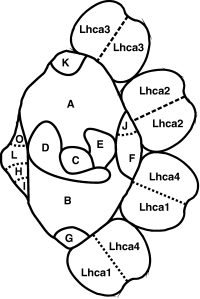
Source : "Physiologie végétale" (1995) - Laval-Martin & Mazliak Le PSI assure le transfert des électrons de la plastocyanine (lumen de la membrane thylakoïde) à la ferrédoxine (stroma). Chez les plantes supérieures, PSI est un complexe multiprotéique qui forme (figure ci-dessous) :
Source : Jensen et al. (2003)
Les protéines du "coeur" fixent :
Source : Kargul et al. (2003) Chaque protéine LHCa de l'antenne périphérique fixe environ une dizaine de molécules de Chl a , 2 Chl b, 2 caroténoïdes
PSI fixe également LHCII dans l'état 2 de l'état de transition. |
| 8. Liens Internet et références bibliographiques |
|
"Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X "Physiologie végétale" (1995) volume I - "Nutrition et métabolisme", Danielle Laval-Martin et Paul Mazliak, Collection "Méthodes", Hermann / ISBN : 2 7056 6253 7 |
|
| "Photosystem I" - "Plant Physiology Online" | Aller au site |
|
Schubert et al. (1997) "Photosystem I of Synechococcus elongatus at 4 A resolution: comprehensive structure analysis" J. Mol. Biol. 272, 741 - 69 Jensen et al. (2003) "Molecular dissection of photosystem I in higher plants: topology, structure and function" Physiologia Plantarum 119, 313 -321 Iwai et al. (2008) "Molecular Remodeling of Photosystem II during State Transitions in Chlamydomonas reinhardtii" The Plant Cell 20, 2177 - 2189 |
|
|
Haldrup et al. (2001) "Balance of power: a view of the mechanism of photosynthetic state transitions" Trends Plant. Sci. 6, 301 - 305 Jensen et al. (2003) "Molecular dissection of photosystem I in higher plants: topology, structure and function" Physiol. Plantarum 119, 313 - 321 Kargul et al. (2003) "Three-dimensional Reconstruction of a Light-harvesting Complex I- Photosystem I (LHCI-PSI) Supercomplex from the Green Alga Chlamydomonas reinhardtii" J. Biol. Chem. 278, 16135 - 16141 |
|
![]()