| Le complexe calcium/calmoduline et l'activation des protéines |
| Tweet |
|
|
1. L'ion calcium et la calmoduline 2. Séquences nucléotidiques et polypeptidiques (primaires) de la calmoduline 3. Structures de la calmoduline
|
4. Mécanismes de changement de conformation de la calmoduline
5. Protéines fixant le calcium
6. Mécanisme d'activation des pompes Ca2+-ATPases par la calmoduline 7. Liens Internet et références bibliographiques |
|
1. L'ion calcium et la calmoduline L'ion calcium (Ca2+) est le métal le plus abondant dans le corps humain (2 % du poids total). Le calcium :
Le rayon ionique du calcium est plus important que celui d'autres ions métalliques trouvés dans les organismes (Na+, K+, Mg2+). Son enveloppe électronique lui permet d'établir 6 à 8 liaisons de coordination avec les protéines avec un bon degré de fléxibilité. Il se fixe donc fréquemment dans des "poches" constituées d'acides aminés acides ou polaires (Asp et Glu, Asn), ce qui ne contraint pas ou peu la conformation du squelette carboné. Dans une cellule au repos, la concentration du calcium libre et d'environ 50 à 100 nM. Un stimulus externe peut induire une augmentation de la concentration à 10 µM. L'ion calcium est un messager intracellulaire trés important.
La calmoduline a été découverte par Wai Yiu Cheung en 1970, puis par Shiro Kakiuchi et ses collègues plus tard la même année :
Cette découverte a contribué à la compréhension du mécanisme d'action du calcium :
|
2. Séquences nucléotidiques et polypeptidiques de la calmoduline L'alignement de séquences de la CaM révèle des résultats surprenants. a. Séquences nucléotidiques Séquence consensus codant la CaM issue de 44 séquences de CaM de plantes (E. Jaspard - non publié) ATGGC.GAtc agCTcAccGA cGA.CAGAtc tc.GAgTTcA AgGAgGCcTT caGCcT.TTc GAcAAgGA.G GcGAtGGtTG cATcAC.ACc AAGGAgcTtG GaACtGTgAT GcG.TCacTg GG.CAgAAcC CaACtGAgGC tGAgCT.CAg GAcATGATcA AtGAaGTtGA TGCtGATGG. AAtGGaACcA TtGAcTTcCC .GAGTTccT. AAccTgATGG C.cG.AAgAT GAAgGAcACt GAtTCtGAGG AgGAgCTcAA gGAgGCtTTC ag.GTgTTtG AcAAgGAtCA gAAtGG.TTc ATcTCtGCtG CtGAgcT.CG cCAtGT.ATG AC.AAcCT.G G.GAgAAgCT .AC.ga.ga. ..aGAgGTtG AtGAGATGAT ccg.GAgGCt GAtgT.GAtG GtGATGG.CA gaTcAAcTAt GAgGAgTTtG TcAAggTcAT GATGGCcAAG TGA 453
Il existe plusieurs gènes codant la CaM (exemple : 8 gènes chez la pomme de terre) :
|
| b.
Séquences polypeptidiques
La CaM est une protéine constituée d'une chaîne polypeptidique de 148 - 149 acides aminés (de masse molaire d'environ 16.700 Dalton). Séquence primaire consensus de la CaM issue de 39 séquences de CaM de plantes (E. Jaspard - non publié) A..L...Q.. ..KEAF.LFD KDGDGCIT.. E..TV.RSL. QNPTE.ELQD MI.E.D.D.N GTI.F.E.L. LMA.K.K.TD .EE.LKEAF. VFDKDQ.G.I SA...RHVM. NLGEKL... EV..MI.EAD .DGDGQ.N.. EFV..M... 149
|
| Organisme | similarité avec la CaM humaine (%) | identitité avec la CaM humaine (%) |
| Oryza sativa (riz) | 96,6 | 90,5 |
| Tetrahymena pyriformis | 96,6 | 91,9 |
| Dictyostelium discoideum | 93,2 | 89,2 |
| Neurospora crassa | 91,9 | 81,1 |
| Electophorus electricus | 100 % | 99,4 |
|
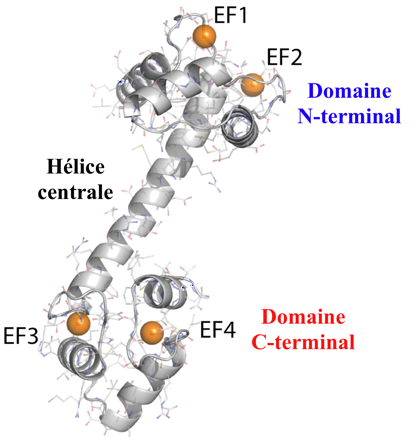
3. Structures de la calmoduline a. Caratéristiques générales de la structure tridimensionnelle La CaM est l'une des protéines dont on connait le mieux la structure : près de 200 structures ont été obtenues par différentes méthodes. On distingue 3 domaines dans la CaM : le domaine N-terminal / une hélice centrale de 7 tours / le domaine C-terminal.
Source : K. Hines - Aldrich lab Le domaine N-terminal et le domaine C-terminal possèdent chacun 2 sites de fixation du calcium :
|
|
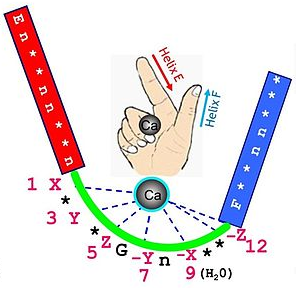
Ce motif est composé de 30 acides aminés et contient 2 hélices α (E et F - figurées respectivement par l'index et le pouce d'une main - figure ci-dessous), reliées par une boucle.
Source : PFAM : PF00036 Lors de la fixation du calcium, l'hélice F passe d'une conformation "fermée" (apo-CaM) à une conformation "ouverte" (holo-CaM). Dans la CaM, les 4 boucles de liaison au calcium inclues dans ces motifs ont des séquences homologues chargées négativement et fortement polaires : [D,N] - x - D - [G,N] - [D,N] - G - [Q,T,Y] - x - x - x - x - E |
|
7 atomes d'oxygène constitue le réseau de coordination du calcium :
|
|
Géomètrie des ligands du calcium dans un motif "EF - hand":
Source : Lewit-Bentley & Réty (2000) |
| positions | ligand |
| X et Y | chaînes latérales des acides aminés Asp et Asn |
| Z | chaînes latérales des acides aminés Asp, Asn et Ser |
| - Y | oxygène du groupement carbonyle de la liaison peptidique |
| - X | molécule d'eau |
| -Z | bidentate ou chaînes latérales des acides aminés Asp et Glu |
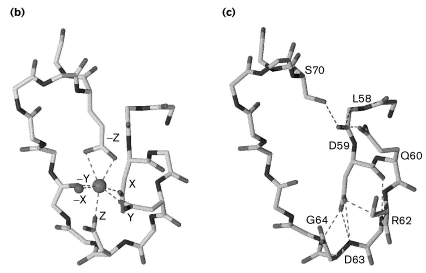
c. Structure de l'apo-calmoduline ou calmoduline non liée au calcium La structure de l'apo-CaM a été obtenue par différentes expériences de RMN, le marquage réverse de résidus Phe* et la mesure de couplage de liaison 3J (3 liaisons séparent 2 noyaux). Les acides aminés Met76 à Ser81, qui forment le lien flexible entre le domaine N-terminal et le domaine C-terminal, sont sous forme hélicoïdale dans la structure de l'apo-CaM, mais pas dans la structure de la CaM liée au calcium. Cette conclusion est corroborée par l'hydrolyse (par des protéases) de cette région de la CaM liée au calcium mais pas de l'apo-CaM, ce qui signifie que cette région est protégée dans l'état non liée au calcium. Ces résultats suggèrent que lorsque le calcium se fixe aux domaines de fixation du calcium, un changement de conformation modifie la structure hélicoïdale de la CaM : ce changement de conformation crée des poches hydrophobes à la surface des domaines de fixation du calcium qui ne se trouvaient pas dans l'état apo-CaM. *Marquage réverse : des bactéries sont cultivées sur un milieu supplémenté en 15NH4Cl, en glucose marqué au 13C et en acides aminés non marqués (ces acides aminés ne sont donc pas marqués ultèrieurement). Seuls les acides aminés qui sont synthétisés par les bactéries en utilisant le 13C-glucose comme source de carbone sont marqués. |
|
d. Structure pleinement étendue de la calmoduline liée au calcium de Homo sapiens à une résolution de 1,7 Å Code PDB : 1CLL Acides aminés des 4 motifs "EF - hand" :
Pour faire apparaître de multiples fonctions du menu Jmol :
|
La CaM possède des propriétés de régulation de la fonction des protéines très particulières puisqu'elle interagit avec environ 100 cibles protéiques (enzymes et protéines) distinctes. Comment, sur la base d'une variabilité maximale de seulement 35% des séquences d'acides aminés qui la composent, la calmoduline adopte-t-elle une telle diversité de conformations lui permettant d'interagir avec autant de protéines cibles, c'est-à-dire avec autant de structures tridimensionnelles différentes ? |
|
La structure ci-contre (code PDB : 1PRW) correspond à une autre structure native que la CaM pourrait adopter in vivo.
|
Cette conformation compacte globulaire offre une moindre surface exposée au solvant. Si cette conformation est adoptée in vivo, elle présente des avantages :
Certaines cibles protéiques liées à la CaM sont sous une forme partiellement étendues (exemples : "anthrax adenylyl cyclase exotoxin", "gating domain of Ca2+-activated K+-channel"). En conséquence :
Source : Haiech et al. (2011) |
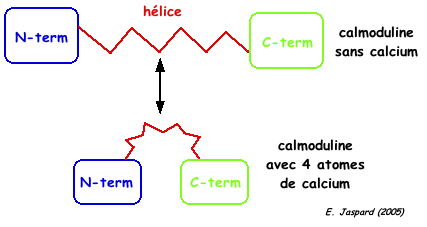
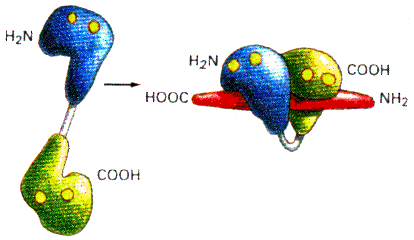
4. Mécanisme de changement de conformation de la calmoduline La fixation des atomes de calcium induit un changement de conformation de la CaM. En particulier, elle modifie la longue hélice centrale. On peut schématiser le changement de conformation qui s'opère sous forme d'une "haltère" (en haut) qui se replie sur elle-même ("pince"- en bas) .
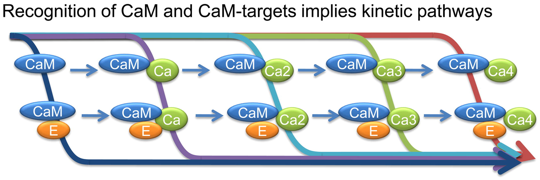
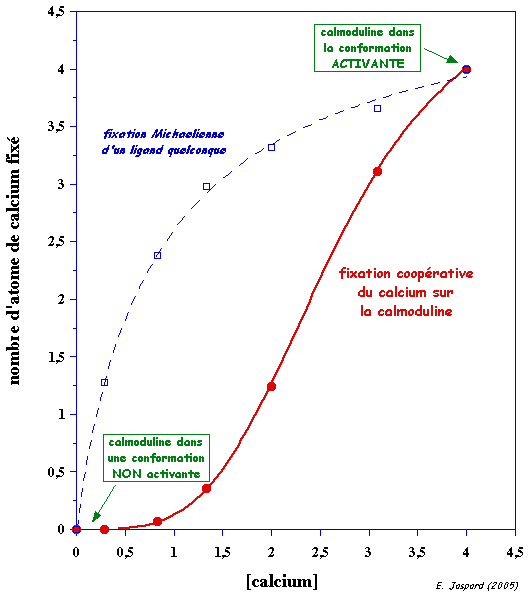
a. Mécanisme lors de la fixation coopérative du calcium apo-CaM (sans calcium) <---> holo-CaM (4 atomes de calcium) La CaM fixe 4 atomes de calcium de manière séquentielle : il y a un effet coopératif (figure ci-dessous) dans la fixation des 4 atomes de calcium (voir le cours sur les protéines à régulation allostérique).
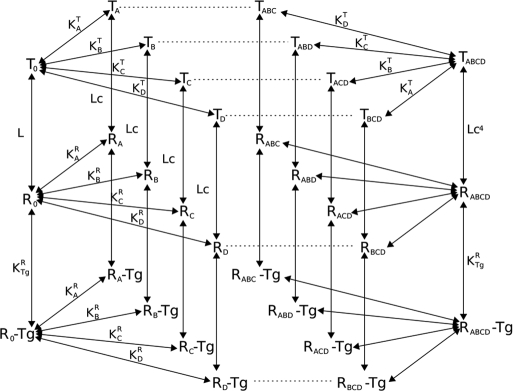
Voir un film sur la transition apo-CaM à holo-CaM. Ce film prend un certain temps à être chargé. Divers modèles mathématiques décrivant la fixation coopérative du calcium par la calmoduline ont été développés.
Source : Stefan et al. (2008)
|
| b. Mécanisme lors de l'interaction avec les
protéines cibles
La CaM adopte alors une conformation activante : c'est dans cette conformation qu'elle interagit avec ses protéines cibles pour en moduler l'activité. La CaM saturée en calcium s'enroule autour de régions spécifiques de la protéine cible.
Voir l'étude des fluctuations conformationnelles de la CaM lors de son repliement : Stigler et al. (2011). Les équilibres ci-dessous décrivent : CaMhaltère + Ca2+ <-> Ca2+-CaM <-> (Ca2+)4-CaMpince EnzCibleinactive + (Ca2+)4-CaMpince <-> [EnzCib)(Ca2+)4-CaM)]active
Bien d'autres modèles théoriques ont été développés pour analyser la spécificité et la sélectivité de la fixation des protéines cibles sur la CaM (voir : Valeyev et al., 2008). Ils ont aussi été développés pour décrypter différents paramètres de l'interaction [CaM - calcium - protéine cible], tels que :
|
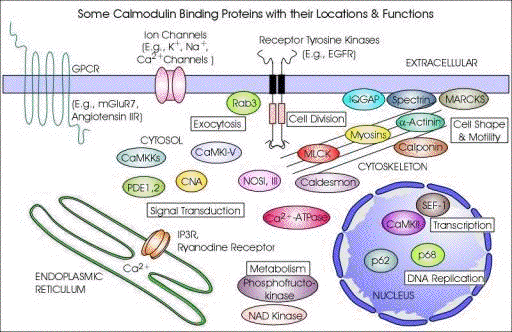
5. Protéines fixant le calcium a. Exemples de complexes [calmoduline-protéine cible] et leurs rôles physiologiques Plusieurs dizaines de types de protéines sont recensées comme interagissant avec la CaM.
Source : EBI - Interpro
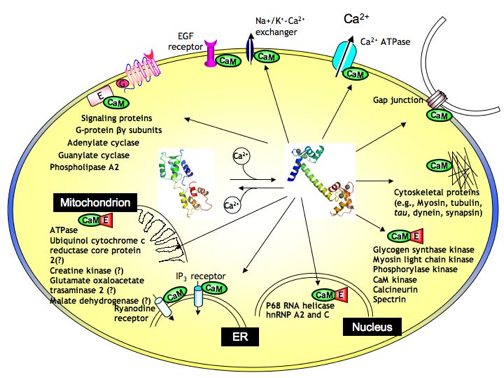
Figure ci-dessous, illustration des interactions de la CaM dans différents compartiments sub-cellulaires : la mitochondrie / le réticulum endoplasmique / le noyau / le cytosquelette
Source : J. Yang Laboratory avec des :
|
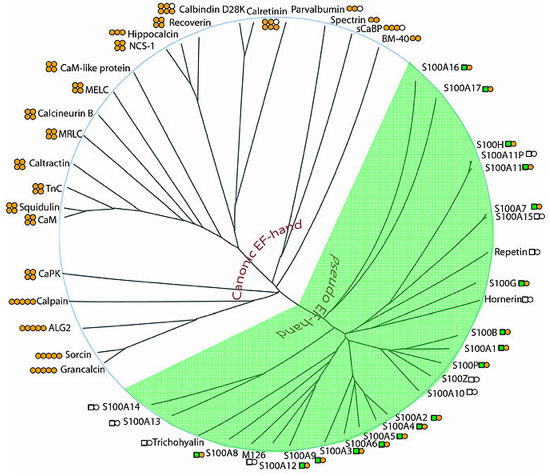
b. Les super-familles de protéines qui fixent le calcium Arbre phylogénétique de la super-famille de protéines fixant le calcium via un domaine structural "EF-hand".
Source : PFAM : PF00036 La super-famille des protéines qui possèdent un motif "EF-hand" contient plus de 66 sous-familles subdivisées en deux groupes principaux: « Canonical EF-hands » dont l'exemple typique est la CaM. « Pseudo EF-hands » : trouvées exclusivement dans le domaine N-terminal des protéines S100 et S100-like. Les 14 acides aminés de ce type de motif fixe le calcium principalement via les groupes carbonyles du squelette carboné. |
Exemples de protéines intracellulaires fixant le calcium et qui contiennent un domaine structural "EF-hand" |
calmoduline parvalbumine (protéine du muscle) super famille des protéines S100 troponine C calcineurines A et B (PP2B et PPP3CA) : seule protéine phosphatase activée par la CaM que l'on connaisse actuellement calretinine (calbindine et calbindine 2) calpaïne caltractine grancalcine chaîne légère et chaîne lourde de myosine spectrine squiduline super famille "Neuronal Calcium Sensor" : frequenine, hippocalcine, recoverine, GCAP, neurocalcine, visinine, ... |
| Exemples de protéines intracellulaires fixant le calcium et qui ne contiennent pas un domaine structural "EF-hand" | calreticuline, calsequestrine annexines protéines qui contiennent un domaine structural C2 : protéine kinase C, sinaptotagmine Calmodulin kinase II (CaMKII) sarcalumenine |
| Exemples de protéines extracellulaires fixant le calcium classées en 6 groupes, selon le type de domaine structural fixant le calcium | 1. domaine "EF-hand" : famille des osteonectines 2. domaine "EGF-like" (EGF : "epidermal growth factor") : facteurs de la coagulation VII, IX et X, protéine C et protéine S, fibrilline, récepteur du LD, récepteurs Notch et delta 3. domaine "γ-carboxyl glutamic acid (GLA)-rich" : facteurs de la coagulation II, VII, IX et X, protéine C et protéine Z, periostine, osteocalcine 4. domaine cadherine : calsyntenines (famille des cadherines) 5. domaine "Ca2+ -dependent (C)-type lectin-like" : selectines, récepteur du mannose, collectines 6. "Ca2+ -binding pockets of family C G-protein-coupled receptors" : récepteur senseur du Ca2+, récepteurs métabotrope du glutamate, récepteurs du GABAB |
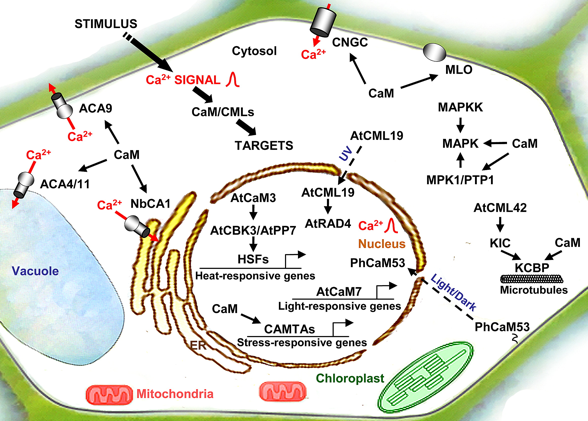
c. Calmoduline et protéines fixant la calmoduline chez les plantes Figure ci-dessous, illustration des interactions de la CaM et autres protéines contenant un ou plusieurs motif "EF-hand" chez les végétaux :
Source : Perochon et al. (2011)
CMLs, CBLs et CDPKs ne sont quasi exclusivement trouvées que chez les plantes et quelques protistes. La superfamille des protéines contenant un ou plusieurs motif "EF-hand" contient environ 250 membres chez le riz (5 gènes CaM et 32 gènes CML) et 250 membres chez Arabidopsis thaliana (7 gènes CaM et 50 gènes CML). Par comparaison, on en dénombre actuellement environ 80 chez l'homme et 130 chez la drosophile. |
| Exemples de protéines de plantes fixant la CaM via des domaines de fixation de la CaM structuralement définis | |
| Phosphorylation - déphosphorylation des protéines | Chimeric Ca2+/CaM-dependent protein kinase Diacylglycerol kinase NAD kinase (NAD2) Cytoplasmic Receptor-like kinase (CRCK1) NtCaMK1 PP2C like phosphatase NPK phosphatase Receptor-like kinase (CRLK1) |
| Protéines nucléaires - régulateurs de la transcription | SRs/CaMTAs AtBT Auxin-induced protein ZmSAUR1 IQD1 AtMYB2 CBP60g AtGT2L Nuclear protein PCBP WRKY group IId AtWRKY7 |
| Métabolisme | Glutamate decarboxylase Catalase FAD-dependent oxidoreductase, DWF1 |
| Protéines membranaires - canaux ioniques - transporteurs |
Cyclic nucleotide gated cation channels (CNCG) |
Plasma membrane Ca2+-ATPase |
|
| Autres | Kinesin-like protein Pollen specific protein (MPCBP, NPG1) Chaperonin 10 DRL1 NADPH-dependent dehydrogenase Tic32 Ubiquitin-specific protease 6 (AtUBP6) |
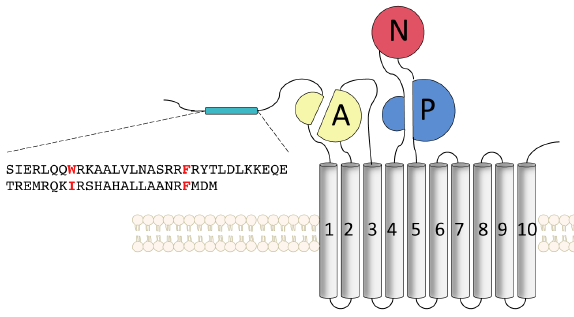
6. Mécanisme d'activation des pompes Ca2+-ATPases par la calmoduline Les ions calcium sont des messagers secondaires de nombreuses voies de transduction du signal. Les cellules investissent beaucoup d'énergie dans le contrôle et le maintien d'un gradient entre la concentration intra-cellulaire (≈ 0.1 μM) et extra-cellulaire (≈ 2 mM) de calcium. La famille des pompes à calcium activées par la calmoduline, à laquelle appartiennent les Ca2+-ATPases de la membrane plasmique ("Plasma-Membrane Ca2+-ATPase"), sont des régulateurs clé de la concentration intra-cellulaire de calcium chez les eucaryotes. Contrairement aux autres ATPases de type P, les Ca2+-ATPases contiennent un domaine N-terminal (chez les plantes) ou C-terminal (chez les mammifères) responsable de l'auto-inhibition. Le domaine N-terminal auto-inhibiteur contient plusieurs acides aminés basiques conservés capitaux pour l'auto-inhibition : Arg58, Arg61, Lys67 et Lys68. Figure ci-dessous : Structure de la Ca2+-ATPase de Arabidopsis thaliana.
Source : Tidow et al. (2012)
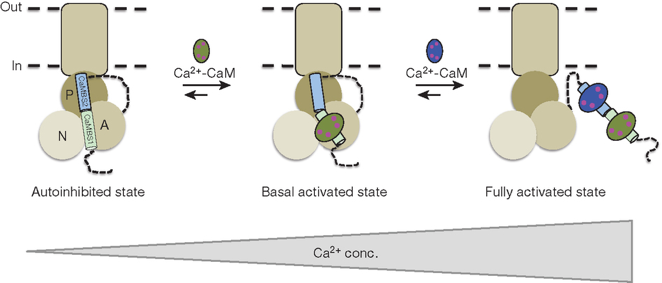
1er temps : quand la concentration en calcium augmente, le complexe Ca2+-CaM se fixe d'abord au premier site de fixation à haute affinité (CaMBS1 - "CaM Binding Site 1"). Cette fixation déplace ce site dans la région entre les domaines A et N et permet les mouvements des domaines cytoplasmiques nécessaires au pompage des ions quand la concentration du calcium est basale. Figure ci-dessous : Mécanisme proposé pour l'activation de la Ca2+-ATPase de Arabidopsis thaliana par la CaM.
Source : Tidow et al. (2012) La fixation de deux molécules de CaM complexée au calcium déplace l'hélice auto-inhibitrice du coeur catalytique, ce qui active la Ca2+-ATPase. 2ème temps : un accroissement supplémentaire de la concentration en calcium déplace le second site CaMBS2 du coeur catalytique : la pompe est alors pleinement activée. Ce mécanisme en 2 temps explique la régulation médiée par la calmoduline sur une telle gamme de concentration de calcium (4 ordres de grandeur). L'analyse des séquences indiquent que l'existence de 2 sites de fixation de la CaM n'est pas propre à Arabidopsis thaliana mais est trouvé chez de nombreuses plantes. |
| 7. Liens Internet et références bibliographiques |
|
"The CaBP Data Library" : Beaucoup d'informations sur la calmoduline et les protéines cibles "Cell signaling" "Calmodulin & EF-Hands proteins" - Interpro |
|
|
Cheung, W. Y. (1970) "Cyclic 3'-5'-nucleotide phosphodiesterase : demonstration of an activator" Biochem. Biophys. Res. Commun. 38, 533 - 538 Means, A. R. & Dedman, J. R. (1980) "Calmodulin : an intracellular calcium receptor" Nature 285, 73 - 77 Lewit-Bentley & Réty (2000) " EF-hand calcium-binding proteins" Curr. Op. Struct. Biol., 10, 637 - 643 |
|
|
Carafoli E. (2002) "Calcium signaling : A tale for all seasons" Proceed. Natl. Acad. Sci. USA 99, 1115 -1122 Clapperton et al. (2002) Biochemistry 41, 14669 -14679 Stefan et al. (2008) "An allosteric model of calmodulin explains differential activation of PP2B and CaMKII" Proc Natl Acad Sci 105, 10768 - 10773 Modèle allostérique de fixation du calcium (base de données "BioModels" - EBI) Stigler et al. (2011) "The Complex Folding Network of Single Calmodulin Molecules" Science 334, 512 - 516 Fallon & Quiocho (2003) "A Closed Compact Structure of Native Ca2+-Calmodulin" Structure 11, 1303 - 1307 |
|
|
Valeyev et al. (2008) "Elucidating the mechanisms of cooperative calcium-calmodulin interactions: a structural systems biology approach" BMC Systems Biol., 2: 48 Perochon et al. (2011) "Calmodulin and calmodulin-like proteins in plant calcium signaling" Biochimie 93, 2048 - 2053 Haiech et al. (2011) "Revisiting intracellular calcium signaling semantics" Biochimie 93, 2029 - 2037 Tidow et al. (2012) "A bimodular mechanism of calcium control in eukaryotes" Nature 491, 468 - 472 |
|
![]()