| La chaîne de transfert d'électrons des mitochondries des végétaux |
| Tweet |
|
|
1. Introduction 2. Caractéristiques générales des complexes protéiques de la chaîne respiratoire 3. Mécanismes du transfert des électrons
|
4. Enzymes supplémentaires spécifiques de la chaîne respiratoire chez les végétaux
5. Bilan - résumé de la chaîne de transport d'électrons 6. Liens Internet et références bibliographiques |
|
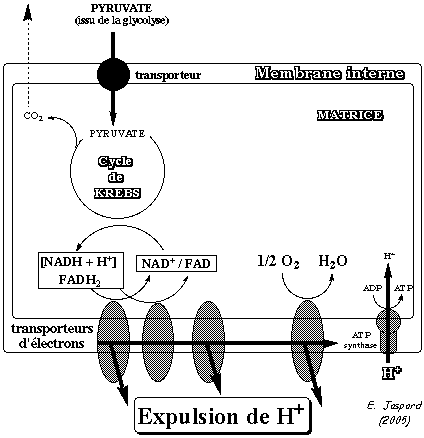
1. Introduction La mitochondrie est limitée par deux membranes de propriétés très différentes :
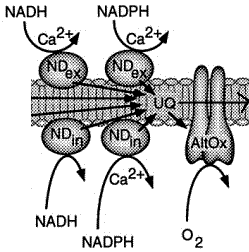
La chaîne respiratoire ou chaîne de transport d'électrons (figure ci-contre) est localisée dans la membrane interne des mitochondries. Chaque mitochondrie contient des milliers d'exemplaires de la chaîne de transport d'électrons. |
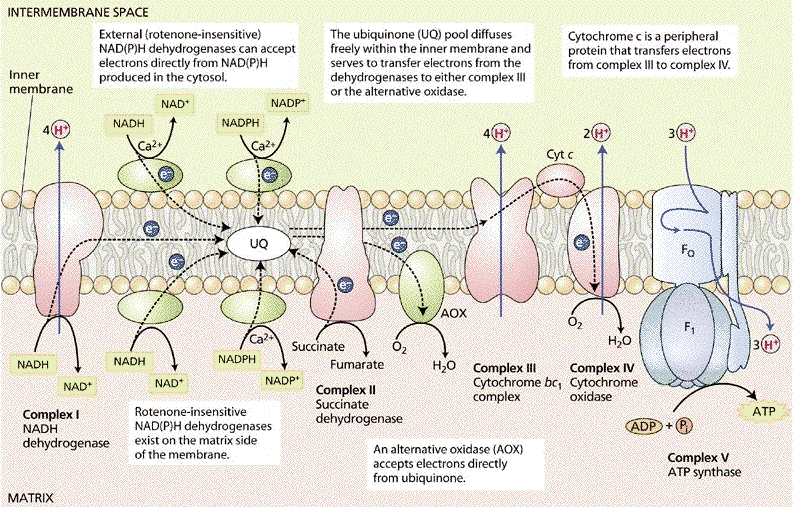
Figure ci-dessous : représentation de la chaîne de transport d'électrons chez les végétaux
Source : "Plant Physiology Online" Elle contient des complexes supplémentaires par rapport aux animaux :
|
| Règne | Complexe ou protéine | Sous-unités | Masse molaire | Composants | Transfert | Inhibiteurs |
| Animaux et végétaux |
NADH - coenzyme Q |
25 | 800 kDa | 1 FMN 22 à 24 atomes [Fe - S] dans 5 à 8 centres |
électrons |
roténone amytal |
|
succinate
- coenzyme Q |
4 | 125 kDa | 1 FAD 7 à 8 atomes Fe - S dans 3 centres cytochrome b560 | électrons | malonate | |
| coenzyme Q | 1 | 0,86 | lipide isoprénoide |
électrons |
-------- | |
| Spécifiques aux végétaux | NAD(P)H déshydrogénases | 4 protéines : 2 NDint et 2 NDext cytochrome b560 | électrons | diphénylèneiodonium (DPI) insensibles à la roténone | ||
| oxydase alternative (AOX) | 2 | 64 kDa | électrons | SHAM propylgallate insensible au KCN | ||
| Animaux et végétaux |
coenzyme Q
- cytochrome c |
8 | 220 kDa | 2 centres [Fe - S] cytochrome b560 cytochrome b566 cytochrome c1 |
électrons |
antimycine |
| cytochrome C | --- | 12 kDa | hème (Fe) | électron | ||
|
cytochrome c oxydase |
12 | 200 kDa |
cytochrome a
cytochrome a3 |
électrons |
KCN / CO / NaN3 |
| 3. Mécanismes du transfert des électrons
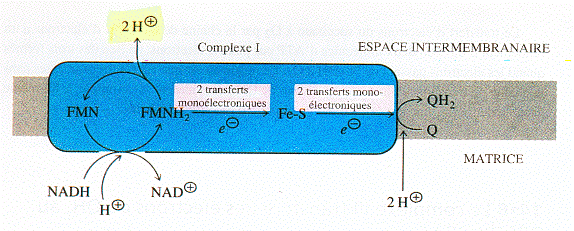
a. Le complexe I : NADH - coenzyme Q oxydoréductase Le complexe I catalyse le transfert de 2 électrons du NADH (formé au cours du cycle de Krebs) au coenzyme Q via la flavine mononucléotide et un ensemble de centres [Fe - S].
Source : "Principes de Biochimie " Horton et al. (1994) |
|
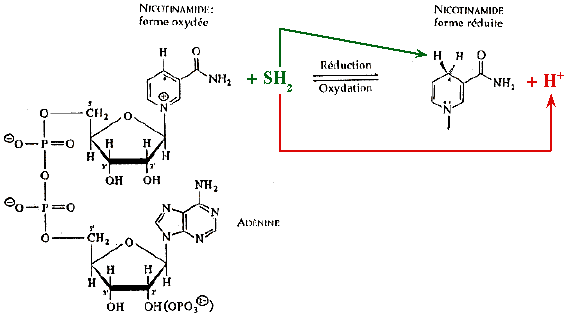
Le NADH cède ses électrons par paire (sous forme d'ion hydrure, H-). La réaction d'oxydo-réduction du NAD(P)+ concerne le groupe nicotinamide :
Ce transfert est un exemple remarquable de la stéréospécificité des réactions chimiques catalysées par les enzymes déshydrogénases.
Source : "Principes de Biochimie " Horton et al. (1994) Puis le complexe I catalyse la réduction de la flavine mononucléotide (FMN) par le (NADH + H+) selon les réactions : +H+, + H- -H+ , - e- -H+ , - e- |
|
De nombreuses protéines appelées flavoenzymes comportent un groupe prosthétique qui participe au mécanisme catalytique :
Le centre réactionnel est un noyau aromatique tricyclique, l'isoalloxazine (la partie en rouge). Source : "Principes de Biochimie" Horton et al. (1994) |
|
|
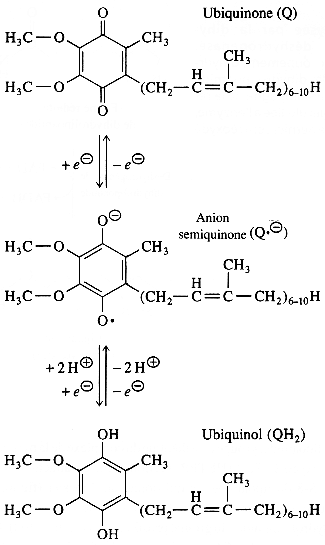
La FMN est donc une sorte de "convertisseur" d'un flux bi-électronique en un flux mono-électronique, puisqu'elle cède ses électrons à des centres [Fe - S], accepteurs mono-électroniques. Ces centres [Fe - S] cèdent à leur tour cet électron au coenzyme Q ou ubiquinone qui est réduit en passant par un intermédiaire semi-quinone (figure ci-dessous) selon les réactions : +e- +2H+ , + e- Le coenzyme Q possède trois niveaux d'oxydation :
Source : "Principes de Biochimie" Horton et al. (1994) Le coenzyme Q est une benzoquinone qui porte 4 substituants dont une longue chaîne hydrophobe. Cette particularité structurale lui confère une grande solubilité dans les lipides. En conséquence les trois formes Q, Q.- et QH2 diffusent d'une face à l'autre de la membrane interne de la mitochondrie. Le coenzyme Q est un transporteur mobile de la chaîne respiratoire. Lors du transfert des électrons au sein du complexe I, 3 ou 4 protons (selon les données expérimentales) sont expulsés de la matrice vers l'espace intermembranaire. |
|
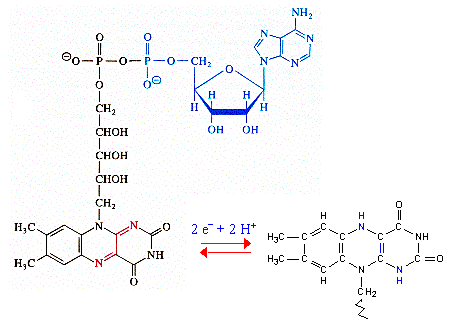
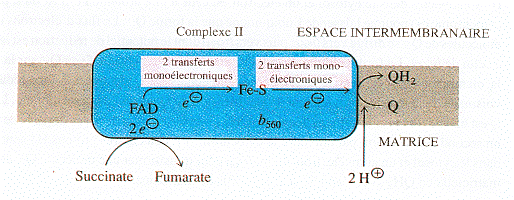
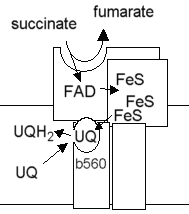
b. Le complexe II : succinate - coenzyme Q oxydoréductase Le complexe II accepte deux électrons du succinate et catalyse la réduction du coenzyme Q en QH2, le succinate étant transformé en fumarate (c'est l'une des réactions du cycle de Krebs).
Source : "Principes de Biochimie" Horton et al. (1994) La réaction met en jeu le FAD, des centres [Fe - S] et le cytochrome b560 attaché au complexe II.
|
|
Le complexe II joue un rôle important puisque c'est par lui qu'entrent les électrons :
La variation d'énergie libre de la réaction est faible du fait d'une faible différence de potentiel de réduction entre les donneurs et les accepteurs d'électrons. En conséquence, le complexe II ne contribue pas à l'expulsion de protons de la matrice vers l'espace intermembranaire. |
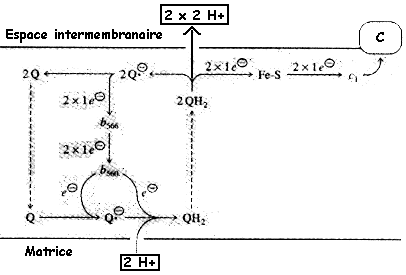
c. Le complexe III : coenzyme Q - cytochrome c oxydoréductase Le complexe III est constitué entre autre :
Les électrons sont cédés par le complexe III au cytochrome c (différent du cytochrome c1) qui les amène au complexe IV.
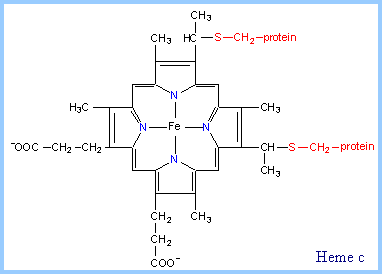
Adapté de : "Principes de Biochimie" Horton et al. (1994) Le trajet des électrons n'a été résolu que quand un trajet circulaire, appelé cycle Q (figure ci-contre), fût suggéré par Peter Mitchell puis détaillé entre autres par Bernard Trumpower. Le coenzyme Q et QH2 diffusent d'une face à l'autre de la membrane mitochondriale interne et l'ensemble des deux hèmes du cytochrome b occupe toute l'épaisseur de cette membrane. Le cytochrome c est un autre transporteur mobile de la chaîne respiratoire. Bilan du cycle Q : coenzyme QH2 + 2 cyt cox + 2 H+ mat ---> coenzyme Q + 2 cyt cred + 2 H+ eim L'hème est un groupe prosthètique des cytochromes. Il contient un atome de fer au sein d'un noyau porphyrine. L'atome de fer est lié à 4 atomes d'azote du noyau porphyrine. |
Seul l'hème du cytochrome c est covalemment lié à des résidus cystéine via une liaison thioether (figure ci-contre). Les hèmes des 3 classes de cytochrome (a, b, c) diffèrent légèrement au niveau des substituants du noyau porphyrine. Par exemple, l'hème du cytochrome a possède une longue chaîne farnesyle qui inclue 3 unités isoprènoides. Source : "Electron Transfer Chain" |
|
|
La variation d'énergie libre liée au parcours des électrons au sein du complexe III est légèrement plus faible que celle générée par le complexe I. Cependant, le complexe III contribue à l'expulsion de (2 x 2) protons de la matrice vers l'espace intermembranaire. |
|
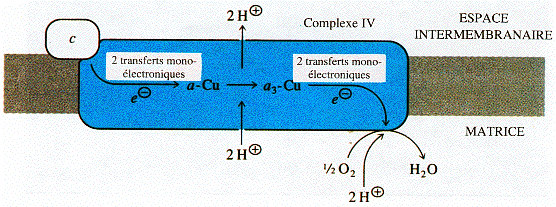
d. Le complexe IV : cytochrome c oxydase Le complexe IV est constitué entre autre :
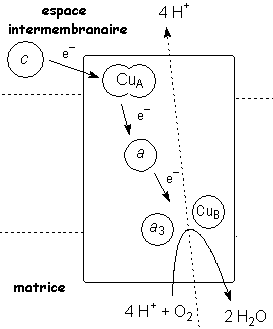
Source : "Principes de Biochimie" Horton et al. (1994) Le complexe IV est le dernier de la chaîne de transport d'électrons. Il catalyse la réduction de l'oxygène moléculaire en eau : O2 + 4 H+ + 4 e- ---> 2 H2O Les 4 électrons sont transferés un à un du cytochrome c au complexe IV. L'ordre de transfert des électrons est donc : cyt c ---> CuA ---> hème a ---> [hème a3/CuB]
Source : "Mitochondrial Electron Transport Chain" Quand les électrons traversent les cytochrome a et a3, les atomes de cuivre des hèmes changent d'état d'oxydation. Les hèmes des cytochromes du complexe IV et les 2 atomes de cuivre CuA et CuB sont ligandés à des atomes d'azote de résidus histidine. Le complexe IV expulse 2 protons de la matrice vers l'espace intermembranaire. Par ailleurs, il contribue d'autant plus à la formation du gradient de concentration de protons que, du fait de la formation de molécule d'eau, il soustrait des protons de la matrice. |
Ces 2 déshydrogénases constituent un contournement du complexe I et elles sont insensibles à la roténone (inhibiteur de ce complexe). Par voie de conséquence, ces réoxydations se substituent aux systèmes de "navette" que l'on trouve chez les animaux. |
Source : I. Moller (1997) |
|
Les NAD(P)H déshydrogénases n'expulsent pas de protons puisqu'elles "court-circuitent" le premier site d'expulsion des protons (le complexe I). |
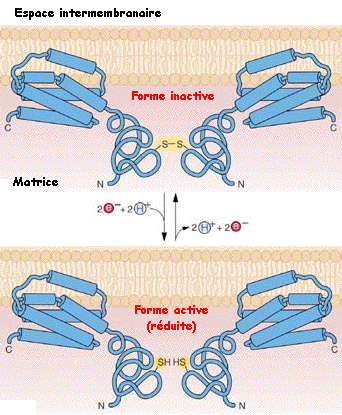
b. L'oxydase alternative ou AOX La chaîne respiratoire des végétaux possède une seconde oxydase terminale (contrairement aux animaux) : l'oxydase alternative ou AOX (insensible au cyanure). Cette voie alternative transfère les électrons des quinones à l'oxygène et contourne ainsi 2 sites d'expulsion de protons. Par voie de conséquence, elle contribue peu (voire pas) à la production finale d'ATP. L'AOX a pour rôle :
En conditions normales, l'AOX est faiblement transcrite dans les mitochondries des végétaux. En revanche, son expression augmente en conditions de stress (froid, sécheresse, pathogènes, ...).
Figure ci-dessus : structure et mode de régulation de l'AOX adaptée de "Plant Physiology Online". |
|
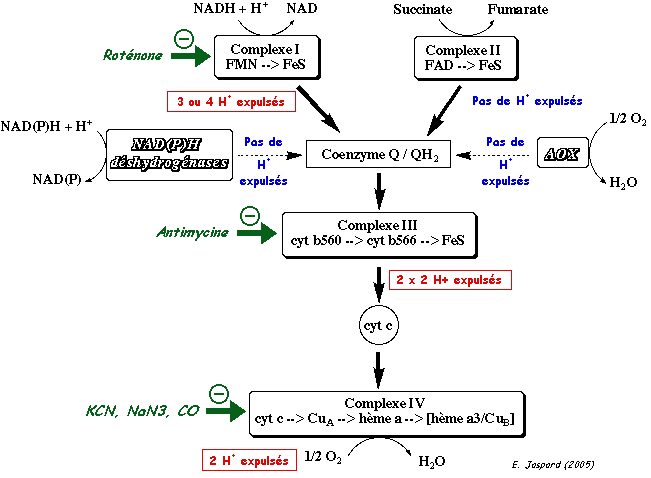
5. Bilan - résumé de la chaîne de transport d'électrons La figure ci-dessous résume l'ensemble des réactions de la chaîne de transport d'électrons des mitochondries des végétaux.
Dans l'enchaînement des réactions d'oxydo-réduction qui ont lieu lors du transfert des électrons de l'ensemble de la chaîne de transport d'électrons , si l'on ne considère que :
Les deux demi-réactions rédox sont :
|
|
D'après la relation qui lie la différence de potentiel de réduction standard, ΔE°'réaction, à la variation d'énergie libre standard, ΔG°'réaction : ΔG°'réaction = - n . F . ΔE°'réaction = - (2 . 96500 . 1,14) = - 220 kJ.mol-1 Puisque la variation d'énergie libre standard de la synthèse de l'ATP est environ +30,5 kJ.mol-1, la variation d'énergie libre standard liée à la réoxydation du (NADH + H+) devrait, en théorie, permettre la synthèse de : (220 kJ.mol-1 / 30,5 kJ.mol-1) = 7 molécules d'ATP par molécule de (NADH + H+) réoxydé. En fait ce sont 3 molécules d'ATP qui sont synthétisées.
|
| 6. Liens Internet et références bibliographiques |
| "Plant Physiology Online" | Aller au site |
| Eubel et al. (2004) "Respiratory chain supercomplexes in plant mitochondria" Plant Physiol. and Biochem. 42, 937 - 942 | Article |
| "Complex I - Home page" | Aller au site |
| "Electron Transfer Chain" | Aller au site |
|
I. Moller (1997) "The oxidation of cytosolic NAD(P)H by external NAD(P)H dehydrogenases in the respiratory chain of plant mitochondria" Physiol. Plant. 100, 85 "Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X |
|
![]()