| Régulation du métabolisme énergétique : la protéine kinase activée par l'AMP (AMPK) |
| Tweet |
|
|
1. Introduction : rôle majeur de l'AMPK 2. Structure de l'AMPK 3. Double rôle de l'AMP dans l'activation de l'AMPK 4. Activation de l'AMPK |
5. Rôle de l'AMPK dans l'activation de la glycolyse 6. Rôle de l'AMPK dans le transport du glucose 7. Liens Internet et références bibliographiques |
|
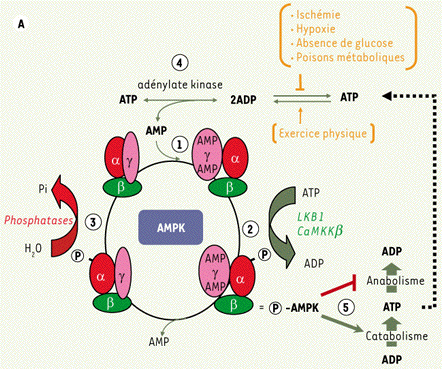
1. Introduction : rôle majeur de l'AMPK Chez les mammifères, les cellules doivent maintenir une balance énergétique stable, caractérisée par un rapport des concentrations [ATP/ADP] égal à 10/1. Tous les organismes ont donc développé des mécanismes pour adapter leur métabolisme aux contraintes externes variables et en particulier aux modifications énergétiques qui résultent :
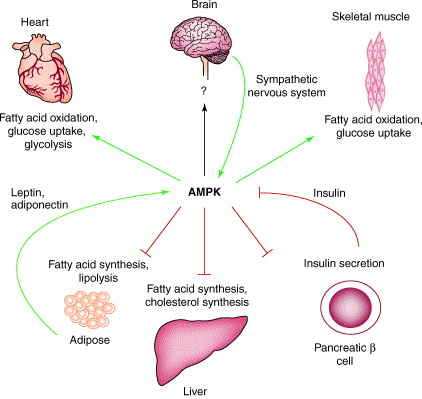
Afin de garder constante l'homéostasie énergétique, l'hypothalamus contrôle la prise alimentaire et la dépense énergétique selon les informations reçues des tissus périphériques (par exemple, via la leptine, protéine hormonale de 167 acides aminés). L'homéostasie est la capacité d'un organisme à maintenir son équilibre physiologique interne malgré les contraintes extérieures. Parmi les divers moyens dont dispose l'organisme pour palier à ces modifications énergétiques, la protéine kinase activée par l'AMP ("5'-AMP-activated protein kinase" ou AMPK) joue un rôle tout à fait majeur. En effet, bien qu'elle agisse au niveau cellulaire, l'AMPK joue aussi un rôle dans la régulation de la prise alimentaire et de la dépense énergétique de l'organisme entier.
Source : Carling D. (2004) Remarque : ne pas confondre l'AMPK avec la protéine kinase dépendante de l'AMP cyclique ("Cyclic-AMP dependent protein kinase"). |
L'AMPK est une enzyme ubiquitaire :
Elle est considérée comme un senseur métabolique car, dans sa forme activée, l'AMPK :
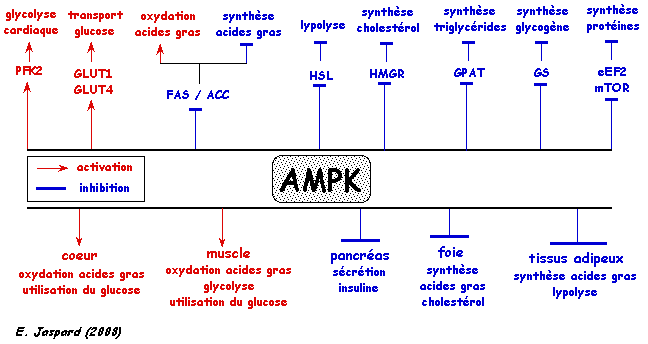
Schéma ci-dessous : principales protéines cibles et cascades de signalisation induites par l'activation de l'AMPK.
PFK2 : phosphofructokinase 2; FAS : fatty acid synthase; HSL : hormone-sensitive lipase; GPAT : glycerol-3-phosphate acyltransferase; MCD : malonyl-CoA decarboxylase; GS : glycogen synthase and creatine kinase; eEF2 : translation elongation factor 2; mTOR : mammalian target La régulation du métabolisme cellulaire par l'AMPK dépend de la phosphorylation de ses protéines cibles. Le motif consensus du site de phosphorylation est φX(B,X)XX[Ser/Thr]XXXφ avec P0 = site de phosphorylation [Ser/Thr]; des résidus hydrophobes (φ) en positions P+4 et P-5 ; un résidu basique (B, typiquement Arg) en position P-1 à P-4; X est n'importe quel acide aminé. |
|
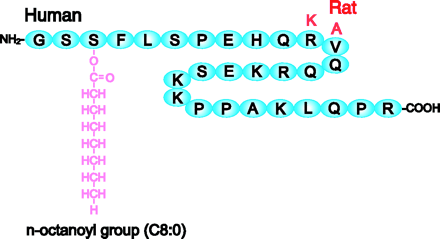
L'AMPK est donc une enzyme clé de la régulation du métabolisme énergétique en réponse notamment : (i) à des signaux hormonaux (exemple : la ghréline, une hormone qui stimule l'appétit); (ii) à des signaux métaboliques; (iii) aux effets d'adipokines (leptine, adiponectine). Figure ci-dessous : séquences en acides aminés de la ghréline de l'homme et du rat (les 2 acides aminés en rouge).
Source : Kojima & Kangawa (2004) La ghréline est un peptide de 28 acides aminés, dont la Ser 3 est modifiée par un acide gras (acide n-octanoique). Cette modification est essentielle pour l' activité de la ghréline. L'AMPK joue également un rôle dans la prise alimentaire : (i) régulation de la synthèse d'insuline; (ii) modulation de certaines fonctions de l'hypothalamus; (iii) régulation de la sensation de satiété. Elle a donc des impacts sur l'obésité et diverses formes de diabètes. Enfin, l'activation de l'AMPK exerce des effets à long terme sur la transcription des gènes et la synthèse des protéines. En regard de l'importance de ses fonctions biologiques, c'est une cible pharmacologique à visée métabolique (insulinorésistance, obésité, diabète) et cardiologique (ischémie cardiaque). |
|
L'AMPK est constituée de 3 sous-unités (hétérotrimère) :
La réaction catalysée par la sous-unité α (EC 2.7.11.1) peut s'écrire : ATP + une protéine -> ADP + une phosphoprotéine. Exemples :
|
| sous-unité / isoforme | acides aminés (Homo sapiens) | N° d'accession PDB | N° d'accession UniProt |
| α1 | 550 | ----- | Q13131 |
| α2 | 552 | 2YZA | P54646 |
| β1 | 270 | 1Z0N | Q9Y478 |
| β2 | 272 | 2F15 | O43741 |
| γ1 | 331 | 2UV4 | P54619 |
| γ2 | 569 | 2V8Q | Q9UGJ0 |
| γ3 | 489 | ----- | Q9UGI9 |
La sous-unité α
Source : Carling D. (2004) La sous-unité β
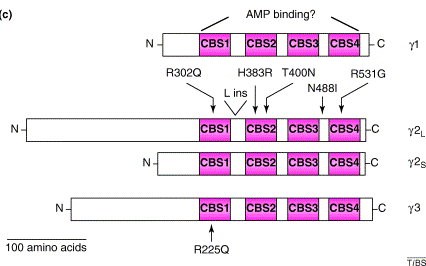
La sous-unité γ
Source : Carling D. (2004) Les structures secondaires [β-α-β-β-α] d'un domaine CBS se replient en une structure tertiaire globulaire qui forme un feuillet β anti-parallèle à trois brins avec deux hélices α d'un côté. Les domaines CBS sont toujours par paire dans la structure des protéines qui les contiennent. Cette paire de domaines forme un pseudo-dimère via leurs feuillets β et la structure ainsi obtenue s'appelle domaine Bateman. La sous-unité γ fixe l'AMP et l'ATP :
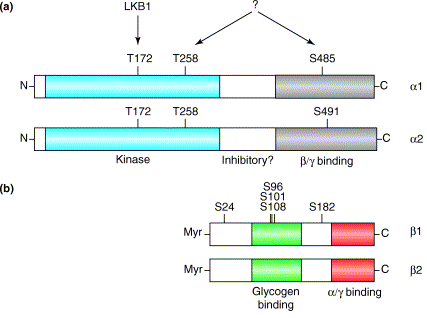
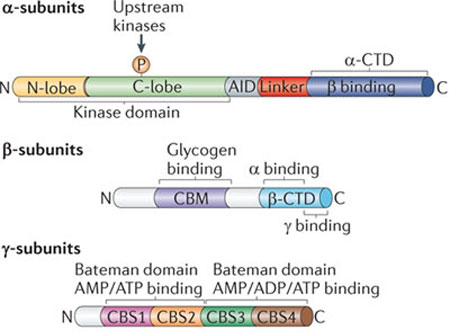
Représentation globale des différents domaines et de leurs fonctions des 3 sous-unités de l'AMPK
Source : Hardie et al. (2012) AID ("Auto-Inhibitory Domain") : domaine auto-inhibiteur; CBM ("Carbohydrate-Binding Module") : domaine de fixation des sucres; N-lobe : lobe N-terminal du domaine kinase. |
| 3. Double rôle de l'AMP dans l'activation de l'AMPK a. Fixation coopérative de l'AMP Quand le muscle se contracte, l'ATP est hydrolysé en ADP : ATP + H2O -> ADP + Pi. L'ADP peut reformer de l'ATP en donnant un phosphate inorganique à une autre molécule d'ADP, ce qui aboutit à la formation d'ATP et d'AMP : 2 ADP -> ATP + AMP. Cette réaction est catalysée par l'adénylate kinase (EC 2.7.4.3). Plus l'AMP est formé au cours de la contraction musculaire, plus le rapport des concentrations [AMP/ATP] augmente.
L'élévation du rapport des concentrations [AMP/ATP] augmente l'activité de l'AMPK : plus précisément, l'AMP active l'AMPK d'un facteur 2 à 10. |
| activateur | inhibiteur |
| AMP | ATP |
récepteurs couplés à la phospholipase Cγ |
phosphocréatine |
| hormones sécrétées par les tissus adipeux (les adipokines) telles que la leptine et l'adiponectine | ------ |
b. Induction de la phosphorylation de la sous-unité catalytique La fixation de l'AMP induit des changements de conformation de la sous-unité catalytique α (effet allostérique). Dés lors, T172 du site catalytique de la sous-unité α est phosphorylée par l'AMPK kinase LKB1 (voir ci-dessous). Cette phosphorylation est indispensable à l'activité de l'AMPK. L'AMP augmente la phosphorylation de l'AMPK d'un facteur 50 à 100 et inhibe sa déphosphorylation. |
|
Il a fallu beaucoup de temps avant d'identifier la kinase responsable de la phosphorylation de T172 de l'AMPK. Cette kinase est souvent appelée AMPK kinase ou AMPKK dans la littérature. C'est un complexe de plusieurs protéines : "Pseudokinase STE-related adaptor protein" (STRAD) ; "mouse protein 25" (MO25); CaMKKβ : [Ca2+/calmodulin-activated protein kinase]-kinase-β; LKB1 (ou "STK11") : sérine/thréonine kinase (EC 2.7.11.1) qui phosphoryle l'AMPK sur Thr 172.
Source : Foretz, M. (2006) Chez l'homme, LKB1 est composée de 433 acides aminés et le domaine protéine - kinase (domaine catalytique) se situe entre les acides aminés 44 et 309.
Elle est cependant exprimée dans moins de tissus que ne l'est LKB1 qui est ubiquitaire. Le relarguage de Ca2+ intracellulaire engendre une demande en ATP (régulation du métabolisme du glycogène). Ainsi, l'activation de l'AMPK en réponse aux flux calciques confère à la cellule un moyen pour anticiper une augmentation des besoins en ATP. |
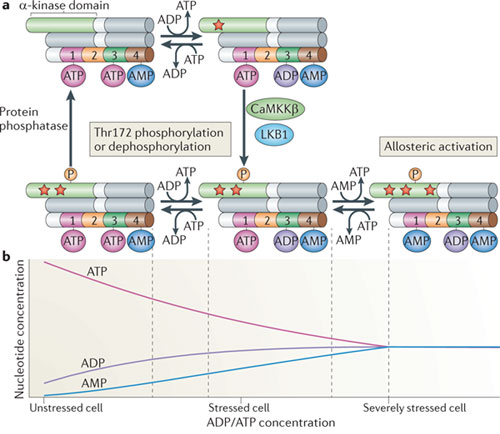
Figure a - ci-dessous : modèle résumant les différents états des 3 sous-unités de l'AMPK.
Source : Hardie et al. (2012) L'AMPK est activée par des augmentations de concentration d'AMP et d'ADP.
Le graphique de la figure b ci-dessus montre les changements de concentrations de l'ATP, de l'ADP et de l'AMP qui accompagnent le passage d'une cellule complètement chargée au repos (à gauche) dans un état où cette cellule subit un sévère stress énergétique (à droite), qui correspond à une augmentation d'un facteur dix du rapport [ADP/ATP]. Ce graphique a été généré en supposant que la réaction catalysée par l'adénylate kinase (voir ci-dessous) est à l'équilibre et que la concentration totale des nucléotides adényliques demeure constante. Dans la cellule complètement chargée, la concentration d'AMP est très faible. Mais la variation de concentration d'AMP (quand le rapport [ADP/ATP] augmente) est toujours beaucoup plus élevée que celles de l'ATP ou de l'ADP. |
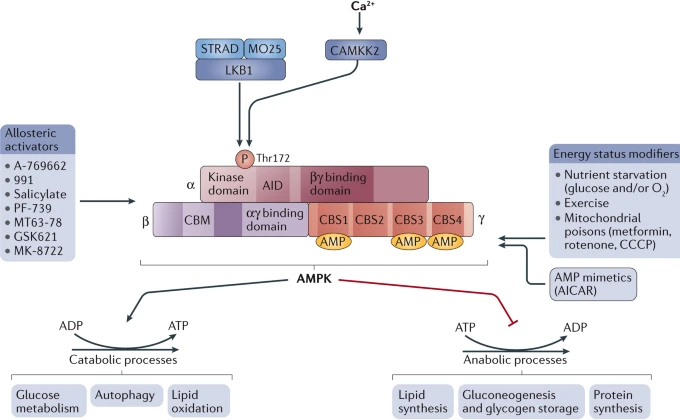
Les activateurs de l'AMPK Plusieurs facteurs conduisent à l'activation de l'AMPK (figure ci-dessous) :
Source : Herzig & Shaw (2018) |
| Activateurs directs de l'AMPK | Activateurs indirects de l'AMPK |
| 5-aminoimidazole-4-carboxamide riboside (AICAR) dérivés de la thienopyridone (A-769662) et du benzimidazole (composé 911) (laboratoire Abbott) salicylate (précurseur de l'asprine = acétyl salicylate) acide 5-(5-hydroxyl-isoxazol-3-yl)-furan-2-phosphonique (composé 2) et son précurseur : composé-13 acide 2-chloro-5-[[5-[[5-(4,5-dimethyl-2-nitrophenyl)-2-furanyl]methylene]-4,5-dihydro-4-oxo-2-thiazolyl]amino]benzoique (appelé PT1) MT 63–78 (Debio0930 - Mercury & CreaGen) |
biguanides (exemple : metformine) |
| Voir les tableaux 1 et 2 de l'article Steinberg & Carling (2019). | |
|
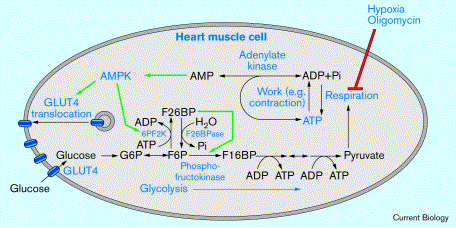
5. Rôle de l'AMPK dans l'activation de la glycolyse L'inhibition de la respiration par le phénomène d'hypoxie / anoxie (diminution du taux d'oxygène dans le sang) ou par l'oligomycine (inhibiteur de la phosphorylation par l'ATP synthase) abolit la synthèse d'ATP à partir de l'ADP et du Pi (figure ci-dessous). Plus la contraction du muscle a lieu, plus le rapport des concentrations [ADP/ATP] augmente puisque l'ATP est hydrolysé : ATP + H2O -> ADP + Pi Or une augmentation du rapport des concentrations [ADP/ATP] entraîne une augmentation beaucoup plus grande du rapport des concentrations [AMP/ATP] du fait de la réaction catalysée par l'adénylate kinase (EC 2.7.4.3) : 2 ADP -> ATP + AMP.
Source : Hardie D. G. (2000) Le nom des enzymes est en bleu; les flèches noires indiquent les interconversions métaboliques; les flèches vertes indiquent les processus d'activation des réactions et la flèche rouge les processus d'inhibition. C'est l'une des explications de l'effet Pasteur (1861) observé en absence d'oxygène : la synththèse d'ATP chute => le rapport des concentrations [AMP/ATP] augmente => l'AMP active l'AMPK => l'AMPK active la phosphofructokinase-2 (6PF2K) en la phosphorylant sur la Ser 466 => la phosphofructokinase-2 synthétise le fructose-2,6-bisphosphate (F26BP) => le F26BP active la phosphofructokinase 1 : la glycolyse anaérobie (fermentation) est activée. |
|
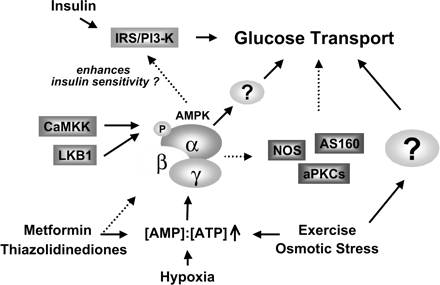
6. Rôle de l'AMPK dans le transport du glucose La mobilisation du glucose par les muscles squelettiques représente environ 70% du glucose prélevé au sérum. Ce processus est donc extrêmement important dans l'homéostasie du glucose. La contraction et l'insuline sont les stimuli majeurs qui activent le transport du glucose dans les muscles squelettiques.
Bien que les effets soient semblables, les mécanismes du transport du glucose médiés par l'AMPK indiquent que les signaux sont distincts de ceux de l'insuline.
Source : Fujii et al. (2006) La metformine (composé contre le diabète de la famille des biguanides) et les thiazolidinediones (utilisés dans le traitement de la forme 2 du diabète) activent l'AMPK. Cependant, les mécanismes exacts du transport du glucose médiés par l'AMPK sont encore assez mal connus. Voir un diagramme général [format PDF] qui récapitule les voies dans lesquelles l'AMPK est impliquée. |
| 7. Liens Internet et références bibliographiques |
|
Pasteur L. (1861) "Expériences et vues nouvelles sur la nature des fermentations" Comp. Rend. Acad. Sci. 5 , 1260 - 1264 Hardie et al. (1989) "The AMP-activated protein kinase-a multisubstrate regulator of lipid metabolism" Trends Biochem. Sci. 14, 20 - 23 Foretz M. (2006) "Régulation du métabolisme énergétique par l'AMPK : une nouvelle voie thérapeutique pour le traitement des maladies métaboliques et cardiaques" Médecine/Science 22, 381 - 388 |
|
|
Bateman A. (1997) "The structure of a domain common to archaebacteria and the homocystinuria disease protein" Trends Biochem. Sci. 22, 12 - 13 Hardie D. G. (2000) "Metabolic control: A new solution to an old problem" Curr. Biology 10, R757 - R759 Carling D. (2004) "The AMP-activated protein kinase cascade - a unifying system for energy control" Trends Biochem. Sci. 29, 18 - 24 |
|
|
Kojima & Kangawa (2004) "Ghrelin: Structure and Function" Physiol. Rev. 85, 495 - 522 Fujii et al. (2006) "AMP-activated protein kinase and the regulation of glucose transport" Am. J. Physiol. Endocrinol. Metab. 291, E867 - E877 Hardie et al. (2012) "AMPK: a nutrient and energy sensor that maintains energy homeostasis" Nat. Rev. Mol. Cell Biol. 13, 251 - 262 Herzig & Shaw (2018) "AMPK: guardian of metabolism and mitochondrial homeostasis" Nat. Rev. Mol. Cell Biol. 19, 121 - 135 Steinberg & Carling (2019) "AMP-activated protein kinase: the current landscape for drug development" Nat. Rev. Drug Discov. 18, 527 - 551 |
|
![]()