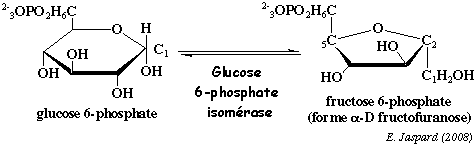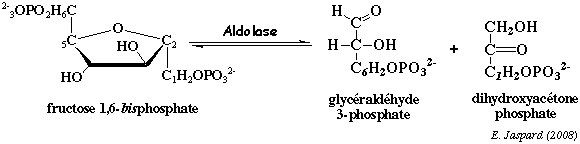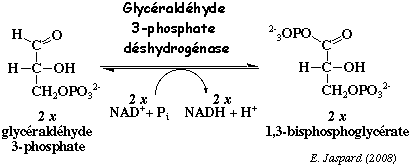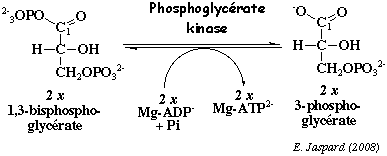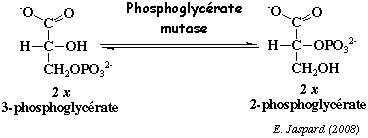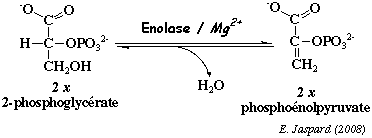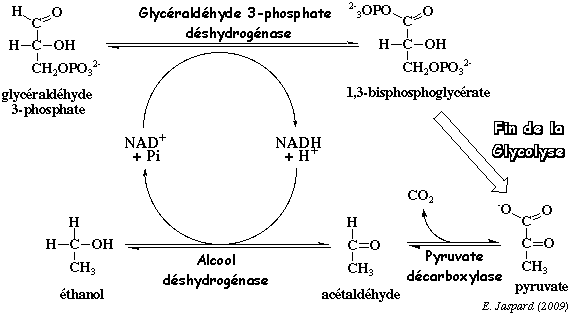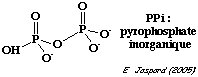| La glycolyse |
| Tweet |
|
|
1. Historique et généralités. 2. Représentation cyclique de Haworth du glucose 3. Les sources de glucose : glycogène, amidon et saccharose 4. Les réactions du tronçon hexoses de la glycolyse
5. Les réactions du tronçon trioses de la glycolyse
|
6. Bilan de la glycolyse 7. Le devenir du pyruvate en anaérobiose
8. Particularité de la glycolyse chez les végétaux
9. Liens Internet et références bibliographiques |
1. Historique et généralités. A la fin du 19è siècle, Eduard Buchner et Martin Hahn ont obtenu un extrait acellulaire des protéines de la levure, capable de convertir le sucre en éthanol et en dioxyde de carbone. Eduard Buchner a publié ses résultats ("Alkoholische Garung ohne Hefezellen" (1897), Berichte der Deutschen Chemischen Gesellschaft) avec ce commentaire (trad. allemand) : "Tout d'abord il a été prouvé que l'initiation du processus de fermentation ne nécessite aucun appareil aussi compliqué que celui que représente une cellule de levure. Le vecteur de l'effet de fermentation du jus pressé est plutôt une substance dissoute, sans doute un corps protéique, qui devrait s'appeler zymase." Eduard Buchner (1860 - 1917) a reçu le Prix Nobel en 1907 pour ses recherches en biochimie et sa découverte de la fermentation sans cellule.
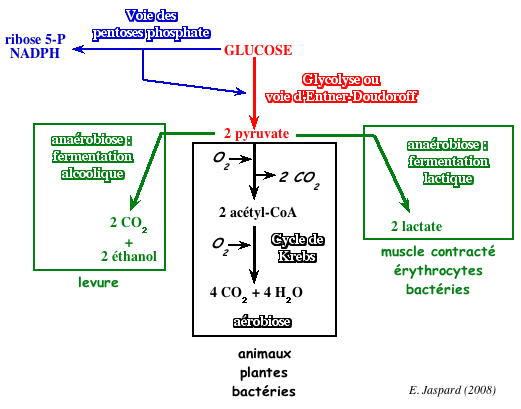
Par ailleurs, Fritz Lipmann (Prix Nobel en 1953) et Herman Kalchar ont établi, en 1941 le rôle énergétique de l'ATP. C'est ainsi que la glycolyse a été la première voie métabolique élucidée et c'est probablement la mieux connue. Cette voie métabolique (du grec "glykys" - sucré et "lysis" - dissolution) est une suite de réactions enzymatiques qui transforment 1 molécule de glucose en 2 molécules de pyruvate. Cette conversion s'accompagne de la production nette de 2 molécules d'ATP. La voie glycolytique se déroule dans le cytoplasme et pratiquement toutes les cellules la possèdent. Pour certaines cellules, c'est même la seule source d'ATP. La glycolyse peut être schématiquement divisée en deux tronçons
Remarques :
|
|
Le glucose est le "fuel" énergétique de la plupart des organismes. Au laboratoire, la quantité de châleur que dégage la combustion complète du glucose solide est de - 673 kcal/mol. Réaction de combustion complète du glucose : C6H12O6 + 6 O2 -> 6 H2O + 6 CO2 Il y a 6 moles de gaz du côté des réactifs et il y a 6 moles de gaz du côté des produits, donc Δn = 0. De ce fait, la variation du volume est nulle, et le produit PdV l'est également. En conséquence, la variation d'énergie interne du système ΔU n'est autre que la châleur dégagée (ΔU = Q), c'est-à-dire - 673 kcal pour chaque mole de glucose. Le signe négatif indique que cette énergie est cédée par le système et elle sert, entre autre, à former de l'ATP qui est le réservoir énergétique de la cellule. Ce rôle du glucose comme source énergétique essentielle est la raison pour laquelle il occupe une position centrale dans le métabolisme. Chez les animaux et les plantes, le glucose a 3 devenirs principaux :
Chez les organismes anaérobies (ou dans le muscle en forte contraction), le pyruvate subit les réactions de la fermentation alcoolique ou de la fermentation lactique. |
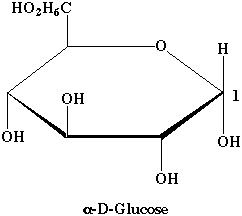
2. Représentation cyclique de Haworth du glucose Voir un cours sur les glucides. Le glucose est un ose à 6 carbones ou hexose. Il existe 8 stéréoisomères de la série D et 8 stéréoisomères de la série L. La figure ci-dessous représente le D - glucose car le carbone 6 est au dessus du cycle.
Il existe deux isomères liés au carbone 1 qui sont sont appelés anomères α et β. La figure ci-contre représente l'α-D-glucose car l'hydroxyle porté par le carbone 1 est en dessous du cycle. Si l'hydroxyle est au dessus du cycle, il s'agit du β-D-glucose. Le glucose comporte 6 sommets et un tel cycle porte le nom de pyranose. Il s'agit donc de l'α-D-glucopyranose. |
|
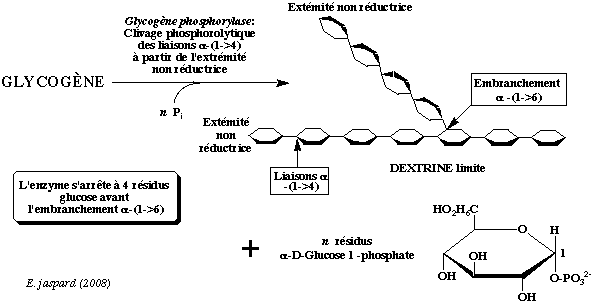
a. Le glycogène C'est un polysaccharide car il est composé de plus de 20 unités d'oses. Le glycogène est constitué :
Les résidus glucose du glycogène sont clivés par la glycogène phosphorylase à partir de l'extrémité non réductrice du glycogène. Voir un cours sur la régulation de la dégradation du glycogène. |
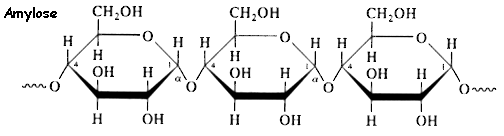
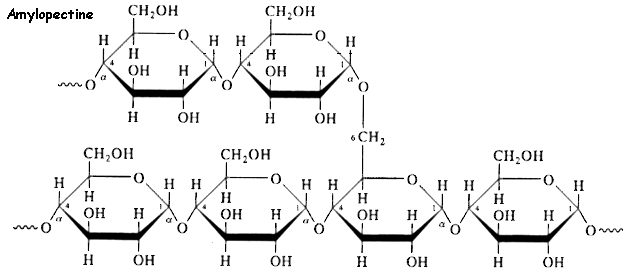
b. L'amidon C'est un polysaccharide des cellules végétales. Il se présente sous la forme d'un mélange de deux polysaccharides : l'amylose et l'amylopectine. L'amylose représente 15 à 30% de la masse de l'amidon. C'est un polymère linéaire de 600 à 1000 résidus glucose liés par une liaison α-(1,4)-D-glucosidique.
Cette longue chaîne prend la forme d'une hélice (6 résidus de glucose par tour d'hélice), stabilisée par des liaisons hydrogène entre les groupements hydroxyle et les molécules d'eau. L'amylopectine représente 70 à 85% de la masse de l'amidon. Elle diffère de l'amylose du fait qu'il s'agit d'un polymère ramifié (jusqu'à 100 millions de Dalton) :
Chez l'homme, il existe 2 formes d'α-amylase (endoglycosidase) :
L'α-amylase a besoin de calcium pour fonctionner. La β-amylase (EC 3.2.1.2) est une exoglycosidase qui catalyse la libération progressive de résidu maltose à partir des extrémités non réductrices de l'amidon. Remarque : le sigle "β" n'a rien à voir avec la configuration du carbone anomère des oses. En effet, l'α-amylase et la β-amylase n'hydrolysent que les liaisons α-(1,4)-D-glycosidiques. Le maltose est un diholoside α-D-glucopyranosyl-(1,4)-β-D-glucopyranose. C'est un sucre réducteur puisque l'hydroxyle du carbone anomère du second glucose est libre. |
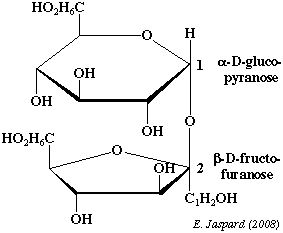
c. Le saccharose ou α-D-glucopyranosyl-β-D-fructofuranoside ou sucrose On peut considérer le saccharose comme étant :
C'est l'une des principales sources alimentaires d'α-D-glucose car il est extrémement représenté dans le règne végétal et tout particulièrement dans la canne à sucre et la betterave. C'est le seul diholoside non reducteur (suffixe oside) trouvé à l'état naturel avec le tréhalose. En effet, l'hydroxyle du carbone anomère du fructose (carbone 2) est engagé dans une liaison osidique avec le carbone anomère du glucose (carbone 1). Il est hydrolysé par la saccharase en α-D-glucopyranose et en β-D-fructofuranose. Les solutions de saccharose présentent un pouvoir rotatoire mais pas le phénomène de mutarotation. |
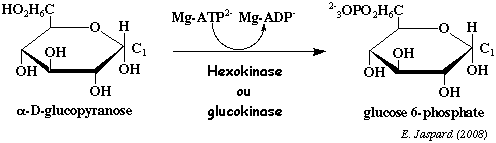
| 4. Les réactions du tronçon hexoses de la glycolyse a. La réaction catalysée par l'hexokinase ou la glucokinase
On donne le nom de kinase aux enzymes qui catalysent une réaction de phosphorylation. Il existe 3 isoformes d'hexokinase (E.C. 2.7.1.1). C'est une enzyme de 102.000 Da chez les organismes multicellulaires et environ 50.000 Da chez les bactéries. L'hexokinase catalyse la phosphorylation de certains autres hexoses : le D-fructose et le D-mannose. La glucokinase est l'isoforme IV de l'hexokinase que l'on trouve dans le foie. C'est un monomère de 465 acides aminés d'environ 50.000 Da. Cette réaction consomme 1 molécule d'ATP par molécule de glucose. C'est donc une réaction trés exergonique, donc irréversible :
|
b. La réaction catalysée par la glucose 6-phosphate isomérase (EC 5.3.1.9) La glucose 6-phosphate isomérase est une enzyme d'environ 63.000 Da chez l'homme. Remarque : ne pas la confondre avec la "glucose isomérase" qui désigne D-xylose isomérase(EC 5.3.1.5). Cette réaction d'isomérisation transforme un aldohexose en cétohexose.
Les structures cycliques pyranose et furanose sont les plus stables en solution. La glucose 6-phosphate isomérase fixe de préférence la forme cyclique du glucose 6-phosphate : elle est alors ouverte au sein du site actif de l'enzyme puis convertie en cétose via un intermédiaire ènediolate. Cette réaction requiert le magnésium. Dans les conditions standard : ΔG°' = + 0,4 kcal.mol-1. En revanche, dans la cellule, c'est une réaction qui se déroule au voisinage de l'équilibre, soit ΔG' # 0 kcal.mol-1. Dans la cellule on trouve : 70% de glucose 6-phosphate et 30% de fructose 6-phosphate. |
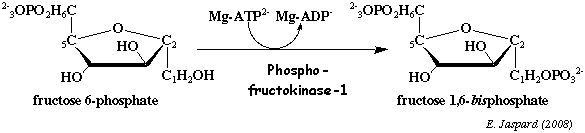
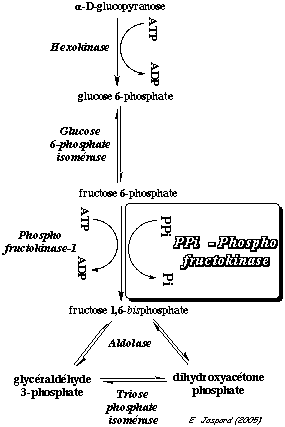
c. La réaction catalysée par la phosphofructokinase-1 (E.C. 2.7.1.11) Cette enzyme phosphoryle une seconde fois le fructose 6-phosphate pour former le fructose 1,6-bisphosphate.
La phosphofructokinase-1 (PFK-1) est l'enzyme clé de la régulation du flux global de la glycolyse (avec la pyruvate kinase). C'est aussi l'une des enzymes clé du métabolisme énergétique de manière générale, de concert avec un certain nombre d'autres enzymes, notamment la protéine kinase activée par l'AMP ("5'-AMP-activated ") ou AMPK. Cette réaction consomme 1 molécule d'ATP par molécule de glucose. C'est donc une réaction trés exergonique, donc irréversible : ΔG°' = - 3,4 kcal.mol-1 et ΔG' = - 5,3 kcal.mol-1. La PFK-1 de Escherichia coli est un homotétramère, c'est-à-dire 4 sous-unités identiques de 34.800 Da (320 acides aminés par sous-unité). Voir un cours sur le rôle clé de la PFK-1 dans la régulation de la glycolyse. |
d. La réaction catalysée par l'aldolase (EC 4.1.2.13) L'aldolase scinde le fructose 1,6-bisphosphate en 2 trioses :
Dans les conditions standard : ΔG°' = + 5,7 kcal.mol-1 donc la réaction est impossible dans le sens de la coupure du fructose 1,6-bisphosphate. En revanche dans la cellule on trouve : [fructose 1,6 bisphosphate] >> 2 [trioses]. En conséquence, du fait des concentrations physiologiques (cellulaires) des métabolites impliqués dans cette récation, elle se déroule au voisinage de l'équilibre dans la cellule (ΔG' # 0 kcal.mol-1). Les aldolases des mammifères et des végétaux supérieurs contiennent 2 acides aminés essentiels pour le mécanisme : une lysine et une cystéine. La lysine forme une base de Schiff (voir le mécanisme catalytique). Les aldolases des micro-organismes suivent un autre mécanisme qui met en jeu un cation métalique.
|
| "Morale"du tronçon hexoses
Il y a un investissement énergétique initial de 2 molécules d'ATP par molécule de glucose. Ceci peut sembler paradoxal pour une voie métabolique qui génère de l'énergie sous forme d'ATP. Mais cet investissement énergétique est rentable. En effet, les deux carbones aux extrémités de la chaîne du glucose (C1 et C6) sont phosphorylés, ainsi, les 2 trioses le sont également. Remarque : en regard du rôle central de la glycolyse dans le métabolisme général, on peut se demander quelle "stratégie la vie aurait mise en place" si 1 seul carbone était phosphorylé ou si les carbones [1 - 2] ou [1 - 3] ou [4 - 6] ou [5 - 6] l'étaient. |
| 5. Les réactions du tronçon trioses a. La réaction catalysée par la triose phosphate isomérase (EC 5.3.1.1)
La suite de la glycolyse implique le glycéraldéhyde-3-phosphate : à la fois celui formé directement et celui formé à partir de la dihydroxyacétone-phosphate selon la réaction décrite ci-contre, catalysée par la triose phosphate isomérase. Cependant, dans la cellule la proportion de triose phosphate sous forme de dihydroxyacétone-phosphate est de 95%. Celà souligne le rôle capital de la triose-phosphate isomérase qui doit fournir en glycéraldéhyde-3-phosphate la voie de la glycolyse. En effet, cette enzyme est l'une des rares dont la vitesse de catalyse est si élevée qu'elle n'a pour limite que la vitesse de diffusion des substrats (voir le cours d'enzymologie qui a trait aux paramètres cinétiques). Dans les conditions standard : ΔG°' = + 1,8 kcal.mol-1. C'est une réaction qui se déroule au voisinage de l'équilibre dans la cellule, soit ΔG' # 0 kcal.mol-1. La triose-phosphate isomérase est un dimère ou un tétramère. Chez l'homme, la sous-unité a une masse molaire 26.700 Da (249 acides aminés). La triose-phosphate isomérase de Giardia lamblia est l'un des rares exemples de dimérisation de cette enzyme via un pont disulfure. |
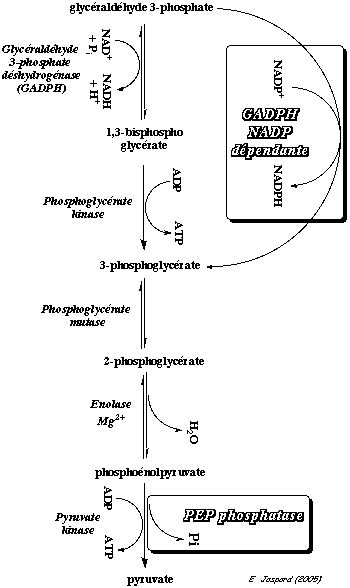
b. La réaction catalysée par la glycéraldéhyde 3-phosphate déshydrogénase (EC 1.2.1.12) Cette enzyme catalyse la seule réaction d'oxydo-réduction de la glycolyse.
La réaction se déroule en 5 étapes faisant intervenir, entre autre, le groupe sulfhydryle d'une cystéine, groupe ionisé par une histidine qui joue le rôle de base générale. Dans les conditions standard : ΔG°' = + 1,5 kcal.mol-1. C'est une réaction qui se déroule au voisinage de l'équilibre dans la cellule, soit ΔG' # 0 kcal.mol-1. Cependant, elle génère une liaison à haut potentiel énergétique (liaison anhydride ou acyl phosphate au niveau du C1 / ΔG°'hydrolyse = - 11,8 kcal.mol-1) contenue dans le 1,3 -bisphosphoglycérate. Cette énergie sera utilisée dans la réaction suivante pour la synthèse d'une molécule d'ATP (liaison phosphoanhydride). La glycéraldéhyde 3-phosphate déshydrogénase est un dimère, un homotétramère ou un hétérotétramère. Chez l'homme, la sous-unité a une masse molaire de 36.000 Da (335 acides aminés). Il existe au moins 4 isoenzymes : phosphorylante à NAD+ / phosphorylante à NADP+ / non phosphorylante à NAD+ / non phosphorylante à NADP+ Cette enzyme est impliquée dans d'autres processus cellulaires : l'activation de la transcription, l'initiation de l'apoptose et la migration de vésicules du réticulum endoplasmique vers l'appareil de Golgi. |
c. La réaction catalysée par la phosphoglycérate kinase (EC 2.7.2.3) Cette enzyme catalyse le transfert du groupement phosphate à haut potentiel énergétique (acyl phosphate sur le C1 du 1,3 -bisphosphoglycérate) sur l'ADP pour former le 3-phosphoglycérate et l'ATP.
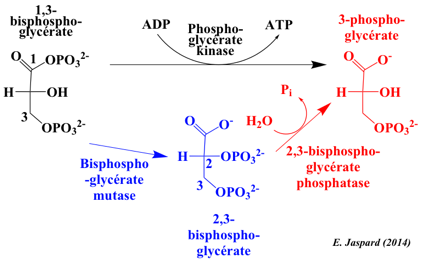
Ce mode de synthèse d'ATP s'appelle phosphorylation au niveau du substrat. L'autre mode de synthèse de l'ATP est la phosphorylation oxydative : cycle de Krebs / réoxydation des co-enzymes réduits par la chaîne respiratoire / force proton motrice et ATP synthase. Dans les conditions standard : ΔG°' = - 4,5 kcal.mol-1. Bien qu'une molécule d'ATP soit synthétisée, c'est une réaction qui se déroule au voisinage de l'équilibre dans la cellule, soit ΔG' # 0 kcal.mol-1. Elle ne constitue donc pas un point de contrôle de la glycolyse à l'inverse des 3 autres réactions de cette voie catalysées par des kinases. La phosphoglycérate kinase est un monomère ou un dimère. Chez l'homme, la sous-unité a une masse molaire 44.600 Da (417 acides aminés). Un intermédiaire capital pour l'oxygénation de l'hémoglobine : le 2,3-bisphosphoglycérate (2,3-BPG) Il est formé à partir du 1,3-bisphosphoglycérate par la bisphosphoglycérate mutase (EC 5.4.2.4) , une enzyme spécifique des érythrocytes et des cellules du placenta.
Les pH faibles inhibent la bisphosphoglycérate mutase et diminuent la concentration du 2,3-BPG. Le 2,3-BPG n'a pas de rôle dans la glycolyse et il est transformé en 3-phosphoglycérate par la 2,3-bisphosphoglycérate phosphatase (EC 3.1.3.13 - voie Rapoport-Luebering). En revanche, dans les érythrocytes et les cellules du placenta, le 2,3-BPG joue un rôle majeur dans le relarguage de l'oxygène fixée sur l'hémoglobine en stabilisant la forme désoxy-hémoglobine (forme T). Il y a un équilibre très fin entre la nécessité de générer de l'ATP (besoins énergétiques de la cellule) et la nécessité de maintenir le rapport optimal [désoxy-hémoglobine / oxy-hémoglobine]. Les plantes ne forment pas le 2,3 -BPG. |
d. La réaction catalysée par la phosphoglycérate mutase (EC 5.4.2.11) Cette enzyme catalyse l'isomérisation du 3-phosphoglycérate en 2-phosphoglycérate.
Dans le cas des isoenzymes de phosphoglycérate mutase d'animaux et de levures, le mécanisme catalytique passe par un intermédiaire où une histidine est phosphorylée qui amène à un intermédiaire 2,3 -bisphosphoglycérate. Chez les isoenzymes de phosphoglycérate mutase de plantes, il n'y a pas formation de l'intermédiaire 2,3 -bisphosphoglycérate. Dans les conditions standard : ΔG°' = + 1,1 kcal.mol-1. C'est une réaction qui se déroule au voisinage de l'équilibre dans la cellule, soit ΔG' # 0 kcal.mol-1. Chez les mammifères, il existe 2 isoenzymes : la phosphoglycérate mutase de type M dans le muscle et de type B dans les autres tissus. La phosphoglycérate mutase est un monomère, un dimère ou un tétramère. La sous-unité a une masse molaire de 32.000 Da (254 acides aminés). |
e. La réaction catalysée par l'énolase (EC 4.2.1.11) Cette enzyme catalyse la formation du phosphoénolpyruvate à partir du 2-phosphoglycérate par une réaction de trans-élimination d'une molécule d'eau.
C'est une métaloenzyme et le magnésium est indispensable à son acticité catalytique : le Mg2+ stabilise la forme dimèrique de l'enzyme et participe à la fixation du substrat. Les ions fluorures (F-) inhibent en se complexant aux ions Mg2+ au site actif. Dans les conditions standard : ΔG°' = + 0,4 kcal.mol-1. C'est une réaction qui se déroule au voisinage de l'équilibre dans la cellule, soit ΔG' # 0 kcal.mol-1. Cependant, le phosphoénolpyruvate formé contient une liaison énol phosphate. Le potentiel énergétique de cette liaison est parmi les plus élevés : ΔG°'hydrolyse = - 14,8 kcal.mol-1. Il existe 3 types de sous-unités (α, β, γ / 434 acides aminés) qui peuvent se combiner pour former 5 isoenzymes différentes de l'énolase, selon leur localisation tissulaire. L'énolase active (sous la forme d'un homo- ou d'un hétérodimère) a une masse molaire de 82.000 à 100.000 Da selon l'isoenzyme. L'énolase est impliquée dans d'autres processus : par exemple elle joue un rôle de récepteur et d'activateur du plasminogène à la surface des leukocytes et des neurones. Elle participe également à la stimulation de la production des immunoglobulines. |
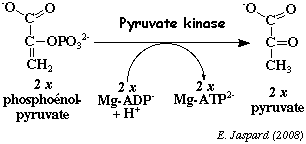
f. La réaction catalysée par la pyruvate kinase (E.C. 2.7.1.40) Cette enzyme catalyse la transformation du phosphoénolpyruvate en pyruvate avec synthèse d'ATP par phosphorylation au niveau du substrat.
La pyruvate kinase est une enzyme à régulation allostérique comme la phosphofructokinase 1. Cette réaction est l'un des points de contrôle de la glycolyse. Cette réaction forme 1 molécule d'ATP par molécule de glucose. C'est donc une réaction trés exergonique, donc irréversible : ΔG°' = - 7,5 kcal.mol-1 et ΔG' = - 4,0 kcal.mol-1. Chez les mammifères, il existe 4 isoenzymes : type L - foie / type R - hématies / type M1 - muscle, cerveau et coeur / type M2 - tissus du foetus. La pyruvate kinase est un homotétramère d'une masse de 235.000 Da (531 acides aminés par sous-unité). Le pyruvate a deux devenirs :
|
"Morale"du tronçon trioses Les 2 trioses-phosphate issus du tronçon hexoses étant phosphorylés, chaque molécule de glycéraldéhyde-3-phosphate va permettre la formation de 2 molécules d'ATP. Le bilan énergétique de la glycolyse est donc une synthèse nette de 2 molécules d'ATP par molécule de glucose. Celà peut sembler dérisoire en regard de la synthèse d'ATP après la réoxydation des nucléotides réduits par la chaîne respiratoire. Mais pour les organismes anaérobies c'est absolument essentiel. Il n'y a qu'une réaction d'oxydo-réduction dans la glycolyse catalysée par la glycéraldéhyde 3-phosphate déshydrogénase. |
|
glucose (C6H12O6) + 2 ADP + 2 Pi + 2 NAD+ ---> 2 pyruvate + 2 ATP + 2 NADH + 2 H+ + 2 H2O
La formation d'ATP catalysée par la phosphoglycérate kinase et la pyruvate kinase est effectuée par transfert du groupe phosphoryle d'un composé à haut potentiel énergétique sur l'ADP (phosphorylation au niveau du substrat). On obtient respectivement :
|
| Enzyme | E.C. | M. M. | A. A. | [E] | Sact | CoF | ΔG°' | ΔG' | substrat(s) | produit(s) | [Met] |
| hexokinase | 2.7.1.1 | 50 ou 102 |
α-D-glucopyranose + ATP | glucose 6-phosphate + ADP |
G6P : 3.900 ATP : 8.000 |
||||||
| glucokinase | 50 | 465 | monomère | - 4,0 | - 8,0 | F6P : 1.500
ADP : 600 |
|||||
| glucose 6-phosphate isomérase | 5.3.1.9 | 63 | Mg2+ | + 0,4 | # 0 | glucose 6-phosphate | fructose 6-phosphate | F 1,6-bisP : 80 | |||
| phosphofructokinase-1 | 2.7.1.11 | 34,8 | 320 | homotétramère | - 3,4 | - 5,3 | fructose 6-phosphate + ATP | fructose 1,6-bisphosphate + ADP | G3P : 80 | ||
| aldolase | 4.1.2.13 | 160 | 364 | 810 | tétramère | lysine / base de Schiff ou Zn2+ |
+ 5,7 | # 0 | fructose 1,6-bisphosphate | glycéraldéhyde 3-phosphate + dihydroxyacétone phosphate | DHAP : 160 |
| triose phosphate isomérase | 5.3.1.1 | 26,7 | 249 | 220 | di- / tétramère | + 1,8 | # 0 | dihydroxyacétone phosphate | glycéraldéhyde 3-phosphate | 1,3 - BPG : 50 |
|
| glycéraldéhyde 3-phosphate déshydrogénase | 1.2.1.12 | 36 | 335 | 1,400 | di- / tétramère | NAD+ | + 1,5 | # 0 | glycéraldéhyde 3-phosphate + NAD+ | 1,3 -bisphosphoglycérate + NADH | NAD+ : 540 NADH : 50 |
| phosphoglycérate kinase | 2.7.2.3 | 44,6 | 417 | 130 | mono- / dimère | - 4,5 | # 0 | 1,3 -bisphosphoglycérate + ADP | 3-phosphoglycérate + ATP | 3 - PG : 200 |
|
| phosphoglycérate mutase | 5.4.2.1 | 32 | 254 | 240 | mono- / di- / tétramère | + 1,1 | # 0 | 3-phosphoglycérate | 2-phosphoglycérate | 2 -PG : 20 | |
| énolase | 4.2.1.11 | 82 à 100 | 434 | 540 | homo- / hétérodimère | Mg2+ | + 0,4 | # 0 | 2-phosphoglycérate | phosphoénolpyruvate | PEP : 64 |
| pyruvate kinase | 2.7.1.40 | 235 | 531 | homotétramère | Mg2+ / K+ | - 7,5 | - 4,0 | phosphoénolpyruvate | pyruvate | pyruvate : 380 | |
Les liens hypertexte renvoient vers des données contenues dans la base de données "Expasy - Uniprot". On y trouve une trés grande quantité d'informations sur la ou les fonction(s) des enzymes, leur structure, leur régulation, leur localisation cellulaire et les processus biologiques dans lesquels elles sont impliquées.
|
|||||||||||
7. Le devenir du pyruvate en anaérobiose Les organismes qui effectuent la glycolyse en conditions anaérobies ont développé des systèmes simples pour réoxyder le NADH formé dans le cytosol. Ainsi, la glycolyse se maintient indéfiniment. a. La fermentation alcoolique par les levures Au cours de la fermentation alcoolique, le pyruvate est d'abord décarboxylé en acétaldéhyde et CO2 par la pyruvate décarboxylase (EC 4.1.1.1).
La pyruvate décarboxylase utilise deux cofacteurs : la thiamine pyrophosphate et l'ion magnésium (deux de chaque sont requis pour la réaction). C'est un homodimère ou un homotétramère (60.000 Da et 563 acides aminés par sous-unité). Les levures sont parmi les rares organismes à possèder la pyruvate décarboxylase. C'est ainsi que les brasseurs font la bière. L'acétaldéhyde est ensuite réduit en éthanol par l'alcool déshydrogénase (EC 1.1.1.1). Au cours de cette réaction, le NADH est réoxydé en NAD+ qui retourne dans la glycolyse au niveau de la réaction catalysée par la glycéraldéhyde éshydrogénase. L'alcool déshydrogénase utilise l'ion zinc comme cofacteur et elle est présente chez quasiment tous les organismes :
|
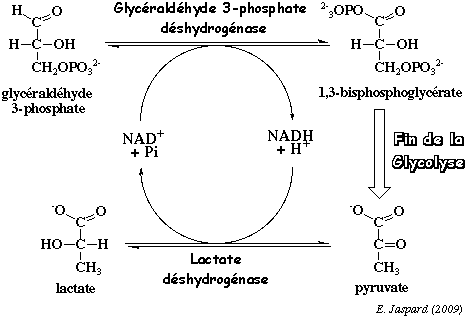
b. La voie homofermentaire ou voie homolactique La plupart des organismes sont dépourvus de pyruvate décarboxylase et donc incapables de transformer le pyruvate en éthanol. Ces organismes réduisent donc le pyruvate en lactate par la lactate déshydrogénase (EC 1.1.1.27).
Au cours de cette réaction, le NADH est réoxydé en NAD+ qui retourne dans la glycolyse au niveau de la réaction catalysée par la glycéraldéhyde 3-phosphate déshydrogénase. Comme il n'existe d'autre issue pour le lactate qu'un retour au pyruvate, on considère le lactate comme une impasse métabolique. L'acide lactique formé abaisse le pH. On utilise cette propriété avec les bactéries lactiques : en effet, la caséïne du lait (et d'autres protéines) sont alors dénaturées et s'agrègent. Il y a formation de yaourth. |
|
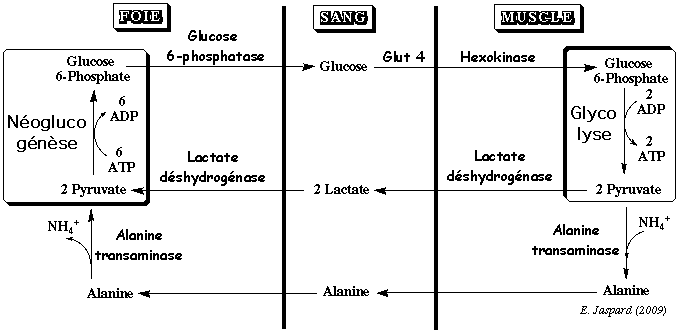
Chez l'homme, il existe 5 isoenzymes de lactate déshydrogénase. Celle du muscle est un homotétramère (35.000 Da et 332 acides aminés par sous-unité). Dans le muscle squelettique, la concentration de la lactate déshydrogénase est 300 µM, celle du lactate 3.700 µM et celle du pyruvate 380 µM. Quand le muscle squelettique se contracte, le pyruvate est transformé en lactate par une lactate déshydrogénase. Le lactate rejoint le sang puis le foie où il est re-transformé en pyruvate par une lactate déshydrogénase.
Le foie métabolise le pyruvate :
Le foie est le principal pourvoyeur de glucose aux muscles, notamment pour la contraction musculaire. Cette synergie entre organes où le foie reçoit du pyruvate des muscles et renvoie du glucose aux muscles s'appelle le cycle des Cori. Ce cycle ne peut durer indéfiniment car il y a une consommation nette de 4 molécules d'ATP. Le cycle des Cori vient du nom du couple de biochimistes qui ont contribué à son élucidation, Gerty et Carl Cori (Prix Nobel en 1947 pour leur description du métabolisme du glycogène). |
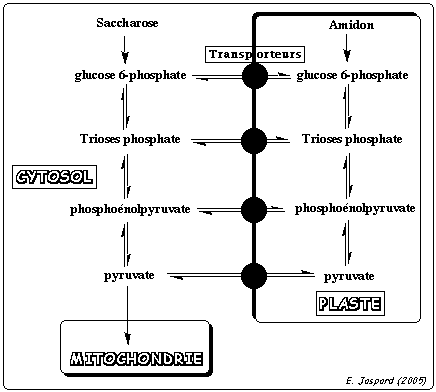
8. Particularité de la glycolyse chez les végétaux La glycolyse chez les végétaux présente des particularités qui assurent une plus grande flexibilité du métabolisme (adaptation à l'environnement). La glycolyse a lieu dans 2 compartiments cellulaires : le cytosol et les plastes.
Les métabolites passent d'un compartiment à l'autre via des transporteurs membranaires. |
|
Tronçon hexoses : la pyrophosphate inorganique (PPi) - phosphofructokinase catalyse la conversion du fructose 6-phosphate en fructose 1,6-bisphosphate. Elle duplique donc l'action de la phosphofructokinase-1.
Tronçon trioses :
Voir un cours sur la respiration chez les végétaux. Voir un cours sur la photosynthèse. |
| 9. Liens Internet et références bibliographiques |
| Vision globale du métabolisme (Expasy) - (Figure interactive) | Aller au site |
| "Biocarta" : site interactif sur la signalisation cellulaire | Aller au site |
| "The UCSD-Nature Signaling Gateway" - A comprehensive resource for information about cell signaling. | Aller au site |
| KEGG PATHWAY Database | Aller au site |
![]()