| L'adénylate cyclase et l'AMP cyclique (AMPc) |
| Tweet |
|
|
1. Les classes et les isoformes de l'adénylate cyclase (ou adénylyl-cyclase) (EC 4.6.1.1) 2. Structure des adénylates cyclases 3. L'adénosine mono-phophate cyclique (AMPc) 4. Activation des protéines kinases A par l'AMPc |
5. Rôle de l'AMPc dans la répression catabolique 6. Dégradation de l'AMPc par la phosphodiesterase 7. Liens Internet et références bibliographiques |
|
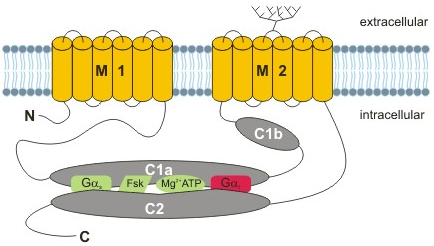
1. Les classes et les isoformes de l'adénylate cyclase (ou adénylyl-cyclase) - EC 4.6.1.1 L'adénylate cyclase a été décrite par Earl Sutherland et Ted Rall en 1962 après leur découverte de l'AMP cyclique comme médiateur de l'effet de l'épinéphrine dans la glycogènolyse (1957) et comme médiateur de l'effet d'autres hormones sur la lipolyse. L'AMP cyclique a été le premier messager secondaire décrit. Nom systématique de l'adénylate cyclase : 3',5'-AMP cyclique synthétase.
|
| Adénylates cyclases de la classe III - Homo sapiens | ||
| Type (le lien renvoie vers Uniprot) | Activation | Inhibition |
Toutes sont activées :
|
Toutes sont inhibées :
|
|
| type 1 (1119 acides aminés) : pourrait être impliquée dans les mécanismes d'apprentissage et de mémorisation |
|
|
| type 2 (1091 acides aminés) |
|
|
| type 3 (1144 acides aminés) |
|
|
| type 4 (1077 acides aminés) |
|
calcium via la calcineurine |
| type 5 (1261 acides aminés) : fortement exprimée dans les tissus cardiovasculaires / régulateurs clé de la rétro-inhibition du rythme cardiaque (comme le type 6) | phosphorylation par la Raf kinase RAF1 et la protéine kinase C |
|
| type 6 (1168 acides aminés) |
|
|
| type 7 (1080 acides aminés) | phosphorylation par la protéine kinase C | |
| type 8 (1251 acides aminés) : pourrait être impliquée dans les mécanismes d'apprentissage et de mémorisation |
|
insensible à la phosphorylation par la protéine kinase C |
| type 9 (1353 acides aminés) | calcium via la calcineurine | |
| type 10 - isoforme soluble (1610 acides aminés) : rôle capital dans la spermatogénèse |
|
|
|
Visualisation du complexe [adénylate cyclase / protéine Gs- sous-unité α] à une résolution de 2,87 Å. Code PDB : 3C16 Le chargement des structures peut prendre un peu de temps. Chaînes représentées :
GSP : 5'-GDP-monothiophosphate
|
|
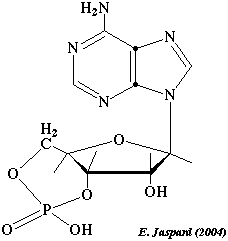
3. L'adénosine mono-phophate cyclique (AMPc) L'adénylate cyclase synthétise l'AMPc (figure ci-dessous) selon la réaction : ATP <=> 3',5'-AMP cyclique + PPi
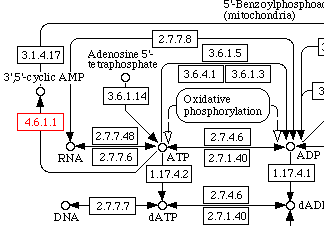
Le groupement phosphate est relié aux carbones 3 et 5 du ribose. La concentration de l'AMPc est controlée par une balance fine entre sa synthèse par l'adénylate cyclase et sa dégradation en 5'-AMP par une phosphodiestérase (voir la PDE ci-dessous). La concentration d'AMPc suit ainsi les variations de concentration d'hormone puisque l'AMPc disparait quand l'hormone n'active plus l'adénylate cyclase. Figure ci-dessous : emplacement de la réaction catalysée par l'adénylate cyclase (EC 4.6.1.1) dans le métabolisme.
Source : KEGG |
|
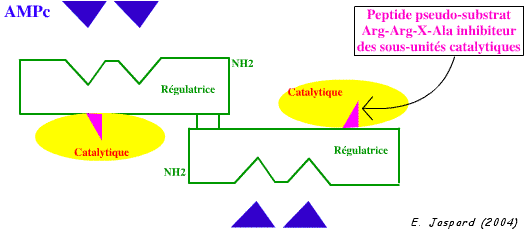
4. Activation des protéines kinases A (PKA) par l'AMPc Les PKA sont constituées de 2 sous-unités catalytiques et de 2 sous-unités régulatrices. Les sous-unités catalytiques phosphorylent les résidus Ser ou Thr présents dans la séquence Arg-Arg-X-Ser/Thr. L'inhibition des sous-unités catalytiques par les sous-unités régulatrices résulte de l'occupation du site catalytique par un peptide pseudo-substrat Arg-Arg-X-Ala, qui ne peut être phosphorylé (il n'y a pas de groupement hydroxyle sur la chaîne latérale de l'Ala).
La fixation d'AMPc sur les 2 sites de chaque sous-unité régulatrice de la PKA induit un changement de conformation de ces sous-unités. Les deux sous-unités catalytiques sont alors relarguées et deviennent actives. Effet sur la glycogènolyse Le glucagon est une hormone peptidique de 29 acides aminés sécrétée par le pancréas : elle stimule la lipolyse et la conversion des acides gras libres en cétones et inhibe la synthèse et favorise la dégradation des protéines.
|
|
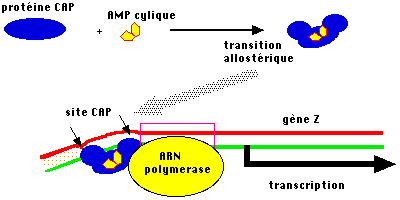
5. Rôle de l'AMPc dans la répression catabolique La transcription de l'opéron lactose est induite si l'on remplace le glucose par du lactose. Inversement, l'opéron n'est pas transcrit tant que le glucose du milieu n'est pas épuisé : c'est ce que l'on appelle la répression catabolique. Des mutations dans les gènes codant pour l'adénylate cyclase ou dans le gène codant pour une protéine appelée CAP ("Catabolite gene Activator Protein") abolissent la répression catabolique. En effet, quand la concentration en glucose diminue, un signal de carence alimentaire est déclenché et se traduit par une augmentation du taux d'AMPc :
Source : R. Jalouzot
|
|
6. Dégradation de l'AMPc par la phosphodiesterase (PDE) La dégradation de l'AMPc est catalysée par la 3',5'-nucleotide cyclique phosphodiesterase (EC 3.1.4.17) selon la réaction : AMPc + H2O <=> 5'-AMP Il existe de multiples formes de PDE chez les Eucaryotes. Ces isoenzymes possède un substrat spécifique et une régulation distincte. |
| isoenzyme de PDE | affinité pour le substrat |
| stimulée par la calmoduline | forte pour GMPc et faible pour AMPc |
| stimulée par le GMPc | faible pour GMPc et AMPc |
| inhibée par le GMPc | forte pour GMPc et AMPc |
| PDE hydrolysant spécifiquement l'AMPc | forte pour AMPc et GMPc |
| PDE hydrolysant spécifiquement le GMPc | forte pour GMPc et AMPc |
| PDE hydrolysant spécifiquement le GMPc | forte pour GMPc |
| PDE hydrolysant spécifiquement l'AMPc | forte pour AMPc |
| Source : V. Lagente et al. - "Potentiels des inhibiteurs de PDE" | |
|
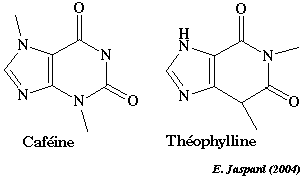
Structure du domaine catalytique de la phosphodiestérase humaine liée à l'AMP (Zhang et al., 2004) : MMDB 28673 - PDB 1TB5 La PDE est inhibée par les méthyl-xanthines comme la caféine ou la théophylline. L'inhibition empèche la dégradation du second messager et prolonge donc le signal et l'activation des effecteurs.
|
| 7. Liens Internet et références bibliographiques |
"Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X |
|
|
Mou et al. (2009) "Structural basis for inhibition of mammalian adenylyl cyclase by calcium" Biochemistry 48, 3387 - 3397 Earl W. Sutherland, Jr. (Prix Nobel 1971) : "for his discoveries concerning the mechanisms of the action of hormones". |
|
![]()