La réponse au stress lié à l'accumulation de protéines mal repliées dans le réticulum endoplasmique |
| Tweet |
|
|
1. Adressage des protéines vers le réticulum endoplasmique 2. Peptide signal et particule de reconnaissance du peptide signal ("Signal Recognition Particle" - SRP) 3. Le canal de translocation des protéines 4. Le cas des protéines glycosylées 5. Présentation de l'UPR 6. La voie traductionnelle et la voie transcriptionnelle
|
7. La protéine chaperonne BiP/GRP78 8. Le facteur de transcription XBP1 9. La voie de dégradation ERAD 10. Aperçu de l'apoptose résultant de l'UPR 11. Liens Internet et références bibliographiques |
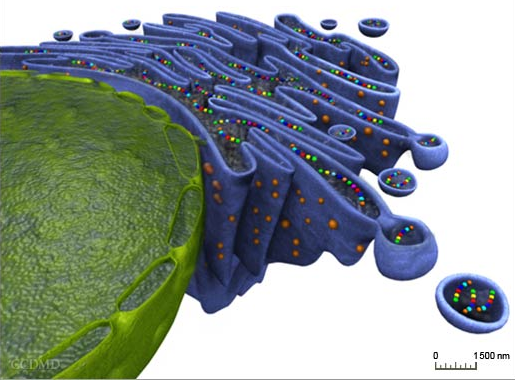
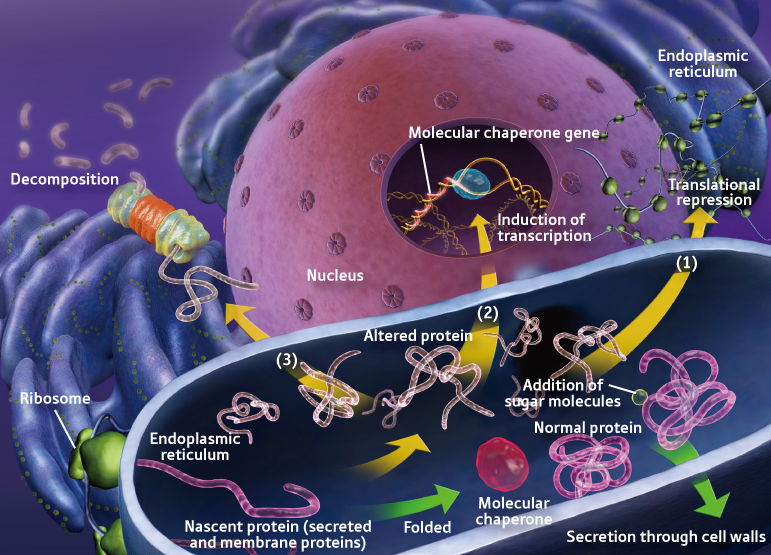
1. Adressage des protéines vers le réticulum endoplasmique Lors de leur biosynthèse, un tiers des protéines d'une cellule sont dirigées vers le réticulum endoplasmique (RE) pour y être repliées , et y subir un contrôle de leur repliement correct et des maturations post-traductionnelles diverses.
Source : La cellule animale Voir le réticulum endoplasmique en 3D. Voici quelques exemples de protéines acheminées vers le RE :
Le repliement des protéines est un processus oxydatif qui génère des espèces radicalaires de l'oxygène activées ("Reactive Oxygen Species" - ROS). Les ROS peuvent cibler les protéines chaperonnes et les canaux ioniques calciques du RE, ce qui conduit au relarguage de calcium du RE vers le cytoplasme et induit des voies de signalisation de stress. Le calcium relargué du RE est alors concentré dans la matrice mitochondriale, où il bloque la chaîne de transport des électrons, ce qui a pour conséquence une production accrue de ROS. Ces ROS produites par la mitochondrie peuvent, en retour, stimuler le relarguage de calcium du RE. Il en résulte une accumulation de ROS qui atteignent un niveau de concentration toxique. De plus, la perturbation de l'homéostasie calcique du RE peut abolir le repliement des protéines effectué dans ce compartiment, ce qui induit un stress du RE qui a pour conséquence une génération encore accrue de ROS. |
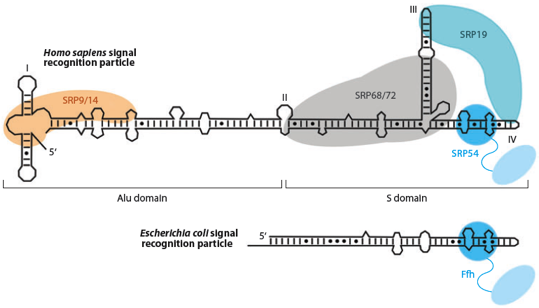
2. Peptide signal et particule de reconnaissance du peptide signal ("Signal Recognition Particle" - SRP) Certaines chaînes polypeptidiques commencent par une séquence appelée peptide signal (ou séquence d'adressage) qui indique à la cellule le compartiment vers lequel les adresser. Günter Blobel a reçu le Prix Nobel en 1999 pour sa découverte que les protéines ont des signaux intrinsèques qui régissent leur transport et leur localisation dans la cellule. Chez les procaryotes et les eucaryotes, un complexe appelé particule de reconnaissance du peptide signal ("signal recognition particle" - SRP) se fixe au peptide signal qui émerge du ribosome. Le complexe SRP, le ribosome et la RNAse P sont les 3 particules ribonucléoprotéiques conservées dans tous les règnes du vivant. Chez les eucaryotes, le complexe SRP peut être structuralement et fonctionnellement divisé en 2 domaines :
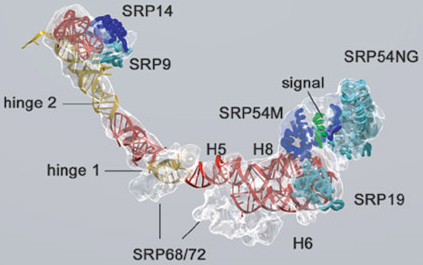
Source : Akopian et al. (2013) Figure du bas, modèle chez les procaryotes (Escherichia coli) : interaction avec la protéine Ffh. Figure ci-dessous : structure de l'ARN 7S, des protéines SRP et du peptide signal.
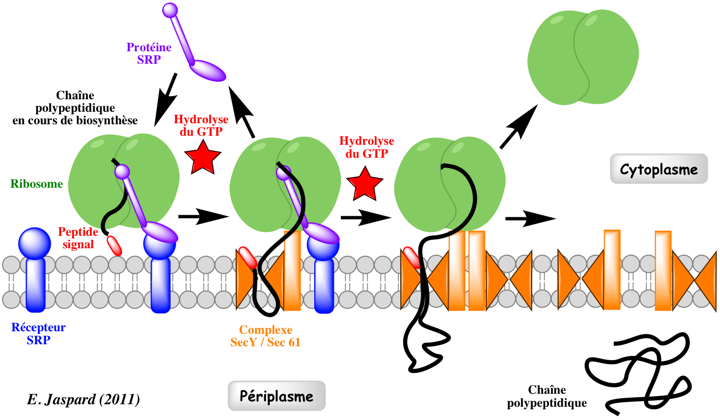
Source : Halic & Beckmann (2005) Dans les chloroplastes des plantes supérieures, SRP a connu de profonds changements liés à l'évolution. A la place d'un ARN, il semble utiliser une protéine spécifique du chloroplaste : cpSRP43. Cependant, des homologues de SRP54 et du récepteur SRP sont présents dans les chloroplastes. Le fonctionnement des chloroplastes implique environ 3000 protéines. Or l'ADN chloroplastique ne code que pour environ 100 protéines. Les chloroplastes importent donc la très grande majorité de leurs protéines. Il en est de même pour la mitochondrie. Mécanisme Le complexe SRP ralentit l'élongation de la chaîne polypeptidique en cours de biosynthèse ("elongation arrest"). Le complexe SRP se fixe au récepteur du SRP : il adresse ainsi les protéines à la membrane plasmique chez les procaryotes ou à la membrane du RE chez les eucaryotes.
Le complexe [ribosome - chaîne polypeptidique en cours de biosynthèse] est délivré à la membrane cible via l'interaction du complexe SRP avec le récepteur de SRP. Ce complexe est transféré au translocon Sec61p chez les eucaryotes ou au translocon secYEG chez les bactéries et les archées (voir ci-dessous). La chaîne polypeptidique en cours d'élongation emprunte le canal pour passer entièrement dans la lumière du RE (dans le cas d'une protéine sécrétée, dirigée vers l'appareil de Golgi) ou pour être intégrée à la membrane (dans le cas d'une protéine membranaire). Le complexe SRP et le récepteur de SRP se dissocient pour participer à la translocation d'une autre chaîne polypeptidique en cours de biosynthèse Il s'agit donc d'un transport co-traductionnel puisque l'élongation de la chaîne polypeptidique continue pendant le passage dans le RE. Le cycle SRP est un processus dont l'énergie est fournie par l'hydrolyse du GTP : le complexe SRP et le récepteur du SRP possèdent une activité GTPase. Voir une belle vidéo du mécanisme : "Signal-Recognition-Particle". |
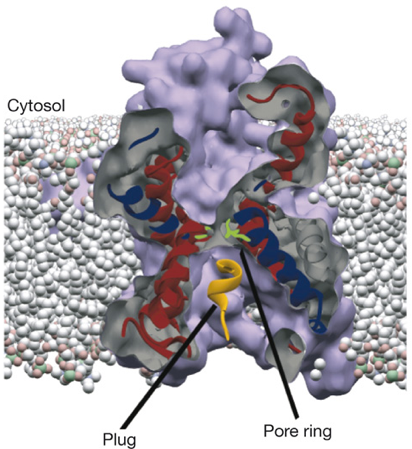
3. Le canal de translocation des protéines Le transit des protéines dans le RE s'effectue via un transporteur, situé dans la membrane du RE, appelé canal de translocation des protéines ("Protein Conducting Channel" - PCC) ou translocon (eucaryotes). La chaîne polypeptidique transportée doit contenir au moins 70 acides aminés pour entrer : 40 acides aminés se trouvent au niveau du ribosome et 30 acides aminés sont dans le canal de translocation. Ce pore fonctionne dans l'autre sens pour éliminer les protéines mal repliées : c'est le transport rétrograde. Le canal de translocation est constitué de plusieurs protéines qui forment un pore (d'environ 4 nm de diamètre) qui permet le passage de la chaîne polypeptidique en cours de biosynthèse, du ribosome vers la lumière du RE. Le canal de translocation est constitué de SecY ou Sec61α (selon le règne) qui est la sous-unité principale avec 10 domaines transmembranaires et de plus petites sous-unités localisées en périphérie de ce canal.
Figure ci-dessus : structure 3D de SecYEG (PDB 2AKI). Source : Rapoport T.A. (2007) Les nomenclatures des sous-unités sont les suivantes :
|
|
Visualisation du canal de translocation des protéines de Escherichia coli obtenue par cryo-microscopie électronique à une résolution de 14,9 Å (2005) Code PDB : 2AKI
|
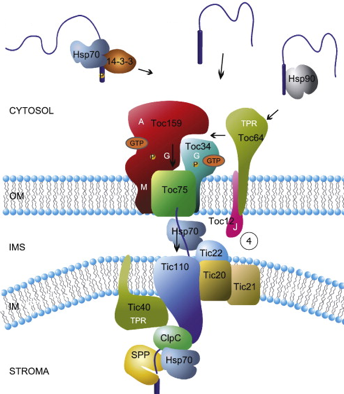
Les chloroplastes possèdent :
Source : Andrès et al. (2010) Les peptides signaux des protéines importées dans le chloroplaste sont reconnus par plusieurs protéines cytosoliques : Hsp70 et Hsp90 (au moins in vitro) ou des protéines 14-3-3. |
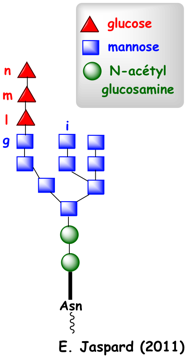
4. Le cas des protéines glycosylées La glycosylation a lieu dans le RE et la coiffe glucidique ajoutée est ensuite modifiée dans l'appareil de Golgi. L'oligosaccharyl transférase (EC 2.4.1.119), associée au translocon Sec61, retarde le repliement de la chaîne polypeptidique naissante pour faciliter le transfert d'un oligosaccharide (figure ci-dessous).
Cet oligosaccharide est composé de :
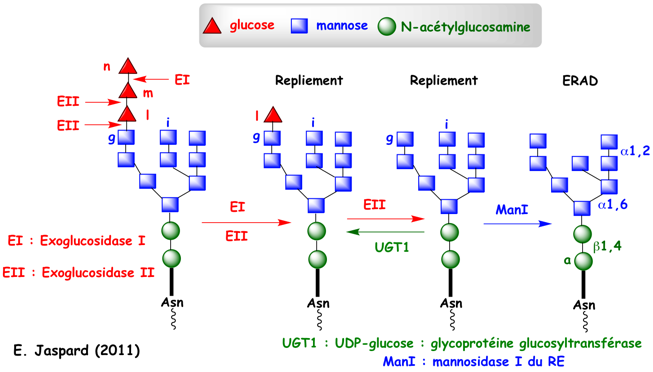
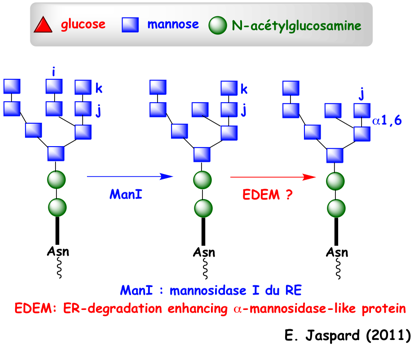
La fixation d'oligosaccharides (Glc3-Man9-GlcNAc2) aux chaînes polypeptidiques augmentent la solubilité des chaînes naissantes non encore repliées et facilite la maturation des protéines. Immédiatement après l'addition de cet oligosaccharide, les 2 résidus glucose n et m sont hydrolysés respectivement par l'exoglucosidase I puis par l'exoglucosidase II (figure ci-dessous).
Les protéines liées à des oligosaccharides monoglucosylés (Glc1-Man9-GlcNAc2) bénéficient alors d'un ensemble de protéines pour leur repliement :
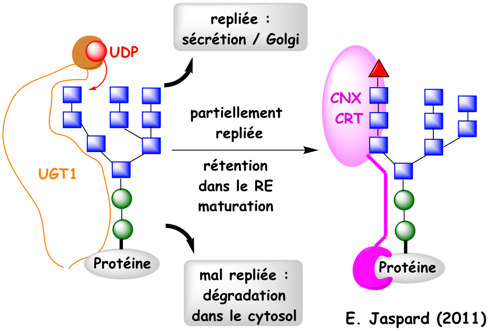
Au moment du relarguage de CNX et CRT, le glucose l est hydrolysé par l'exoglucosidase II. Ces étapes sont sous le contrôle d'une enzyme capitale : l'UDP-glucose:glycoprotéine glucosyltransférase (UGT1). Cette enzyme re-glycosyle spécifiquement le mannose terminal g des chaînes polypeptidiques mal repliées (figure ci-dessous), les renvoyant ainsi pour un nouveau cycle d'essai de repliement (en association avec CNX).
UGT1 est une grosse protéine soluble de 1555 acides aminés (170 kDa) chez l'homme. Son domaine N-terminal est le senseur du repliement qui effectue le contrôle qualité des glycosylations correctes. Remarque : on recense plus de 260 hexosyltransferases (EC 2.4.1.). Ce cycle s'achève quand la mannosidase I du RE hydrolyse le mannose i (Man8-GlcNAc2 - figure ci-dessus) ce qui évite une réassociation avec UGT1. A l'issue de ces différentes vérifications :
|
|
Divers stimuli qui perturbent l'homéostasie du RE peuvent être cause d'un mauvais repliement des protéines :
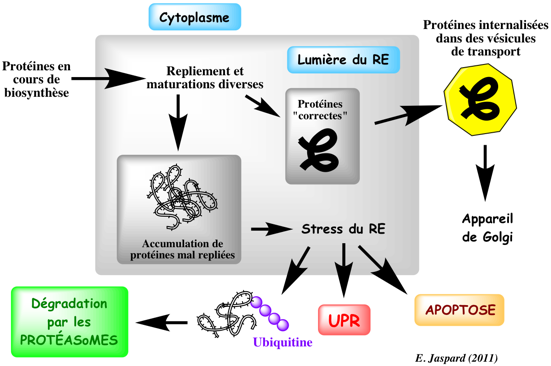
Le RE est donc le compartiment dans lequel les protéines (transmembranaires, sécrétées ou qui y résideront) sont repliées avec l'aide (ou non) de protéines chaperonnes puis maturées par des modifications post-traductionnelles. Quand le repliement des protéines est inhibé ou qu'il se fait mal, des voies de signalisation sont activées et elles ont pour buts :
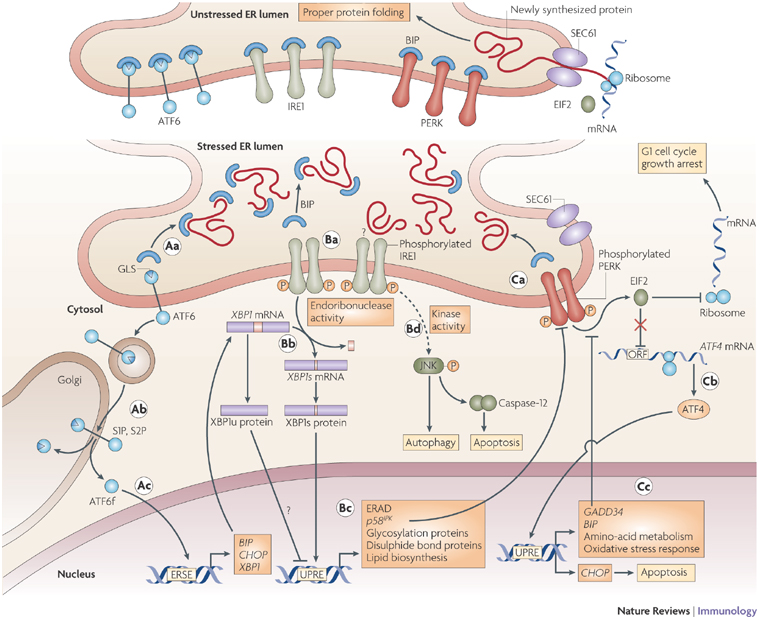
Source : Iwawaki T. (2009) Cet ensemble de voies de signalisation qui est une réponse adaptative physiologique de la cellule à l'accumulation de protéines mal repliées s'appelle "Unfolded Protein Response" ou UPR.
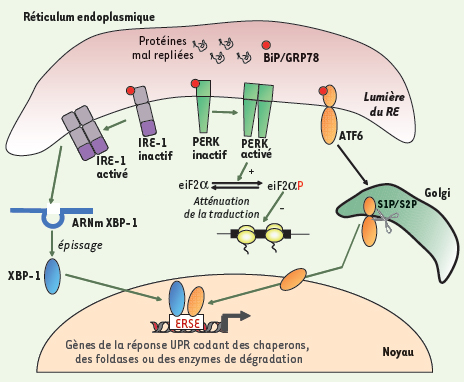
Source : Foufelle & Ferré (2007) - ERSE ("ER stress-response element") : séquences des promoteurs de gènes de l'UPR. Chez les mammifères, l'UPR inclue 3 voies de signalisation impliquant trois protéines transmembranaires du RE :
La levure ne possède que la voie IRE-1 : l'UPR y active cependant la transcription de près de 400 gènes. Voir une carte interactive de l'UPR au KEGG ("Kyoto Encyclopedia of Genes and Genomes"). |
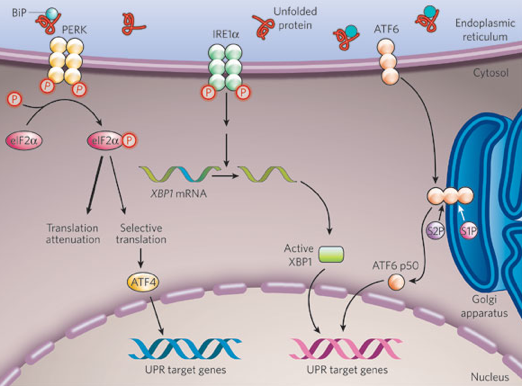
6. La voie traductionnelle et la voie transcriptionnelle Chez Saccharomyces cerevisiae, l'UPR active la transcription de gènes qui codent les protéines chaperonnes, les protéines de glycosylation et les protéines de la voie de dégradation ERAD. Chez les mammifères, l'UPR est un processus plus complexe puisque 2 voies sont activées (figure ci-dessus). Une voie traductionnelle L'interaction des protéines PERK, ATF6 et IRE-1 avec la protéine chaperonne BiP/GRP78 les maintient inactives. L'accumulation de protéines mal repliées entraîne l'activation des trois protéines et l'UPR. ATF6 migre vers l'appareil de Golgi. PERK se dimèrise stimulant ainsi son activité kinase qui phosphoryle (Ser 51) la sous-unité α du facteur d’initiation de la traduction eIF2 : la traduction est alors inhibée. Cependant, la traduction de l'ARN messager codant le facteur de transcription ATF4 qui participe à l'UPR est augmentée. En effet, il contient des phases de lecture ouvertes en amont ( "upstream open reading frame" - uORF) dans sa partie 5' non traduite. Ce type de phases de lecture permet l'expression de cet ARN messager malgré l'inhibition de la synthèse protéique. Un uORF est composé d'un uAUG, d'au moins un codon non-stop et d'un codon stop en phase en aval de l'uAUG. Une voie transcriptionnelle ATF6
Source : Zhang & Kaufman (2008) IRE-1
L'épissage d'ARN messager dans le cytoplasme est l'un des mécanismes régulateurs majeurs de l'UPR. Le mécanisme moléculaire de l'épissage cytoplasmique est unique et complètement différent de l'épissage conventionnel qui a lieu dans le noyau. Chez les mammifères, le substrat de cet épissage cytoplasmique de l'UPR est l'ARN pré-messager codant le facteur de transcription XBP1. La forme épissé de l'ARN messager qui en résulte aboutit à la synthèse de la forme active XBP1s. Cependant, l'ARN pré-messager codant XBP1 est aussi traduit sous la forme d'une protéine XBP1u qui régule négativement (répresseur) l'UPR. En conséquence, les cellules de mammifères peuvent s'adapter très rapidement aux changements de conditions qui règnent dans le RE en basculant d'une forme XBP1 répresseur de la transcription à une forme activatrice de la transcription. |
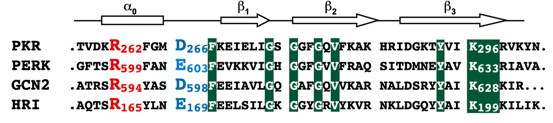
a. La protéine PERK ("double-stranded RNA-activated protein kinase (PKR)-like Endoplasmic Reticulum Kinase") La dimèrisation du domaine N-terminal de PERK (situé dans la lumière du RE) induit l'autophosphorylation de différents acides aminés de son domaine kinase C-terminal, dont Thr 980 située dans la boucle d'activation kinase. La phosphorylation de Thr 980 stabilise cette boucle d'activation et l'hélice αG du lobe C-terminal, rendant cette hélice apte à fixer la sous-unité α du facteur 2 d'initiation de la traduction (eIF2α). PERK ainsi activée peut alors phosphoryler la Ser 51 de eIF2α, ce qui inhibe l'initiation de la synthèse des protéines et réduit la quantité de protéines dépliées qui entre dans le RE. La structure du domaine kinase de PERK (PDB 3QD2) montre que les protomères au sein du dimère sont parallèles et "dos-à-dos" via notamment un pont salin entre Arg 262 d'un protomère et Asp 266 de l'autre. Ces 2 acides aminés et d'autres sont conservés chez toutes les kinases de eIF2α. Exemples sur la figure ci-dessous :
Source : Dey et al. (2007)
b. Le facteur de transcription ATF6 ATF6 ("Cyclic AMP-dependent transcription factor ATF-6" ou "Activating Transcription Factor 6") est un facteur de transcription de 670 acides aminés (75 kDa) de la famille ATF. Il contient un domaine "basic region leucine zipper" (bZIP) N-terminal qui lui permet de se fixer sur les séquences ERSE ou UPRE ("unfolded protein response responsive element"). Ces séquences sont présentes au niveau des promoteurs des gènes cibles spécifiques de l'UPR. Il se fixe sur les séquences ERSE (5'-CCAAT-N(9)-CCAC[GA]-3') et ERSE II (5'-ATTGG-N-CCACG-3'). L'état N-glycosylé (Asn 472, Asn 584 et Asn 643) de ATF6 sert de senseur pour l'homéostasie du RE : ATF6 est activé ou non pour enclencher ou non l'UPR. Au cours de l'UPR, ATF6 est réduit. Le monomère réduit est transporté dans l'appareil de Golgi pour y être protéolysé : un fragment d'environ 50 kDa contenant le domaine cytoplasmique qui possède l'activité facteur de transcription est relargué. Le clivage semble s'opérer de manière séquentielle par S1P et S2P. Le domaine N-terminal qui porte l'activité facteur de transcription ainsi libéré se dimèrise (formation d'homodimère ou d'hétérodimère avec ATF6-β). Ce dimère est délocalisé dans le noyau pour y activer la transcription de gènes de protéines chaperonnes du RE. ATF6 peut être délocalisé dans l'appareil de Golgi en absence de stress du RE mais il retourne vers le RE parce que la forme oxydée de ATF6 est résistante à la protéolyse par S1P. La réponse de phase aigüe (inflammation) Quand des cytokines inflammatoires, telles que TNF-α, IL-1β et IL-6, sont présentes dans l'environnement extracellulaire, le gène codant le facteur de transcription CREBH ("cAMP-Responsive-Element-Binding protein H") est transcrit. CREBH est une protéine semblable à ATF6. CREBH est un facteur de transcription contenant un domaine bZIP localisé sur la membrane du RE. Cependant, CREBH est principalement exprimé par les hépatocytes, tandis que ATF6 est exprimé par tous les types de cellules. En conditions de stress du RE (cytokines inflammatoires ou lipopolysaccharides bactériens), CREBH est délocalisé vers l'appareil de Golgi, où il est protéolysé par les protéases S1P and S2P, qui relarguent un fragment cytosolique. Le stress du RE active aussi ATF6. Les formes activées de CREBH et de ATF6 peuvent alors former des homodimères ou des hétérodimères et migrer vers le noyau, où elles activent la transcription de gènes codant le composant amyloïde P du sérum ("serum amyloid P component") et la protéine C - réactive qui médient la réponse de phase aigüe. |
c. L'enzyme bifonctionnelle IRE-1 ("Inositol-Requiring Enzyme-1") IRE-1 est une protéine transmembranaire de type I (977 acides aminés - 110 kDa chez l'homme) activée par la baisse de concentration de l'inositol.
Via TRAF2 ("TNF Receptor-Associated Factor"), IRE-1 recrute et active la protéine ASK1 ("Apoptosis Signal-regulating Kinase 1" - un médiateur de l'apoptose) qui à son tour active JNK ("c-Jun N-terminal Kinase"). Cela induit la mort cellulaire par apoptose.
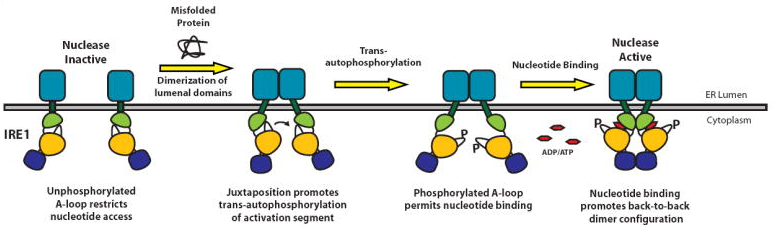
Source : Muoio & Newgard (2004) IRE-1 joue un rôle important dans l’insulino-résistance périphérique. Le stress du RE et l’altération de l’axe de signalisation IRE-1/XBP1 sont liés à une perturbation de la voie de signalisation du récepteur de l’insuline dans le foie. La protéine IRE-1 de Saccharomyces cerevisiae est composée d'un domaine situé dans la lumière du RE et d'un domaine cytosolique. Le domaine de IRE-1 situé dans la lumière du RE contient :
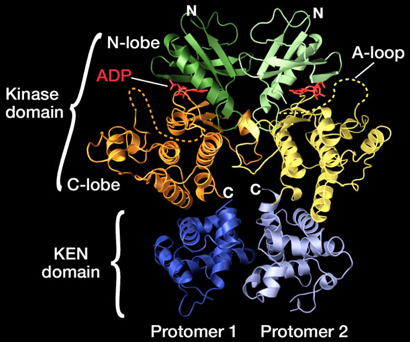
Cette dimérisation induit des changements de conformation qui sont transmis au domaine cytosolique via le segment transmembranaire. La dimérisation des domaines de IRE-1 situés dans la lumière du RE entraîne une juxtaposition des domaines cytosoliques qui induit une autophosphorylation des domaines kinases qui, à son tour, induit l'activation des domaines endoribonucléases. Chaque protomère de IRE-1 est constitué d'un domaine N-terminal kinase (EC 2.7.11.1) et d'un domaine C-terminal ribonucléase (EC 3.1.26 - "Kinase-Extension Nuclease domain" - KEN).
Figure ci-dessus : structure 3D de IRE-1 (PDB 2RIO). Source : PDBJ Le domaine kinase est divisé en un petit lobe N-terminal ("N-lobe") et un gros lobe C-terminal ("C-lobe"). Une molécule d'ADP est fixé à la jonction entre ces lobes. Le site catalytique ribonucléase de IRE-1 est localisé à proximité de 4 acides aminés (Y1049, R1056, N1057, H1061). L'arrangement de ces 4 acides aminés montre des similarités avec le site catalytique de la "tRNA-splicing endonuclease" (2GJW), ce qui indique qu'ils pourraient être impliqués dans l'épissage de l'intron de l'ARN messager codant le facteur de transcription XBP-1. Le mécanisme de dimérisation de IRE-1
Source : Lee et al. (2008)
|
7. La protéine chaperonne BiP/GRP78 La protéine régulée par le glucose ("78 kDa glucose-regulated protein" - GRP78, aussi appelée "immunoglobulin binding protein" - BiP) appartient à la famille des protéines de choc thermique ("Heat Shock Protein" - HSP70), protéines chaperonnes qui aident au repliement d'autres protéines. C'est une protéine de 78 kDa dont la séquence en acides aminés est très conservée de la levure à l'homme. Elle est composée de 3 domaines: le domaine ATPase (qui va fournir l'énergie au repliement de la protéine dépliée), le domaine de fixation du peptide (qui fixe la protéine dépliée) et un domaine C-terminal dont la fonction n'est pas connue (structure 3D : PDB 3IUC).
|
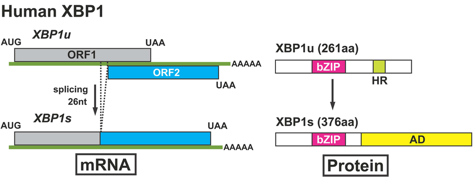
8. Le facteur de transcription XBP1 ("X-box binding protein 1") Chez l'homme, le gène codant XBP1 est localisé sur le chromosome 22. Dans le noyau, IRE-1 catalyse l'excision d'un intron de 26 paires de bases de l'ARN messager codant XBP1, ce qui change le cadre de lecture de la séquence codant cette protéine. Il en résulte la traduction de l'isoforme XBP1s ("s" pour "spliced" - épissé) de 376 acides aminés (54 kDa) à la place de l'isoforme XBP1u de 261 acides aminés (33 kDa) :
Figure ci-dessous : épissage non-conventionnel de l'ARN messager codant XBP1 (homme) et ses conséquences en terme de protéines traduites.
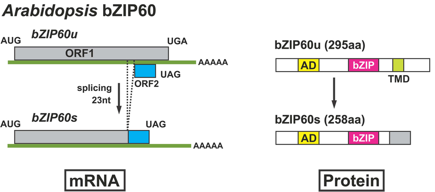
Source : Nagashima et al. (2011) Arabidopsis possède 2 isoformes de IRE-1 fonctionnellement redondantes en ce qui concerne l'épissage de l'ARN messager de bZIP60. Figure ci-dessous : épissage non-conventionnel de l'ARN messager codant bZIP60 (Arabidopsis) et ses conséquences en terme de protéines traduites.
Source : Nagashima et al. (2011) Les 2 isoformes XBP1s et XBP1u forment un hétérodimère qui est exporté vers le cytoplasme pour y être dégradé par le sytème protéasome. Ce mécanisme diminue la quantité d'isoforme XBP1s qu'il est nécessaire de dégrader en absence de stress du RE. XBP1 est un facteur de transcription (de la famille ATF/CREB) nécessaire à la transcription d'un sous-ensemble de gènes des complexes majeurs d'histocompatibilité de classe II ("class II MHC genes") chez l'homme. XBP1 joue aussi un rôle prépondérant dans la synthèse d’immunoglobulines en induisant la différenciation des cellules plasmocytaires. Il contient un domaine "basic region leucine zipper" (bZIP) N-terminal qui lui permet de se fixer sur les séquences ERSE ou UPRE ("unfolded protein response responsive element"). L’épissage, par IRE-1, de l'ARN messager codant XBP1 est un événement transitoire qui disparaît au bout de 16h environ lors d’un stress du RE. L’inhibition de l’épissage de l'ARN messager codant XBP1 ou l’inactivation de XBP1 induit une diminution de la synthèse hépatique de lipides d'où une diminution des triglycérides, des acides gras libres et du cholestérol dans le sérum. La forme active de XBP1 est SUMOylée principalement par PIAS2 sur 2 Lys localisées à l'extrémité C-terminale du domaine de transactivation. La sumoylation est une modification post-traductionnelle qui fixe de manière covalente des protéines "SUMO" ("Small Ubiquitin-like MOdifier"). La sumoylation est proche, dans son principe, de l'ubiquitinylation mais les protéines sumoylées ne sont pas destinées à la dégradation.
|
9. La voie de dégradation ERAD ("ER Associated Degradation") Les protéines mal repliées sortent du RE par le canal de translocation. Elles sont ubiquitinylées et enfin dégradées par les protéasomes dans le cytoplasme. La moitié des protéines qui transitent par le RE sont détruites par cette voie.
La mannosidase I du RE hydrolyse le mannose i (Man8-GlcNAc2). Les protéines mal repliées sont alors reconnues par EDEM ("ER-Degradation Enhancing α-Mannosidase-like protein"), une lectine liée à CNX par son domaine transmembranaire.
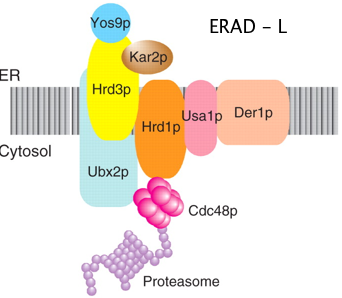
EDEM hydrolyse le résidu mannose k de l'oligosaccharide (Man7-GlcNAc2). Cette coupure génère un signal qui recrute Yos9p, une lectine ERAD qui contient un domaine homologue au récepteur mannose 6-phosphate qui fixe les oligosaccharides Man7. Le mécanisme ERAD-L vérifie l'état de repliement des domaines situés dans la lumière du RE des protéines membranaires. Il vérifie aussi les protéines solubles puisqu'elles sont entièrement dans la lumière du RE.
Source : Hoseki et al. (2010) Le mécanisme ERAD-M vérifie l'état de repliement des protéines transmembranaires. Si un "défaut" de repliement y est détecté, la protéine mal repliée est éliminée.
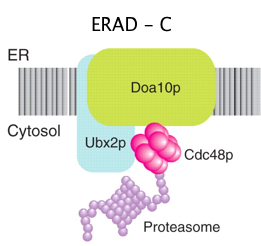
Le mécanisme ERAD-C vérifie l'état de repliement des domaines cytosoliques des protéines membranaires. Si un "défaut" de repliement y est détecté, la protéine mal repliée est éliminée.
Source : Hoseki et al. (2010) Légende : Doa10p : ubiquitine ligase E3 - Cdc48 : "chaperone-like AAA ATPase" - AAA ou AAA+ : "ATPases Associated with diverse cellular Activities" |
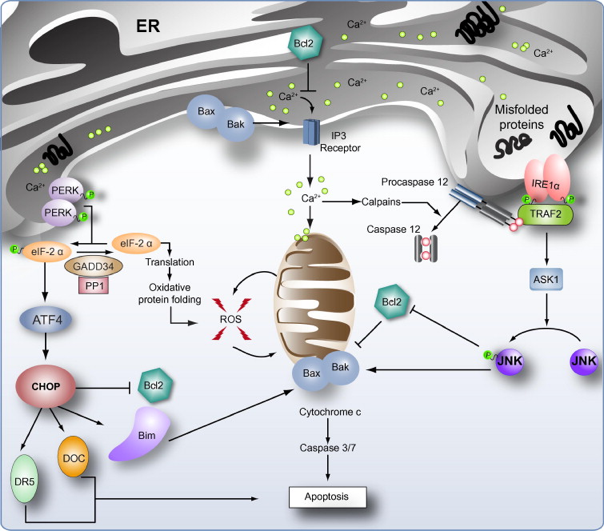
10. Aperçu de l'apoptose résultant de l'UPR La réponse UPR impliquant IRE-1 est cytoprotective et permet à la cellule de s’adapter à un stress lié à la toxicité des protéines mal repliées. Si ce stress ne peut être endigué, la cellule entre alors en apoptose. Si l’épissage de XBP1 (et donc l’activation de la voie IRE-1/XBP1) est maintenu plus de 48h, le nombre de cellules apoptotiques diminue. Les protéines pro-apoptotiques Bax et Bak sont localisées sur la membrane du RE et interagissent avec le domaine cytosolique d'IRE-1. Elles régulent l'homéostasie calcique.
Source : Malhi & Kaufman (2011) Le relarguage du calcium du RE peut activer des calpaïnes, qui peuvent à leur tour activer par protéolyse la caspase 12 qui médie l'apoptose.
CHOP ("C/EBP HomolOgous Protein") induit l'expression de GADD34 qui s'associe avec la protéine phosphatase 1 (PP1) pour déphosphoryler eIF2α (rétro-contrôle négatif). CHOP peut :
La protéine adaptatrice TRAF2 ("TNF Receptor-Associated Factor"), s’associe au domaine kinase de IRE-1. Ce complexe interagit alors avec ASK1 ("Apoptosis Signal-regulating Kinase 1") pour activer la JNK ("c-Jun N-terminal Kinase") qui a des effets pro-apoptotiques : activation induite par phosphorylation de Bim et inactivation des protéines Bcl2. Bien que le pourcentage d'identité entre les séquences des protéines de la famille Bcl-2 soit faible, elles possèdent toutes un à quatre domaines conservés appelés domaine BH ("Bcl-2 homology domain"). Les protéines pro-apoptotiques sont sub-divisées en 2 groupes :
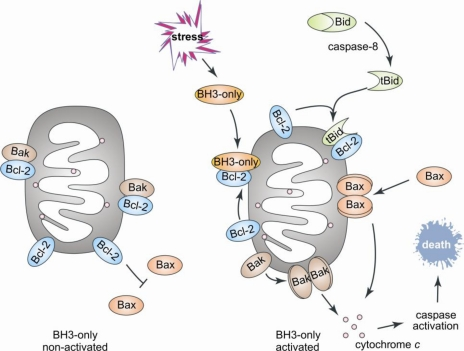
Le schéma ci-dessous décrit le rôle des protéines de la famille Bcl-2 dans l'apoptose mitochondriale "intrinsèque" ou activée.
Source : Rautureau et al. (2010) La protéine "BH3-only" Bid établit le lien entre l'apoptose dont la signalisation est sous le contrôle du récepteur de la mort cellulaire ("extrinsèque") et l'apoptose mitochondriale "intrinsèque". Bid est activée par protéolyse par la caspase-8 au niveau de sa région intrinsèquement désordonnées ("Intrinsically disordered regions"). |
|
Figure ci-dessous : une magnifique synthèse de l'UPR.
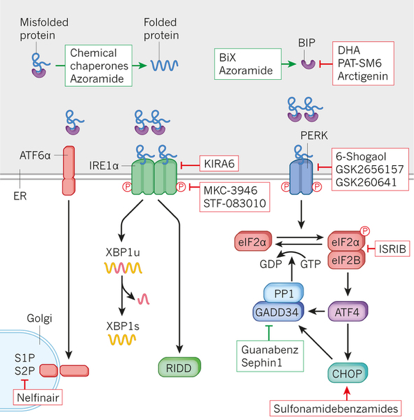
Source : Derrick et al. (2008) Beaucoup de petites molécules atténuent (encadrés verts - figure ci-dessous) ou induisent (encadrés rouges) le stress du RE par différents mécanismes.
Source : Wang & Kaufman (2016) Les chaperons chimiques sont des petites molécules qui empêchent l'agrégation des protéines, qui facilitent leur repliement et qui diminuent le stress du RE (à la fois in vitro et in vivo), en stabilisant les structures intermédiaires du repliement des protéines. Les chaperons chimiques tels que l'azoramide (composé anti-diabète) et la molécule "BiP inducteur X" (BiX) atténuent le stress du RE en renforçant la capacité de repliement du RE. Les molécules Guanabenz et Sephin1 inhibent GADD34 ("DNA-damage-inducible protein 34") ce qui induit une phosphorylation persistante de eIF2α : il en résulte une diminution de la traduction des ARN messagers et du stress du RE. Les petites molécules qui bloquent l'activation de l'UPR incluent la molécule "Integrated Stress Response Inhibitor" (IRSIB) qui cible :
CHOP : "CCAAT/enhancer-binding protein (C/EBP) homologous protein" |
| 11. Liens Internet et références bibliographiques |
|
Derrick et al. (2008) "The endoplasmic reticulum stress response in immunity ans autoimmunity" Nature Rev. Immunol. 8, 663 - 674 Iwawaki T. (2009) "Learning more about physiological endoplasmic reticulum stress using the ERAI system" RIKEN Advanced Science Institute Lynes & Simmen (2011) "Urban planning of the endoplasmic reticulum (ER): How diverse mechanisms segregate the many functions of the ER" BBA 1813, 1893 - 1905 |
|
|
Liu & Kaufman (2003) "The unfolded protein response" J. Cell Sci. 116, 1861 - 1862 Foufelle & Ferré (2007) "La réponse UPR - Son rôle physiologique et physiopathologique" Med. Sci. 23, 291 - 296 Zhang & Kaufman (2008) "From endoplasmic-reticulum stress to the inflammatory response" Nature 454, 455 - 462 Bouchecareilh & Chevet (2009) "Stress du réticulum endoplasmique - Une réponse pour éviter le pIRE" Med. Sci. 25, 281 - 287 |
|
|
Carte interactive de l'UPR - KEGG ("Kyoto Encyclopedia of Genes and Genomes") Visualisation du réticulum endoplasmique en 3D Vidéo du mécanisme : "Signal-Recognition-Particle". "Ulysse" : ensemble de cours de l'Université de Bordeaux |
|
|
Rapoport T.A. (2007) "Protein translocation across the eukaryotic endoplasmic reticulum and bacterial plasma membranes" Nature 450, 663 - 669 Kusters & Driessen (2011) "SecA, a remarkable nanomachine" Cell. Mol. Life Sci. 68, 2053 - 2066 Ali et al. (2011) "Structure of the Ire1 autophosphorylation complex and implications for the unfolded protein response" The EMBO J. 30, 894 - 905 Dey et al. (2007) "Conserved Intermolecular Salt Bridge Required for Activation of Protein Kinases PKR, GCN2, and PERK" J. Biol. Chem. 282, 6653 - 6660 |
|
|
Muoio & Newgard (2004) "Biomedicine. Insulin resistance takes a trip through the ER" Science 306: 425 - 426 Nadanaka et al. (2007) "Role of disulfide bridges formed in the luminal domain of ATF6 in sensing endoplasmic reticulum stress" Mol. Cell Biol. 27, 1027 - 1043 Lee et al. (2008) "Structure of the dual enzyme Ire1 reveals the basis for catalysis and regulation in non-conventional RNA splicing" Cell 132, 89 - 100 Nagashima et al. (2011) "Arabidopsis IRE1 catalyses unconventional splicing of bZIP60 mRNA to produce the active transcription factor" Sci. Rep. 1, 29 |
|
|
Hoseki et al. (2010) "Mechanism and components of endoplasmic reticulum-associated degradation" J. Biochem. 147, 19 - 25 Rautureau et al. (2010) "Intrinsically Disordered Proteins in Bcl-2 Regulated Apoptosis" Int. J. Mol. Sci. 11, 1808 - 1824 Bernasconi & Molinari (2011) "ERAD and ERAD tuning: disposal of cargo and of ERAD regulators from the mammalian ER" Curr. Opin. Cell Biol. 23, 176 - 183 Malhi & Kaufman (2011) "Endoplasmic reticulum stress in liver disease" J. Hepatol. 54, 795 - 809 |
|
|
Halic & Beckmann (2005) "The signal recognition particle and its interactions during protein targeting" Curr. Opin. Struc. Biol. 14, 116 - 125 Andrès et al. (2010) "The TOC complex: Preprotein gateway to the chloroplast" BBA 1803, 715 - 723 Träger et al. (2012) "Evolution from the Prokaryotic to the Higher Plant Chloroplast Signal Recognition Particle: The Signal Recognition Particle RNA Is Conserved in Plastids of a Wide Range of Photosynthetic Organisms" Plant Cell 24, 4819 - 4836 Akopian et al. (2013) "Signal Recognition Particle: An Essential Protein-Targeting Machine" Ann. Rev. Biochem. 82 Wang & Kaufman (2016) "Protein misfolding in the endoplasmic reticulum as a conduit to human disease" Nature 529, 326–335 |
![]()