| Régulation de la glycolyse |
| Tweet |
|
|
1. Préambule 2. L'hexokinase et la glucokinase 3. La phosphofructokinase-1 (PFK-1) 4. La pyruvate kinase |
5. La phosphofructokinase-2 6. La charge énergétique adénylique 7. Rôle de l'insuline 8. Liens Internet et références bibliographiques |
|
1. Préambule La glycolyse produit de l'énergie rapidement mais sur de courtes durées. La régulation de la glycolyse est liée aux besoins énergétiques de la cellule. Les sites de régulation de la glycolyse correspondent aux 3 étapes irréversibles de cette voie métabolique. Ce sont les réactions catalysées par des enzymes à régulation allostérique. La modulation de l'activité de ces enzymes est un moyen de contrôler le flux global de la glycolyse. Enfin, pour bien comprendre la régulation de la glycolyse, il faut aussi prendre en compte le rôle clé d'une enzyme qui ne fait pas partie de cette voie métabolique mais qui a une incidence majeure sur le métabolisme énergétique en général : la protéine kinase activée par l'AMP ("5'-AMP-activated protein kinase") ou AMPK. |
| Résumé des activateurs (A) et des inhibiteurs (I) des 3 enzymes clé de la régulation de la glycolyse | |||
| hexokinase* | phosphofructokinase | pyruvate kinase | |
| glucose 6-phosphate | I | ||
| fructose 1,6-bisphosphate | A | ||
| fructose 2,6-bisphosphate | A | A (pour certains organismes tels que Kinetoplastida - Trypanosoma) | |
| ATP (substrat et effecteur) | I (concentrations > 10-3 M : effet hétérotrope) | I | |
| ADP | A | A (substrat - effet homotrope) | |
| AMP | A | I | |
| acétyl CoA | I | ||
| NADH | A puis I | ||
| citrate | I | I | |
| PEP | I | A (substrat - effet homotrope) | |
| Pi | A | ||
|
*La glucokinase n'est pas inhibée par le glucose 6-phosphate. |
|||
|
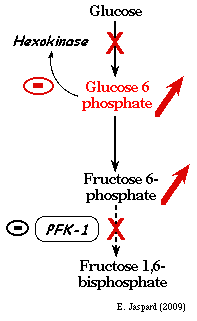
2. L'hexokinase et la glucokinase L'hexokinase (E.C. 2.7.1.1) est inhibée par le produit de la réaction qu'elle catalyse : le glucose-6-phosphate. Cette inhibition se fait de deux manières :
La régulation de l'hexokinase n'est pas un point de contrôle majeur du flux de la glycolyse car une grande partie du glucose-6-phosphate provient de l'hydrolyse du glycogène (glycogène --> glucose 1-phosphate) puis isomérisation en glucose 6-phosphate par la phosphoglucomutase. En conséquence la réaction catalysée par l'hexokinase peut être contournée. En revanche, la régulation de l'activité de l'hexokinase est importante dans le cadre de la réversion de la glycolyse et de l'activation de la néoglucogénèse quand la concentration en ATP est élevée. |
|
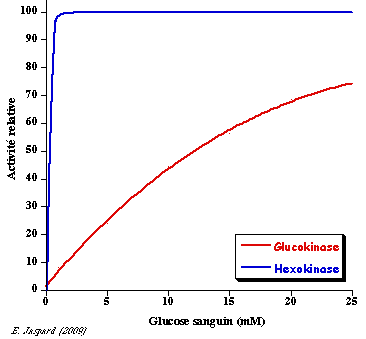
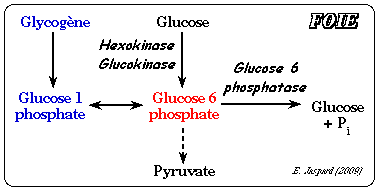
La glucokinase (E.C. 2.7.1.2) est l'isoforme IV de l'hexokinase (mammifères) que l'on trouve dans le foie. Elle a un KM élevé pour le glucose et n'est donc active qu'à fortes concentrations de glucose (figure ci-dessous) :
KMglucokinase = 10 mM - KMhexokinase = 0.1 mM - [glucose sanguin] = 5 mM Le KM élevé de la glucokinase pour le glucose permet que le foie stocke celui-ci sous forme de glycogène quand le taux de glucose sanguin est élevé. A l'inverse la glucose-6-phosphatase du foie (enzyme de la néoglucogénèse - E.C. 3.1.3.9) catalyse l'hydrolyse du glucose-6-phosphate et le glucose est relargué dans le sang ce qui maintient sa concentration circulante. En conséquence, la glucokinase et la glucose-6-phosphatase, que l'on ne trouve quasi exclusivement que dans le foie, permettent à celui-ci de contrôler le taux de glucose sanguin.
|
|
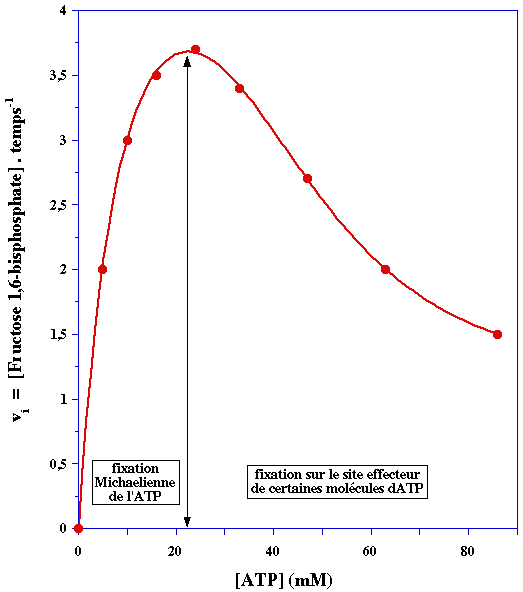
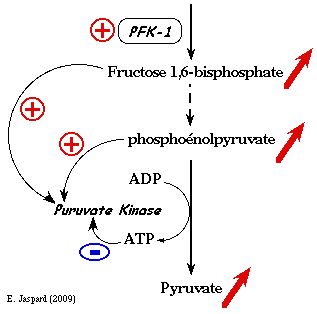
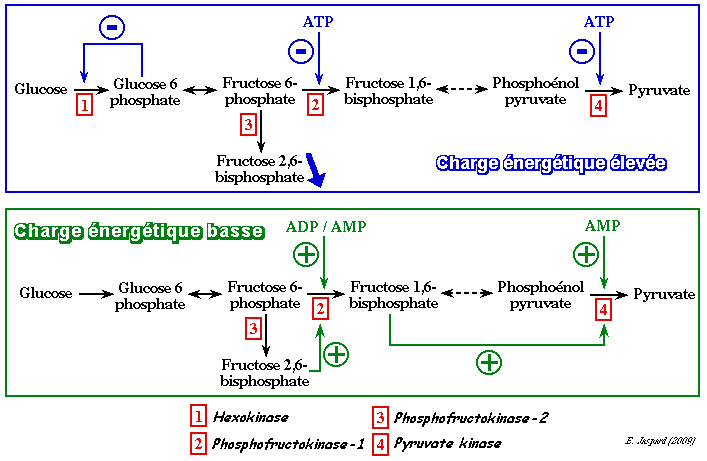
3. La phosphofructokinase-1 (PFK-1) Elle joue un rôle primordial dans la régulation du flux de la glycolyse. En effet, l'activité de cette enzyme est régulée par la concentration de ces substrats (l'ATP et le fructose 6-phosphate) mais aussi par celles de nombreux effecteurs liés à la production d'énergie par la phosphorylation oxydative. Ces effecteurs sont : l'ATP lui-même (voir ci-après) / l'ADP / l'AMP / le phosphoénolpyruvate / le phosphate inorganique / le citrate / le NADH L'ATP est un cas particulier car c'est l'un des deux substrats de la PFK-1 mais c'est aussi un effecteur, puisque la PFK-1 possède :
1. Partie gauche de la courbe de saturation ci-dessous
Elle traduit l'effet de l'ATP sur la vitesse de la réaction enzymatique en tant que substrat. L'allure hyperbolique de cette première partie de la courbe indique que la fixation de l'ATP aux 4 sites catalytiques (la PFK-1 est un homotétramère) s'effectue selon un mécanisme "Michaelien" (voir le cours). Celà signifie qu'il n'y a pas d'effet coopératif dans la fixation des molécules d'ATP. 2. Partie droite de la courbe de saturation A partir d'une certaine concentration, des molécules d'ATP se fixent aussi sur un site qui n'est pas le site catalytique : ce site régule l'activité catalytique. C'est un site effecteur :
3. En conséquence : une partie des molécules d'enzyme étant inactive, la concentration réelle de complexe enzyme - substrat ([ES]) est moindre que la concentration d'enzyme. La vitesse de catalyse diminue puisque : vi = kcat x [ES]. |
|
Les différentes
phosphofructokinases : nomenclatures et réactions catalysées - Les liens renvoient vers la base de données
"Expasy" EC 2.7.1. : phosphotransferases avec un groupement alcool comme accepteur (kinases et phosphotransferases) |
||
| EC | Nomenclature | Réaction catalysée |
| 2.7.1.11 |
|
ATP + D-fructose 6-phosphate ---> ADP + D-fructose 1,6-bisphosphate |
| 2.7.1.56 |
|
ATP + D-fructose 1-phosphate ---> ADP + D-fructose 1,6-bisphosphate |
| 2.7.1.90 |
|
diphosphate + D-fructose 6-phosphate ---> phosphate + D-fructose 1,6-bisphosphate |
| 2.7.1.105 |
|
ATP + D-fructose 6-phosphate <==> ADP + D-fructose 2,6-bisphosphate |
| 2.7.1.146 |
|
ADP + D-fructose 6-phosphate ---> AMP + D-fructose 1,6-bisphosphate |
4. La pyruvate kinase (E.C. 2.7.1.40) Elle est activée par le fructose 1,6-bisphosphate. Or ce métabolite se situe en amont de la réaction catalysée par la pyruvate kinase : on appelle ce phénomène "feed - forward". En revanche la pyruvate kinase est inhibée par l'ATP.
La réaction catalysée par la pyruvate kinase est controlée dans le foie en partie par la modulation de la quantité d'enzyme synthétisée. C'est le cas d'un très grand nombre d'enzymes qui contrôlent le flux d'une voie métabolique. |
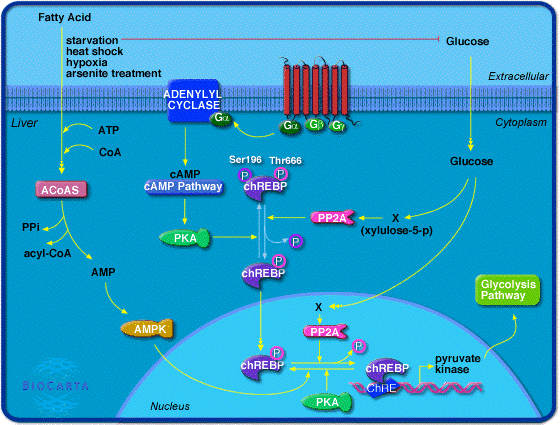
La transcription du gène codant la pyruvate kinase est sous le contrôle d'un facteur de transcription : "Carbohydrate Responsive Element Binding Protein" - ChREBP.
Source : Biocarta
De fortes concentrations de glucose activent la protéine phosphatase 2A xylulose-5-phosphate-dépendante (PP2A) qui déphosphoryle Ser 196 ce qui entraîne la délocalisation ChREBP dans le noyau. Dans le noyau, PP2A déphosphoryle Thr 666 de ChREBP, ce qui entraîne l'activation de la transcription du gène codant la pyruvate kinase. L'excès de glucose est alors converti en pyruvate puis en acétyl-CoA, le principal précurseur de la synthèse des acides gras, qui est la forme de stockage de l'énergie à long terme. |
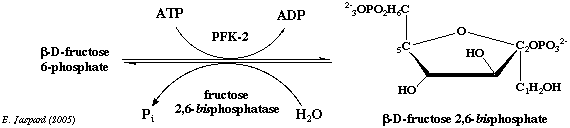
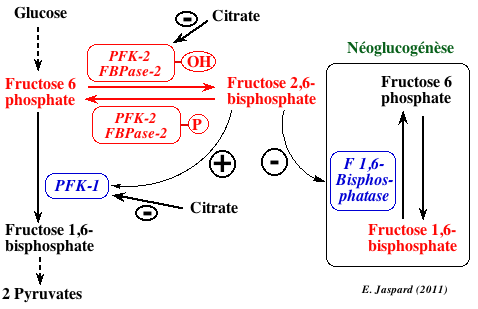
5. La phosphofructokinase-2 (E.C. 2.7.1.105) Le fructose 2,6-bisphosphate (F2,6BP) est formé à partir du fructose 6-phosphate par la phosphofructokinase-2 ou PFK-2. La PFK-2 est un homodimère d'environ 100.000 daltons.
Le F2,6BP est un puissant activateur de la PFK-1 (sauf chez certains protistes pour lesquels il est activateur de la pyruvate kinase). Dans le foie, le F2,6BP est un inhibiteur de la fructose-1,6-bisphosphatase, une enzyme de la néoglucogénèse. Le citrate est un inhibiteur de la PFK-2 : il y a moins de F2,6BP synthétisé. Or le F2,6BP étant un activateur de la PFK-1, celà amplifie l'inhibition de la PFK-1 par le citrate.
Dans les cellules humaines, le citrate cytosolique est un précurseur capital pour la synthèse des acides gras, des triacyl-glycérols, du cholestérol et des lipoprotéines de faible densité ("low-density lipoprotein"). De plus, le citrate cytosolique régule la balance énergétique de la cellule en activant la synthèse des acides gras et en ralentissant la glycolyse et la β-oxidation des acides gras. La vitesse de synthèse des acides gras dans le foie et les cellules adipeuses (les 2 principaux tissus pour cette synthèse) est directement corrélée à la concentration du citrate dans le cytosol. Cette concentration dépend partiellement de l'import du citrate au travers de la membrane plasmique via le transporteur Na+-citrate. |
|
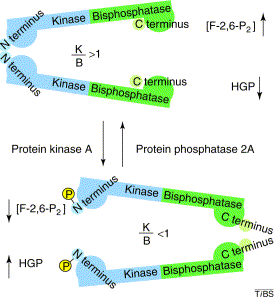
Régulation de la PFK-2 par le glucagon Dans le foie, la PFK-2 est sous le contrôle du glucagon, une hormone produite par le pancréas quand le taux de glucose sanguin s'abaisse. Quand la concentration du glucagon augmente, la protéine kinase AMP cyclique dépendante (PKA) phosphoryle une sérine ou une thréonine de la [PFK-2 / FBPase-2] (exemples : foie de rat : S32 - coeur de boeuf : S84 - levure : T157).
Source : Okar et al. (2001) - K/B : rapport des activités [PFK-2/FBPase-2] - HGP : production du glucose par le foie La déphosphorylation est catalysée par la protéine phosphatase 2A xylulose-5-phosphate-dépendante (PP2A) qui est activée par le glucose (voir ci-dessus).
La phosphorylation :
Conséquences :
|
|
6. La charge énergétique adénylique (Atkinson, DE, 1968) La charge énergétique adénylique (CEA) de la cellule est définie par la relation :
[ATP] + 1/2 [ADP] Cette relation exprime la fraction molaire en ATP (qui contient 2 liaisons phosphoanhydride à haut potentiel énergétique) plus la moitié de la fraction molaire en ADP (qui n'en contient qu'une). La CEA est donc un senseur énergétique de la cellule qui lui permet de mesurer l'énergie dont elle dispose à un instant donné. Il s'agit d'un paramètre de valeur universelle atteignant des valeurs comparables chez tous les organismes vivants. La CEA est très sensible aux variations de l'environnement interne des organismes ou de l'environnement extérieur : plus un organisme subit l'effet d'un stress, plus il consomme d'ATP pour contre-balancer ces effets, plus la CEA de cet organisme baisse. En théorie, la valeur de la CEA peut varier de 0 (il n'y a que de l'AMP) à 1 (il n'y a que de l'ATP). Cependant, dans la plupart des cellules, la CEA est trés finement régulée et on observe des variations de 0,7 (forte hydrolyse de l'ATP et de l'ADP en AMP) à 0,95 (typiquement une cellule au repos).
|
|
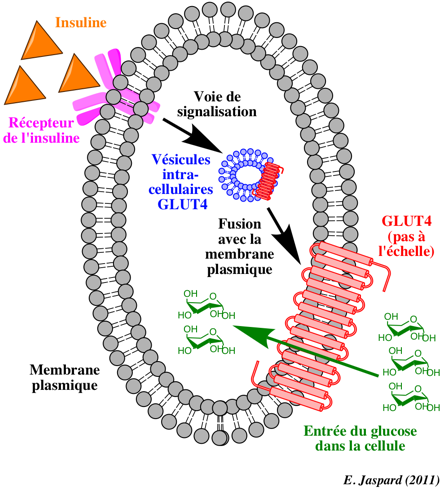
La mobilisation du glucose par les muscles squelettiques représente environ 70% du glucose prélevé au sérum. Ce processus est donc extrêmement important dans l'homéostasie du glucose. La contraction et l'insuline sont les stimuli majeurs qui activent le transport du glucose dans les muscles squelettiques. Dans une cellule non stimulée ou quand la concentration en insuline est faible, le transporteur insulino-dépendant du glucose GLUT4 ("glucose transporter 4" - gène : SLC2A4) est localisé dans des vésicules de stockage des cellules hépatiques et musculaires.
Quand le niveau de glucose circulant est élevé, l'insuline est libérée par les ilots de Langerhans et elle augmente la mobilisation du glucose via une augmentation :
L'absorption du glucose augmente : le glucose pénètre davantage dans le muscle et le tissu adipeux par l'intermédiaire de ce type de transporteur. |
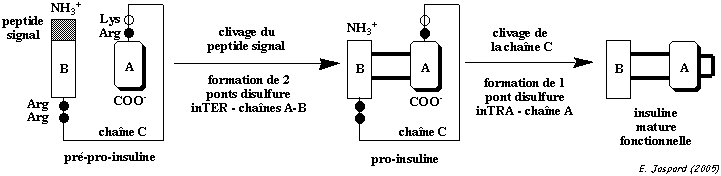
Maturation de l'insuline L'insuline est synthétisée dans les cellules β des îlots de Langerhans sous la forme d'un précurseur : la pré-pro-insuline.
L'insuline adopte sa forme fonctionnelle. Cette maturation par l'élimination du peptide C retarde l'apparition de l'activité hormonale de l'insuline jusqu'à ce qu'elle soit empaquetée dans les granules de sécrétion.
L'effet net est une augmentation du stock de glycogène (foie) et une diminution de la production de glucose (muscle, tissu adipeux, foie). Une carence en insuline (diabète insulino dépendant) a donc pour conséquence une élévation de la concentration du glucose sanguin. |
|
Visualisation de l'insuline de porc à une résolution de 1,5 Å Code PDB : 4INS Les 4 chaînes (2 fois A et 2 fois B - dimère de dimère) sont colorées.
|
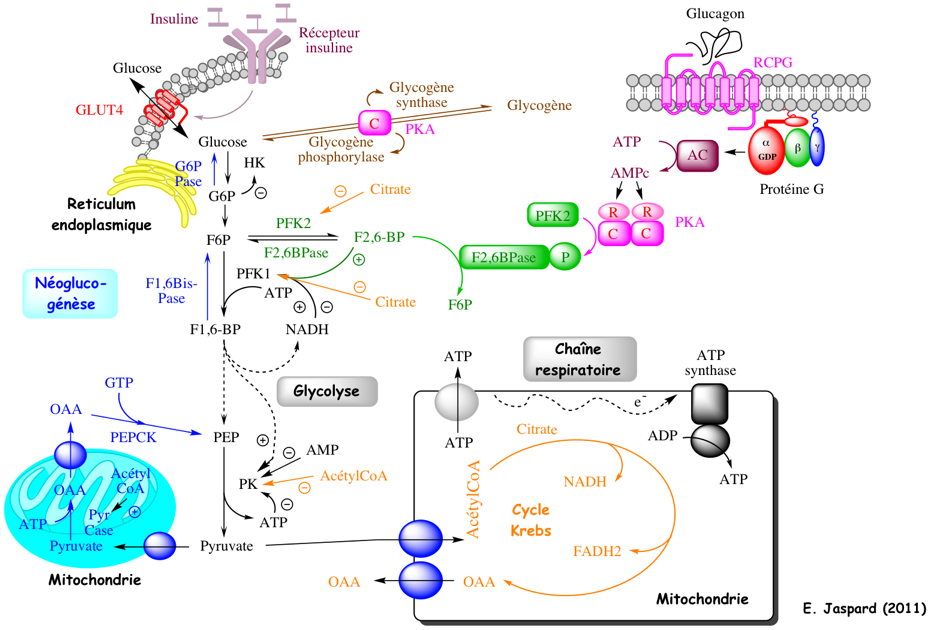
Schéma général simplifié de l'homéostasie du glucose et du métabolisme énergétique N'est pas représenté le rôle de la protéine kinase activée par l'AMP ("5'-AMP-activated protein kinase") ou AMPK.
|
| Abréviations de la figure | ||
|
RCPG : récepteur couplé à une protéine G OAA : oxaloacétate AC : adénylate cyclase |
PFK2 : phosphofructokinase 2 F2,6BPase : fructose-2,6-bisphosphatase F2,6-BP : fructose-2,6-bisphosphate |
|
| 8. Liens Internet et références bibliographiques |
|
Vision globale du métabolisme (Expasy) - Figure interactive KEGG PATHWAY Database |
![]()