| Les organites des cellules des Eucaryotes |
| Tweet |
|
|
1. La membrane plasmique
2. Le noyau, le nucléole et les ribosomes, les pores nucléaires |
3. Le réticulum endoplasmique 4. L'appareil de Golgi - Exocytose et endocytose 5. Le cytosquelette 6. La mitochondrie 7. Les chloroplastes 8. Les lysosomes, les peroxysomes et les glyoxysomes 9. Liens Internet et références bibliographiques |
Préambule : le projet "Human Cell Atlas" (HCA) On estime que le corps humain contient environ 37.200 milliards de cellules. Le but du HCA est de caractériser toutes les cellules :
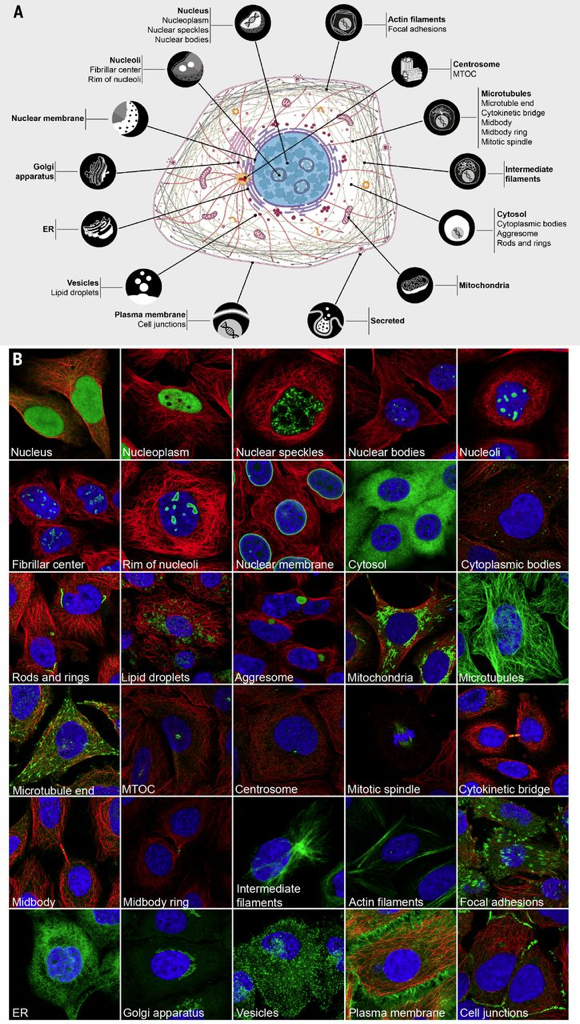
La figure ci-dessous est une carte basée sur la distribution in situ de 12.000 protéines subcellulaires. Cette carte est construite en intégrant la transcriptomique, la microscopie d'immunofluorescence à base d'anticorps et une validation par spectrométrie de masse.
Source : Thul et al. (2017) (A) La cartographie de protéines humaines dans 30 structures subcellulaires d'une seule cellule ("single cell") permet de définir 13 protéomes d'organites majeurs et 1 protéome de protéines sécrétées. (B) Structures subcellulaires visualisées par microscopie à immunofluorescence : protéines (vert); microtubules marqués avec un anticorps anti-tubuline (rouge); noyau contre-coloré au DAPI (bleu). Le côté d'une image représente 64 µm. Base de données :
|
|
1. La membrane plasmique a. Composition et structure La frontière extérieure de la cellule est la membrane plasmique d'une épaisseur de 5 - 6 nm.
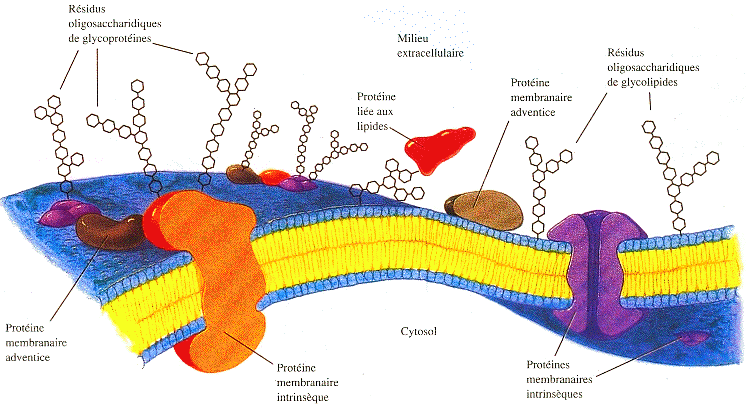
Source : "Principes de Biochimie" Horton et al. (1994) Selon le modèle de la "mosaïque fluide" de Singer et Nicolson (figure ci-dessous), la membrane plasmique est constituée d'une bicouche lipidique, de protéines et de glucides associées de diverses manières à la bicouche.
Source : Singer & Nicolson, 1972 |
| Membrane | plasmique d'érythrocyte | myélinique | microsome | interne de mitochondrie |
| Protéines | 50 | 18 | 62 | 76 |
| Lipides | 42 | 79 | 32 | 24 |
| Glucides | 8 | 3 | 6 | 0 |
|
Les protéines membranaires intrinsèques ou transmembranaires sont composées d'hélices α avec des acides aminés hydrophobes. Cette propriété leur permet d'occuper toute l'épaisseur de la bicouche lipidique en débordant sur chacune des faces de la membrane. Les protéines intrinsèques diffusent latéralement dans la membrane, de manière limitée, ou orientée ou au hasard, selon la protéine (expérience de Frye et Edidin). Celles qui sont ancrées au cytosquelette (côté intra-cellulaire de la membrane) sont immobiles. Les protéines membranaires adventices sont attachées de manière lâche à la surface de l'un des feuillets de la membrane. Il s'agit d'interactions ioniques ou de liaisons hydrogène impliquant les têtes polaires des lipides membranaires. Les chaînes oligosaccharidiques (constituées de : glucose, galactose, mannose, fucose, xylose, galactosamine, glucosamine, acide sialique, acide N-acétylneuraminique, ...) des glycoprotéines et des glycolipides de la membrane plasmique sont situés exclusivement sur sa face extracellulaire. Ces protéines ne passent pas d'un feuillet à l'autre. Les oses représentent 2% à 10% du poids total de la membrane. Les oses à la surface des cellules jouent un rôle important dans la reconnaissance / communication entre les cellules ou entre une cellule et des hormones. Exemples : lectines et activation des lymphocytes, immnuglobulines et réaction avec les antigènes, récepteurs et ligands ... Les protéines liées à un lipide sont attachées par une liaison covalente à la membrane via une ancre glycosyl - phosphatidyinositol. Exemples de fonctions de certaines protéines membranaires :
|
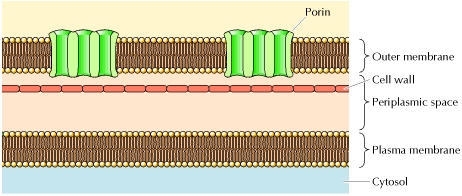
Les protéines de la membrane externe La membrane plasmique de certaines bactéries est elle-même entourée par une enveloppe ("cell wall") puis une membrane externe.
Source : Structure of the Plasma Membrane
On trouve des protéines apparentées aux porines dans la membrane externe des mitochondries et des chloroplastes. |
|
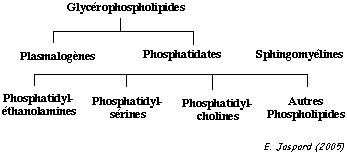
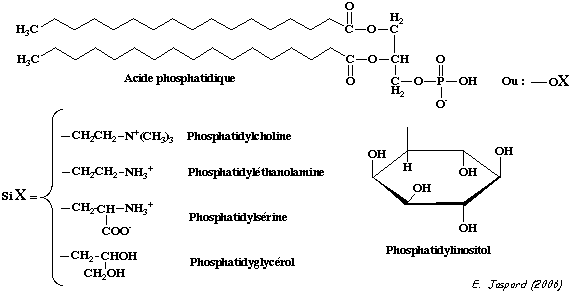
La structure en double couche de la membrane plasmique est due aux propriétés amphiphiles (ou amphipathiques) des multiples types de lipides qui la constituent. Il existe 3 classes principales de lipides membranaires : les phospholipides, les glycolipides et les stérols.
Parmi les les phospholipides, les glycérophospholipides sont les plus abondants. Leur structure est la suivante (figure ci-dessous) :
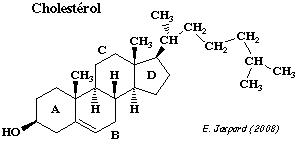
Parmi les lipides membranaires, le cholestérol fait partie de la catégorie des stéroïdes (composés polyprényles, dérivés de l'isoprène) qui contiennent 4 cycles fusionnés (A à D). Le cholestérol est trés bien représenté dans les membranes plasmiques des mammifères mais exceptionnel chez les végétaux et absent chez les procaryotes.
|
| type de membrane (foie de rat) | cholestérol | PC | SM | PE | PI | PS | PG | DPG | PA | GL |
| plasmique | 30 | 18 | 14 | 11 | 4 | 9 | --- | --- | 1 | --- |
| RE rugueux | 6 | 55 | 3 | 16 | 8 | 3 | --- | --- | --- | --- |
| RE lisse | 10 | 55 | 12 | 21 | 7 | --- | --- | 2 | --- | --- |
| mitochondie - interne | 3 | 45 | 2,5 | 24 | 6 | 1 | 2 | 18 | 0,7 | --- |
| mitochondie - externe | 5 | 50 | 5 | 23 | 13 | 2 | 2,5 | 3,5 | 1,3 | --- |
| membrane nucléaire | 10 | 55 | 3 | 20 | 7 | 3 | --- | --- | 1 | --- |
| appareil de golgi | 7,5 | 40 | 10 | 15 | 6 | 3,5 | --- | --- | --- | --- |
| lysosome | 14 | 25 | 24 | 13 | 7 | --- | --- | 5 | --- | --- |
| myéline | 22 | 11 | 6 | 14 | --- | 7 | --- | --- | --- | 12 |
| érythrocyte | 24 | 31 | 8,5 | 15 | 2 | 7 | --- | --- | 0,1 | 3 |
| membrane plasmique - E. coli | 0 | 0 | --- | 80 | --- | --- | 15 | 5 | --- | |
|
PC : phosphatidylcholine / SM : sphingomyèline / PE :
phosphatidyléthanolamine / PI : phosphatidylinositol / PS :
phosphatidylsérine PG : phosphatidylglycérol / DPG : diphosphatidylglycérol (cardiolipine) / PA : acide phosphatidique / GL : glycilipides / RE : réticulum endoplasmique |
||||||||||
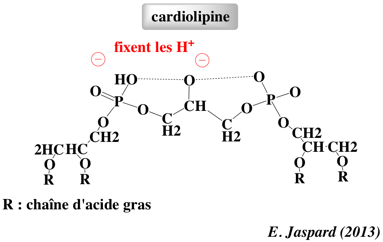
Rôle de la cardiolipine La cardiolipine (diphosphatidylglycérol) est un glycérophospholipide des membranes des bactéries et des mitochondries. La cardiolipine représente 18 à 20% des phospholipides de la membrane interne des mitochondries.
C'est un lipide caractéristique des membranes qui développent un potentiel électrochimique via le transport des électrons pour la synthèse d'ATP. La cardiolipine a de nombreux rôles :
|
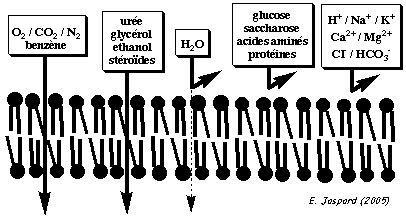
Fluidité des membranes et transposrtsLa bicouche lipidique est fluide car les lipides sont mobiles : mouvement latéral et de rotation. Cependant il y a peu d'échange des lipides d'une couche à l'autre de la membrane : la diffusion d'un lipide d'un feuillet à l'autre est 1 milliard de fois plus lente qu'au sein d'un même feuillet. La fluidité de la membrane dépend du contenu en lipide, de la teneur en cholestérol et de la température. Voir un cours sur la synthèse du cholestérol. La membrane est une barrière très efficace. La figure ci-dessous indique la perméabilité ou non d'une double couche lipidique sans protéine à différents types de biomolécules.
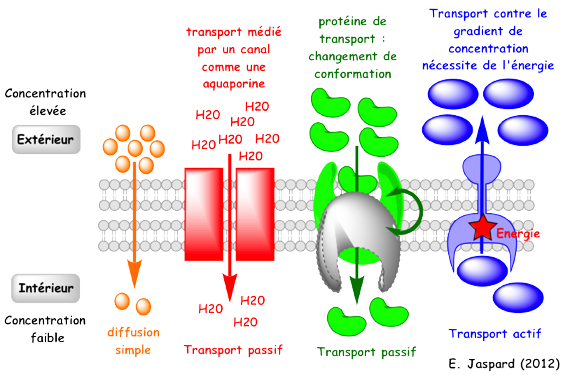
Il existe 3 catégories de transport à travers la membrane :
Figure ci-dessous : illustration schématique des types de transports transmembranaires.
|
| Soluté | Concentration intracellulaire (mM) | Concentration extracellulaire (mM) | Perméabilité (diffusion) de la membrane | Imperméabilité de la membrane |
| Na+ | 12 - 15 | 145 |
|
|
| K+ | 120 - 140 | 4 - 4,5 | ||
| Mg2+ | 0,8 | 1,5 | ||
| Ca2+ | 10-4 | 1,2 - 2 | ||
| H+ | 6,3 108 | 6,3 108 | ||
| Cl- | 5 - 20 | 110 - 116 | ||
| HCO3- / PO43- / protéines / acides nucléiques ... | 148 | 42 |
|
Le passage de matériel biologique trop volumineux ou trop réactif s'effectue par deux autres processus : l'endocytose et l'exocytose. Dans les deux cas, ce transport s'effectue par l'intermédiaire d'une vésicule lipidique, résultat d'une invagination de la membrane plasmique. Voir un cours très complet au NCBI : "Structure of the Plasma Membrane". |
|
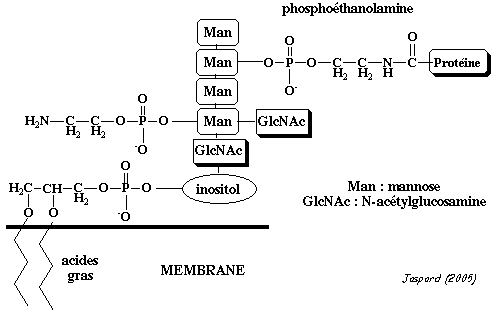
c. Exemples de rôles de lipides membranaires c1. La glypiation et l'ancre glycosyl-phosphatidylinositol La glypiation est la modification post-traductionnelle qui ajoute un groupement glycosyl-phosphatidylinositol ou GPI ou ancre GPI sur un acide aminé en position C-terminale de protéines fixées à la membrane du réticulum endoplasmique. La protéine est fixée à la membrane avant glypiation et le reste après, mais par 2 entités structurales différentes :
Chez Saccharomyces cerevisiae, l'ancrage GPI est essentiel pour la croissance : sur environ 6200 phases de lecture ouvertes ressencées sur son génome, 60 codent des protéines à ancre GPI.
|
|
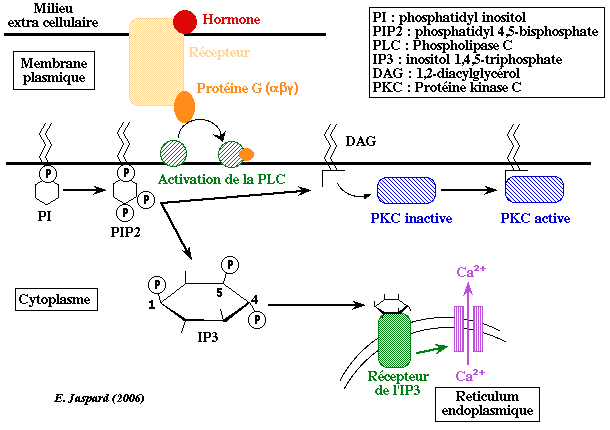
c2. Inositol 1,4,5-triphosphate et diacylglycérol L'hydrolyse d'un phospholipide de la membrane plasmique, le phosphatidyl 4,5-biphosphate (PIP2) par la phospholipase C (une enzyme ancrée à la membrane) forme 2 messagers secondaires (secondaires car il transmettent le signal émis par un messager primaire, l'hormone) : l'inositol 1,4,5-triphosphate (IP3) et le diacylglycérol (DAG). La fonction principale de l'IP3 est la mobilisation du calcium intracellulaire essentiellement stocké dans le réticulum endoplasmique (RE). L'IP3 possède des récepteurs sur la face cytoplasmique de la membrane du RE. Ces récepteurs sont des homotétramères qui forment un canal qui laisse sortir le Ca2+ lorsque chaque monomère fixe une molécule d'IP3. Le Ca2+ cytosolique se fixe à la calmoduline (voir plus haut).
Le DAG provoque : (i) l'augmentation intracytoplasmique du pH (fuite de H+); (ii) l'augmentation intracytoplasmique du taux de Na+; (iii) la phosphorylation des protéines par l'intermédiaire de l'activation de la protéine kinase C. La protéine kinase C est activée par la formation d'un complexe quaternaire [protéine kinase C - DAG - phospholipide - Ca2+]. Le phospholipie le plus efficace pour cette activation est la phosphatidylsérine. Après activation, la protéine kinase C est transferrée du cytoplasme vers la membrane où elle phosphoryle des protéines sur les résidus sérine et thréonine. |
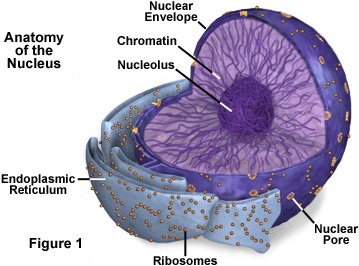
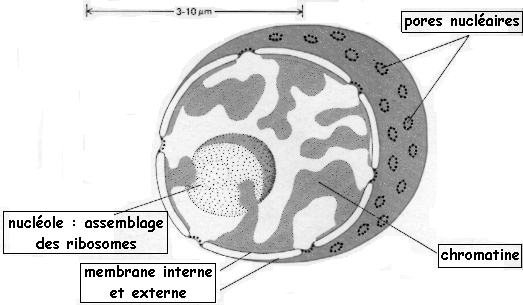
| 2. Le noyau, le nucléole et les ribosomes, les pores nucléaires Le noyau est séparé du cytoplasme par une double membrane, l'enveloppe nucléaire. Celle-ci est continue avec le réticulum endoplasmique. Le contenu du noyau communique avec le cytosol via des pores nucléaires (d'un diamètre de 90 nm). Des structures protéiques trés complexes sont associées aux pores. Elles assurent le transport de certaines macromolécules entre le cytoplasme et la phase aqueuse du noyau, le nucléoplasme.
Tout l'ADN des chromosomes est contenu dans le noyau :
Source : "The Cell Nucleus" |
Le nucléole et les ribosomes Le nucléole est un élément du noyau, dépourvu de membrane (figure ci-dessous), et on peut en trouver plusieurs dans un même noyau.
Source : "Biologie moléculaire de la cellule" Alberts et al. (1983) Le nucléole est le lieu où sont transcrits les ARN ribosomiques. Ces derniers s'associent à des protéines (importées) et ces ensembles forment les deux types de sous unités des ribosomes (petite et grande) :
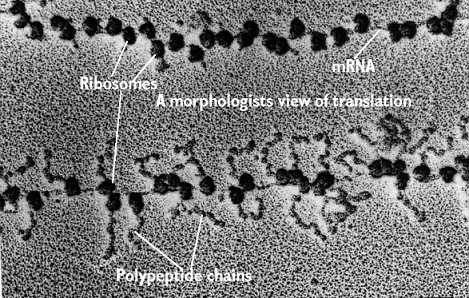
Plusieurs ribosomes peuvent être associés à un même ARN messager et chacun de ces ribosomes synthètise une chaîne polypeptidique (photographie ci-dessous). Ces groupes de ribosomes sont appelés polyribosomes. Ils peuvent être à l'état libre dans le cytoplasme ou liès au réticulum endoplasmique rugueux.
Source : "Role of the Ribosome" - University of Texas Medical Branch |
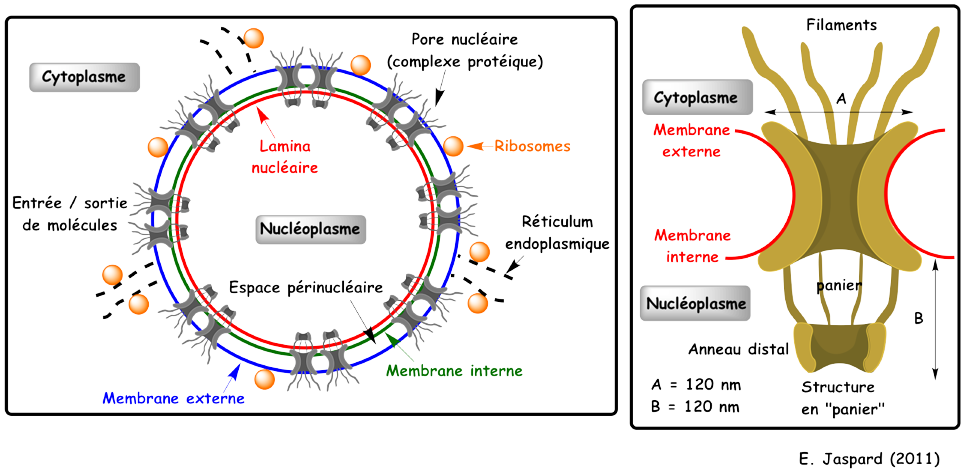
Les pores nucléaires La membrane externe est en continuité avec le réticulum endoplamique rugueux. La membrane interne est associée à un réseau de protéines, la lamina nucléaire (entre la membrane et la chromatine). L'enveloppe nucléaire ne peut être franchie qu'au niveau des pores nucléaires (2000 à 4000 par noyau). Les pores contrôlent l'état des ARN qui sortent du noyau. Les pores ne laissent entrer dans le nucléoplasme que les protéines qui possèdent une séquence signal qui les adresse au noyau ("nuclear localisation signal" - NLS).
Le pore nucléaire de la levure est un complexe protéique (une particule) constitué de 552 chaînes polypeptidiques d'une masse molaire totale estimée à 52 106 Da. Il est constitué de : 8 anneaux, 1 réseau central, 8 rayons, 8 filaments coté cytoplasme, 8 filaments coté nucléoplasme qui adoptent une disposition en panier ("basket"), 1 anneau distal. Les protéines et les ARN qui traversent le pore nucléaire se fixent aux filaments. L'essentiel des protéines qui constituent le pore nucléaire sont des nucléoporines. |
|
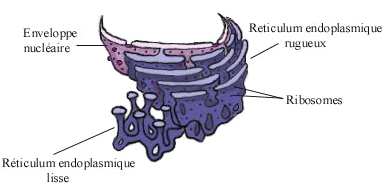
Le réticulum endoplasmique (RE) est une extension de la membrane du noyau. Le RE est divisé en RE lisse et RE rugueux, en fonction de son apparence au microscope. Visualisation du réticulum endoplasmique en 3D.
Source : "L'organisation structurale de la cellule" - M. Forest
Un tiers des protéines totales de la cellule transite par le RE. En voici quelques exemples :
D'autres protéines comme :
Certaines de ces protéines opèrent uniquement dans le RE et y demeurent. D'autres vont dans l'appareil de Golgi. |
|
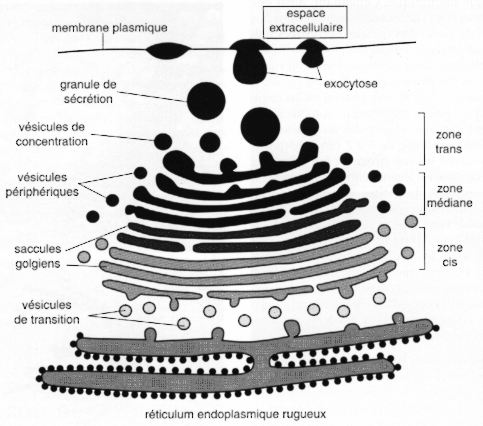
Le transport des macromolécules biologiques est effectué par des vésicules membranaires et selon deux processus. (a) L'exocytose (sortie des macromolécules)
Source : J.C. CALLEN - "Biologie Cellulaire" (1999) (b) L'endocytose (entrée des macromolécules) : des parties de la membrane plasmique s'invaginent vers l'intérieur de la cellule. Elles forment des vésicules d'endocytose. La cellule peut ainsi faire entrer des molécules. Les lysosomes sont des vésicules intracellulaires qui contiennent des enzymes de dégradation. Ces vésicules fusionnent avec les vésicules de phagocytose et leur contenu est dégradé. La cellule récupère ainsi les éléments de base constitutifs des macromolécules. |
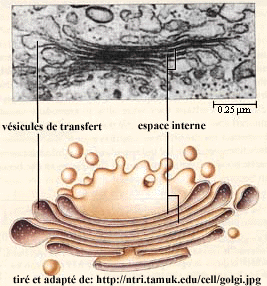
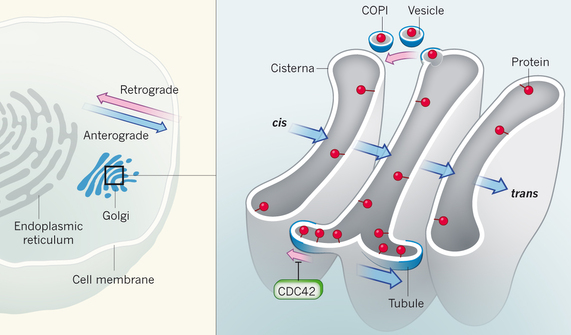
Transport des protéines au travers de l'appareil de Golgi L'appareil de Golgi contient des sacs membranaires empilés appelées citernes. L'appareil de Golgi effectue les opérations de maturation des protéines afin qu'elles soient, par la suite, correctement dirigées vers leur localisation finale dans la cellule. Les protéines peuvent être transportées de 3 façons à travers l'appareil de Golgi.
Source : Nakano A. (2015)
La protéine CDC42 est une petite GTPase qui contribue à polariser la cellule. CDC42 est en concurrence avec le complexe COPI pour le chargement de protéines dans les tubules pour le transport rétrograde (mais pas le transport antérograde) : la protéine CDC42 augmente le transport antérograde et inhibe le transport rétrograde. |
|
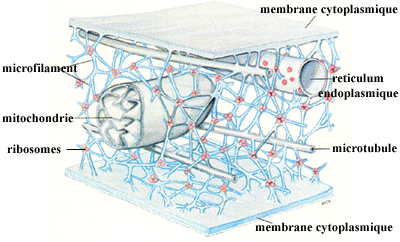
Le cytosquelette a diverses fonctions : forme de la cellule, division de la cellule, défense de la cellule, mouvements de la cellule et mouvements à l'intérieur de la cellule, systèmes de transports intracellulaires.
Source : "L'organisation structurale de la cellule" - M. Forest Il est constitué principalement de 3 types de protéines qui forment une sorte d'échaffaudage : les filaments d'actine ou microfilaments, les filaments dits intermédiaires, les microtubules. |
|
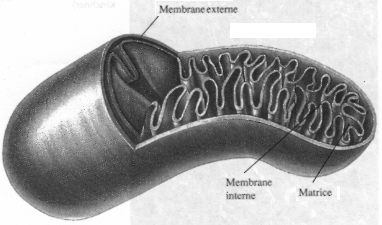
La mitochondrie joue un rôle important dans le métabolisme de la cellule. Dérivées des bactéries, elle contient son propre ADN (ADN mitochondrial). Certaines protéines mitochondriales sont synthétisées par la mitochondrie, mais la plupart d'entre elles sont codées par le génome nucléaire et importées dans la mitochondrie. La mitochondrie est limitée par deux membranes de propriétés très différentes :
Source : "Principes de Biochimie" Horton et al. (1994) La zone interne de la mitochondrie (bordée par la membrane interne) s'appelle la matrice. Elle contient les enzymes du cycle de Krebs et la plupart de celles qui catalysent l'oxydation des acides gras. Voir un développement sur la mitochondrie. |
|
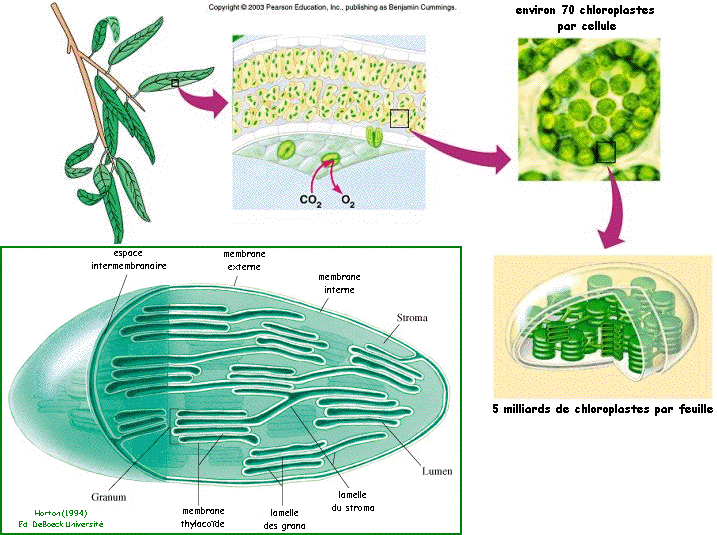
La photosynthèse a lieu dans les chloroplastes. Ce sont des organites spécialisés. Deux membranes (interne et externe) bordent une zone aqueuse appelée stroma (siège de la réduction du CO2 en glucides). Le stroma contient la membrane thylacoïde (siège des réactions lumineuses). La membrane thylacoïde est plissée en un réseau de nombresuses vésicules aplaties qui prend la forme soit d'empilements compacts appelés grana (granum), soit isolées et libres dans le stroma et réunissant plusieurs grana.Les parties de membrane thylacoïde situées au sein des grana et sans contact avec le sont les lamelles des grana baignent dans le stroma sont les lamelles du stroma. L'espace interne enclos par la membrane thylacoïde est le lumen.
Source : "Principes de Biochimie" Horton et al. (1994) - Ed. DeBoeck Université - Pearson Education, Inc. |
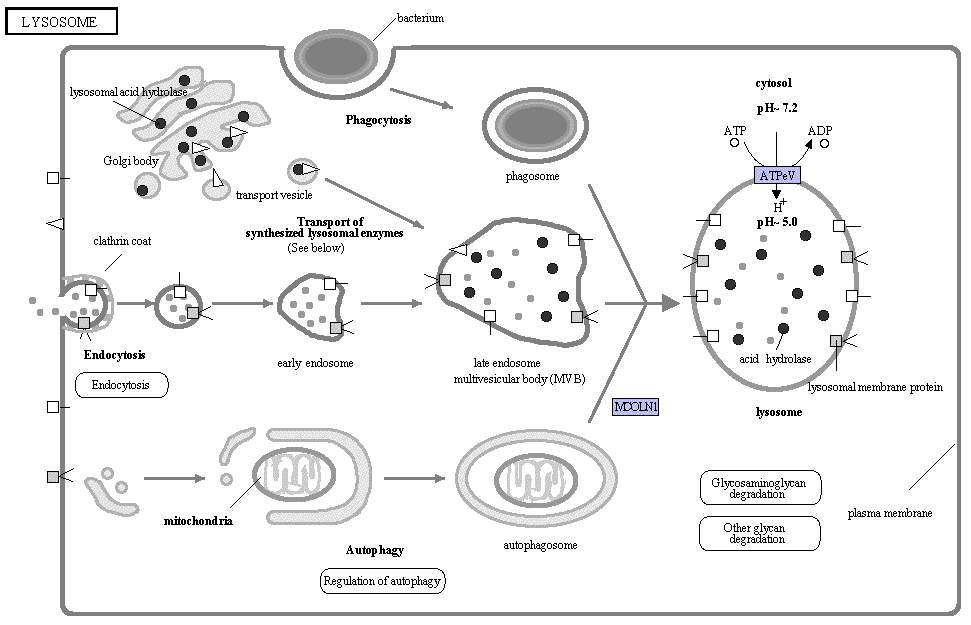
8. Les lysosomes, les peroxysomes et les glyoxysomes Les lysosomes (du grec lysis, dissolution) sont des organites de 0,2 à 0,5 µm (taille et forme variable) qui se trouvent dans le cytosol et sont formés dans l'appareil de Golgi. On les trouve dans toutes les cellules eucaryotes animales, à part les hématies. En revanche, les cellules qui possèdent une paroi extracellulaire comme les cellules végétales n'ont pas de lysosomes. C'est leur vacuole qui joue un rôle similaire.
Source : KEGG La figure du site KEGG est interactive et permet d'accéder à un trés grand nombre d'informations sur les constituants (processus divers tels que l'autophagie, l'endocytose, les diverses enzymes des lysosomes, leurs membranes ...). La membrane des lysosomes contient des pompes à protons (qui utilisent l'énergie de l'ATP) et des protéines de transport. En effet, le pH des lysosomes est maintenu entre 3,5 et 5 (le pH du cytosol étant d'environ 7,2) par ces pompes à protons (H+) et par des canaux ioniques spécifiques des ions chlorures (Cl-). Le pH des lysosomes est acide afin que les enzymes hydrolytiques acides qu'ils contiennent puissent fonctionner. Ces hydrolases acides dégradent les macromolécules et des particules ingérées par la cellule. |
| Hydrolases | Cibles | Hydrolases | Cibles |
| phosphatase acide et phosphodiestérase acide | oligonucléotides | cathepsines | protéines |
| ribonucléase acide et désoxyribonucléase acide | ARN et ADN | collagénase | collagène |
| α-galactosidase, β-galactosidase, β-glucosidase, β-mannosidase,... | galactosides, glycogène, glycoprotéines, ... | peptidase | peptides |
| lysozyme | parois cellulaires des bactéries | estérases | esters d'acides gras |
| hyaluronidase | acide hyaluronique | phospholipases | phospholipides |
|
Il existe bien évidemment un trés grand nombre d'autres enzymes impliquées dans divers processus :
|
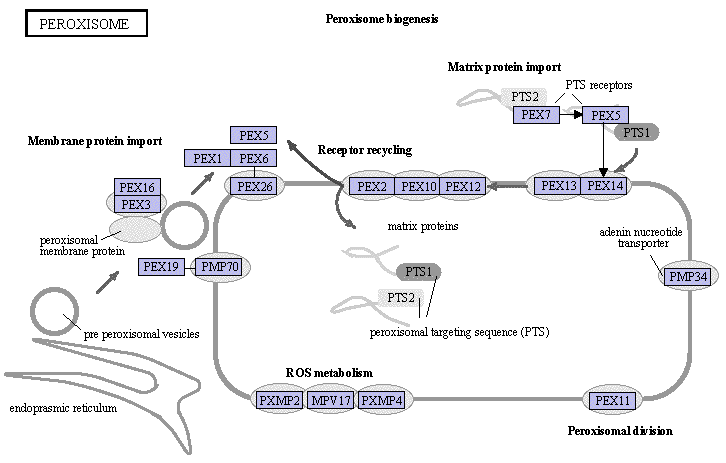
Les peroxysomes sont de petites vésicules entourée d'une membrane. Leur fonction métabolique majeure est la β-oxydation de longues chaînes d'acides gras (supèrieures à 18 carbones) qui, en formant des chaînes plus courtes, alimente la béta β-oxydation mitochondriale. Le nom "peroxysome" est lié à la première étape d'oxydation des acides gras qui se traduit par la formation de peroxyde d'hydrogène (H2O2) . Les peroxysomes sont aussi impliqués dans la synthèse de la bile, du cholestérol et dans le métabolisme des acides aminés et des purines. Par ailleurs, ils jouent un rôle fondamental dans le maintien du pouvoir rédox de la cellule et la détoxification des radicaux libres, dérivés de l'oxygène :
Source : KEGG - La figure du site KEGG est interactive et permet d'accéder à un trés grand nombre d'informations. Les glyoxysomes sont des organites semblables aux peroxysomes chez les végétaux. Ils contiennent en plus les enzymes du cycle du glyoxylate qui transforme les réserves lipidiques des graines en cours de germination en glucides qui alimentent l'embryon pendant la germination. |
| 9. Liens Internet et références bibliographiques |
|
"Biologie cellulaire : exercices et méthodes" Thiry et al. (2014) - DUNOD - ISBN : 978-2-10-070711-9 "Principes de Biochimie" Horton et al. (1994) - Ed. DeBoeck Université - Pearson Education, Inc. |
|
|
Singer, S.J. & Nicolson G. L. (1972) "The fluid mosaic model of the structure of cell membranes" Science 175, 720 - 731 "L'organisation structurale de la cellule" - M. Forest (Canada) |
|
|
"La cellule, ses organites et leurs fonctions" - Excellent cours interactif (Université de Bordeaux) The Cell Image Library |
|
|
Nakano A. (2015) "Cell biology: Polarized transport in the Golgi apparatus" Nature 521, 427 - 428 Thul et al. (2017) "A subcellular map of the human proteome" Science 356, eaal3321 Kim et al. (2018) "Integrative Structure and Functional Anatomy of a Nuclear Pore Complex" Nature 555, 475 - 482 |
|
|
Rozenblatt-Rosen et al. (2021) "Building a high-quality Human Cell Atlas" Nat. Biotechnol. 39, 149 - 153 Heinrich et al. (2021) "Whole-cell organelle segmentation in volume electron microscopy" Nature 599, 141 - 146 |
|
![]()