| Oxydation de l'acétyl CoA : le cycle de l'acide citrique (cycle de Szent-Györgyi & Krebs) |
| Tweet |
|
|
1. Introduction 2. Différentes représentations des réactions du cycle de Krebs 3. Spécificité du cycle de Krebs chez les végétaux : le cycle du glyoxylate |
4. La succinyl-CoA synthétase 5. Régulation du cycle de Krebs 6. Liens Internet et références bibliographiques |
1. Introduction Le cycle de Krebs (appelé aussi cycle des acides tricarboxyliques ou cycle du citrate) est l'un des mieux décrits. Il inclue des réactions et des métabolites élucidés par Albert Szent-Györgyi (Prix Nobel 1937), en particulier la catalyse de l'acide fumarique. Toutes les enzymes qui constituent ce cycle ont été cristallisées et ont fait l'objet de "Molecule of the month" en octobre 2012 ("Protein Data Bank"). Le cycle de Krebs est une "plaque tournante" du métabolisme aérobie. En effet :
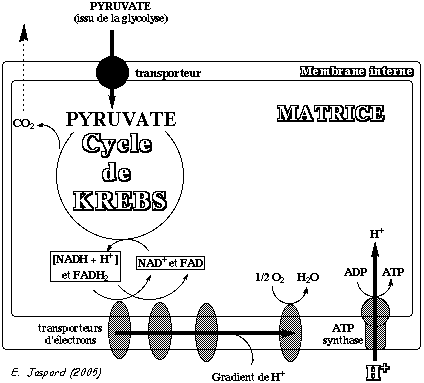
Localisation cellulaire : les enzymes cycle de Krebs sont situées dans le cytosol des Procaryotes et dans la matrice des mitochondries des Eucaryotes.
Principe du cycle :
Dans les cellules des organismes aérobies, le pyruvate formé à l'issue de la glycolyse est oxydé en CO2 et H2O par une série de réactions enzymatiques au cours desquelles une partie de l'énergie est finalement stockée sous forme d'une molécule à haut potentiel énergétique : la molécule d'ATP. Cette partie de l'énergie ainsi stockée et utilisable par la cellule (au contraire de l'énergie dissipée sous forme de châleur) s'appelle l'énergie libre de Gibbs. Le pyruvate est d'abord converti en acétyl coenzyme A (acétyl-CoA) à partir du coenzyme A (CoASH). L'oxydation qui s'en suit du groupe acétyle de l'acétyl CoA s'effectue dans le cycle de Krebs. L'énergie issue des réactions d'oxydation de ce cycle est convertie sous forme de coenzymes réduits qui contiennent une partie de l'énergie initiale du glucose sous forme d'électrons (pouvoir réducteur) :
On doit à Hans Adolf Krebs et ses collaborateurs l'élucidation des réactions de ce cycle (Prix Nobel 1953). On peut rappeler que Hans Krebs a aussi décrit les esquisses du cycle de l'urée (1933). Il a aussi proposé le cycle du glyoxylate qui est une variante du cycle du citrate (Hans Krebs et Hans Kornberg, 1957). |
|
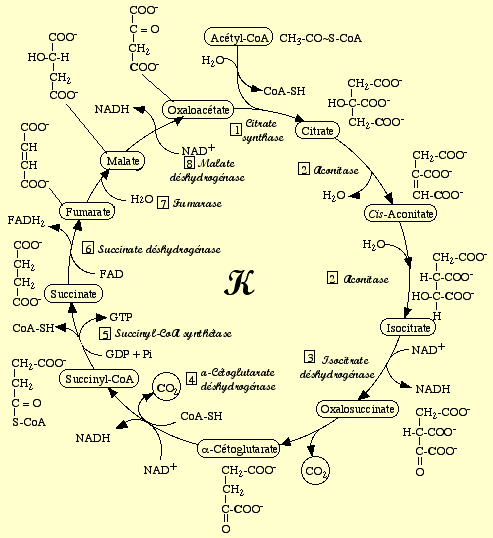
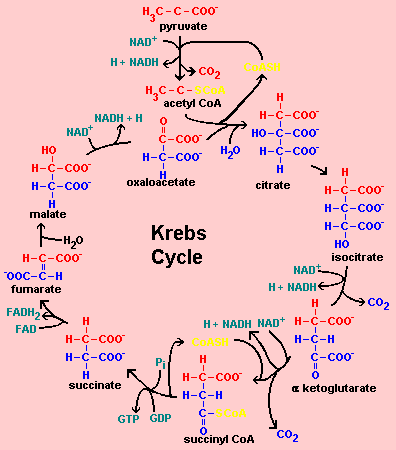
2. Différentes représentations des réactions du cycle de Krebs PLANTES
Source : Cycle de Krebs - Université Jussieu - Paris ANIMAUX
Source : R. Huskey - Université de Virginie |
| Synthèse du cycle de Szent-Györgyi & Krebs | ||||||
| Réaction | Substrats | Produits | Enzyme | EC | Commentaires | |
| 1 | oxaloacétate + acétyl-CoA + H2O | citrate + CoA-SH | citrate synthétase | EC 2.3.3.1 | irréversible | Acétyl-CoA : liaison thioester / intermédiaire : cytroyl-CoA. |
| 2 | citrate | cis-aconitate + H2O | aconitate hydratase | EC 4.2.1.3 | réversible | La même enzyme catalyse ces 2 étapes. |
| 3 | cis-aconitate + H2O | isocitrate | réversible | |||
| 4 | isocitrate + NAD+ | oxalosuccinate + NADH + H+ | isocitrate déshydrogénase | réversible | L'isocitrate déshydrogénase à NAD+ nécessite Mn2+ et Mg2+. |
|
| 5 | oxalosuccinate | α-cétoglutarate + CO2 | irréversible | Même enzyme que l'étape précédente. |
||
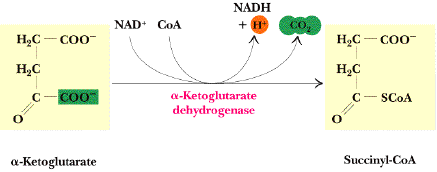
| 6 | α-cétoglutarate + NAD+ + CoA-SH | succinyl-CoA + NADH + H+ + CO2 | α-cétoglutarate déshydrogénase | complexe multi-enzymatique | irréversible | C'est la même réaction que la transformation du pyruvate en acétyl-CoA. |
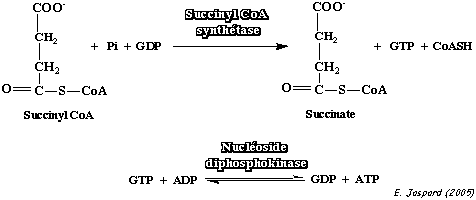
| 7 | succinyl-CoA + GDP + Pi | succinate + CoA-SH + GTP | succinyl-CoA synthétase | réversible | Chez l'homme, une isoforme utilise le GDP (pour former du GTP) et l'autre l'ADP (pour former de l'ATP) . |
|
| 8 | succinate + ubiquinone (Q) | fumarate + ubiquinol (QH2) | succinate déshydrogénase | EC 1.3.1.5 | C'est la sous-unité (flavoprotéine) du complexe II de la chaîne respiratoire. |
|
| 9 | fumarate + H2O | L-malate | fumarase | EC 4.2.1.2 | réversible | Formation d'un intermédiaire carbanion. |
| 10 | L-malate + NAD+ | oxaloacétate + NADH + H+ | malate déshydrogénase | EC 1.1.1.37 | réversible | A ne pas confondre avec l'enzyme malique. |
| Bilan du cycle : acétyl CoA + 3 NAD+ + coenzyme Q + GDP (ou ADP) + Pi + 2 H2O ---> CoASH + 3 NADH + coenzyme QH2 + GTP (ou ATP) + 2 CO2 + 2 H+ |
||||||
|
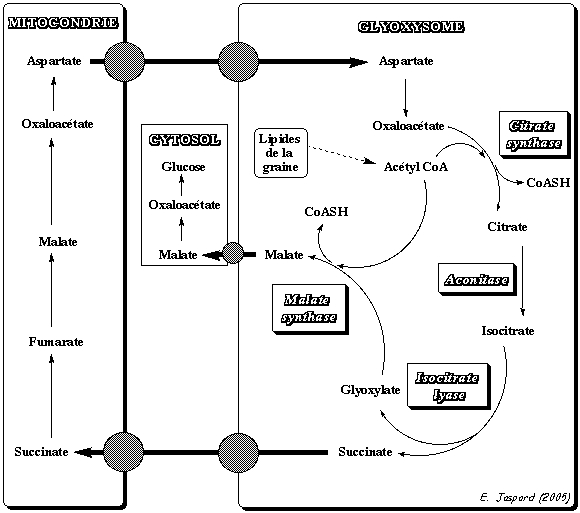
3. Spécificité du cycle de Krebs chez les végétaux : le cycle du glyoxylate Le cycle du glyoxylate n'existe que chez les plantes, les bactéries et les levures. Chez les végétaux oléagineux , les réserves lipidiques de la graine se transforment en glucides qui alimentent l'embryon pendant la germination. La figure ci-dessous illustre ce cycle dans une graine de ricin en germination.
Adapté de : "Principes de Biochimie" Horton et al. (1994)
Or l'oxaloacétate est un précurseur utilisé dans la néoglucogénèse qui aboutit à la reformation de glucose. Le malate peut donc être utilisé comme un précurseur du glucose. |
|
La succinyl-CoA synthétase (aussi appelée succinate thiokinase) catalyse une phosphorylation au niveau du substrat.
|
|
5. Régulation du cycle de Krebs Le premier niveau de régulation est la disponibilité des métabolites points d'entrée du cycle : l'acétyl-CoA et l'oxaloacétate. La concentration de l'acétyl-CoA dépend de l'activité de la pyruvate déshydrogénase et de la régulation de ce complexe multi-enzymatique :
L'oxaloacétate : la disponibilité de ce métabolite dépend de son utilisation par d'autres voies métaboliques comme la néoglucogénèse. |
| Effecteur | Action | Enzyme cible |
| NADH | inhibe | pyruvate déshydrogénase - isocitrate déshydrogénase - α-cétoglutarate déshydrogénase - citrate synthétase |
| acétyl-CoA | inhibe | pyruvate déshydrogénase |
| succinyl-CoA | inhibe | succinyl-CoA synthétase - citrate synthétase |
| ATP | inhibe | citrate synthétase - α-cétoglutarate déshydrogénase |
| citrate | inhibe | phosphofructokinase-1 de la glycolyse |
| calcium | active | pyruvate déshydrogénase - isocitrate déshydrogénase - α-cétoglutarate déshydrogénase |
| Trois réactions du cycle
de Krebs sont irréversibles.
Ces réactions sont catalysées
par des enzymes à régulation
allostérique.
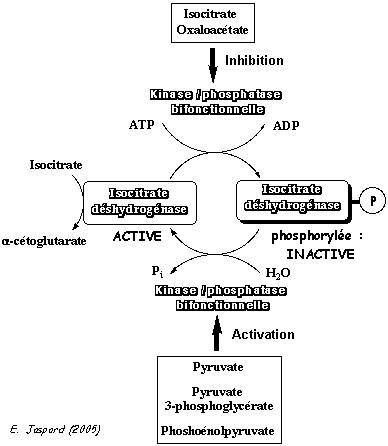
Pour ces 2 raisons (énergétique et catalytique), ces trois réactions constituent des points de contrôle du flux global du cycle de Krebs. Ces enzymes sont : a. La citrate synthase qui catalyse la première réaction du cycle de Krebs. b. L'isocitrate déshydrogénase. Exemple dans le cas de Escherichia coli (figure ci-dessous) : l'enzyme est régulée par [phosphorylation - déphosphorylation] catalysée par une kinase /phosphatase bifonctionnelle.
c. Le complexe de l'α-cétoglutarate déshydrogénase qui catalyse une réaction (figure ci-dessous) analogue à celle du complexe de la pyruvate déshydrogénase.
Source : "The Krebs Cycle" - Garrett & Grisham |
| L'α-cétoglutarate déshydrogénase est un complexe multi-enzymatique composé de 3 enzymes (comme la pyruvate déshydrogénase) | |||
| sous-unité | nom | EC | coenzyme |
| E1 | oxoglutarate déshydrogénase | EC 1.2.4.2 | pyrophosphate de thiamine |
| E2 | dihydrolipoyl succinyltransférase | EC 2.3.1.61 | acide lipoïque |
| E3 | dihydrolipoyl déshydrogénase | EC 1.8.1.4 | FAD |
| Voir un cours sur les mécanismes enzymatiques à plusieurs substrats et plusieurs produits. |
| 6. Liens Internet et références bibliographiques |
| Hans Adolf Krebs (1940) "The citric acid cycle and the Szent-Györgyi cycle in pigeon breast muscle" Biochem. J. 34, 775 - 779 | Article |
|
"Cycle de Krebs" - Equipe multimédia - Université Jussieu - Paris Plantes : "Metacyc pathways" - TCA cycle Plantes : "Metacyc pathways" - glyoxylate cycle KEGG PATHWAY Database |
|
| Fernie et al. (2004) "Respiratory metabolism: glycolysis, the TCA cycle and mitochondrial electron transport" Curr. Opin. Plant Biol. 7, 254 -261 | Article |
![]()