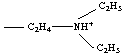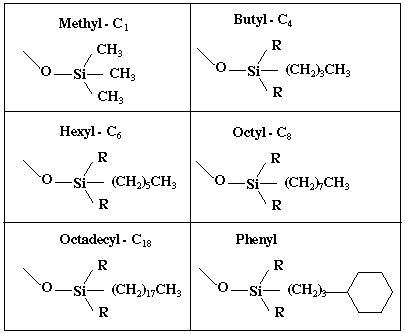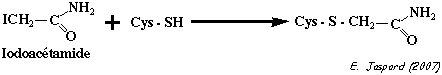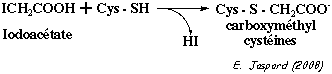| Purification et caractérisation de la sapécine |
| Tweet |
|
|
1. Enoncé 2. Réponse partie A 3. Réponse partie B |
4. Réponse partie C 5. Complément d'information sur la sapécine 6. Références bibliographiques |
|
1. Enoncé Certains diptères synthètisent des polypeptides qui possèdent une fonction double. L'un de ces polypeptides, la sapécine, synthètisé par la mouche Sarcophaga peregrina, est un facteur de croissance des cellules embryonnaires de la mouche et également un antibactérien. Sur la base de ses propriétés antibactériennes, c'est-à-dire sa capacité à inhiber la croissance d'une culture bactérienne, la sapécine est purifiée à partir de cellules embryonnaires en culture. Partie A : à partir de 900 mL de milieu de culture, les étapes de purification donnent les résultats présentés dans le tableau ci-dessous. |
| Etapes | Activité Totale (unités arbitraires) | quantité de protéines (mg) |
| milieu de culture | 6675 | 3258 |
| chromatographie sur carboxyméthylcellulose | 4247 | 26,8 |
| concentration | 2912 | 15,8 |
| filtration sur gel | 1986 | 5,3 |
| chromatographie sur phase reverse | 1540 | 0,53 |
|
a. Rappelez le principe des techniques utilisées à chaque étape. Comment l'activité totale est-elle mesurée ? Partie B : une quantité inconnue du polypeptide purifié est prélevée dans le but d'en déterminer la composition en acides aminés. a. Rappeler le principe de la technique. Partie C : afin de déterminer la
présence éventuelle de ponts disulfures, on fait réagir le polypeptide avec un mélange d'iodoacétamide et d'acide iodoacétique (iodoacétate), tous deux marqués au carbone 14.
|
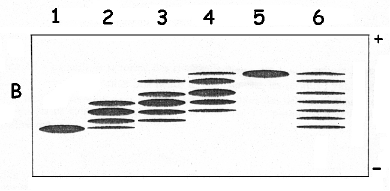
| Piste 1 | iodoacétamide radioactif 100% |
| Piste 2 | iodoacétamide radioactif 66% - iodoacétate radioactif 33% |
| Piste 3 | iodoacétamide radioactif 50% - iodoacétate radioactif 50% |
| Piste 4 | iodoacétamide radioactif 33% - iodoacétate radioactif 66% |
| Piste 5 | iodoacétate radioactif 100% |
| Piste 6 | mélange de tous les dépôts des pistes 1 à 5 |
|
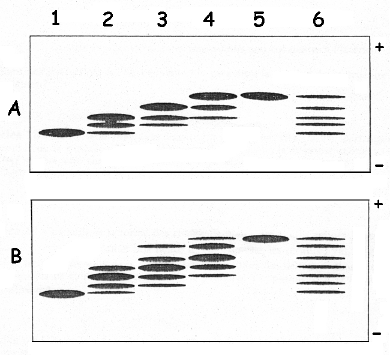
Le polypeptide natif et non traité par les réactifs radioactifs migre sous la forme d'une bande au même niveau que l'échantillon de la piste 1 du gel A ou du gel B (figure ci-dessus). a. Expliquer l'apparition des
bandes radioactives observées sur l'autoradiogramme.
|
| phase stationnaire | principe de séparation | catatéristiques de la phase stationnaire | principe de la fixation et de l'élution |
| liquide | partage | liquide fixé sur un support inerte (papier, silice...) | distribution des composants du mélange à séparer dans les deux phases liquides selon leur coefficient de partage |
| solide | adsorption | adsorbant solide polaire | phénomène de surface : formation de liaisons spécifiques entre les composants et la surface adsorbante |
|
adsorption
(phase inverse) |
molécules hydrophobes greffées sur de la silice | interactions hydrophobes et élution par diminution de la polarité de la phase mobile | |
| échange d'ions | résine (polymères d'oses) porteuse de groupements chargés négativement ou positivement | interactions électrostatiques avec les composants de charge opposée | |
|
exclusion
(filtration sur gel) |
solide poreux | les composants de diamètre supérieur à celui des billes du support sont "exclus" et ceux de diamètre inférieur y diffusent et sont freinés | |
| affinité | support sur lequel est greffée une molécule (le ligand) spécifiquement reconnue par un des composants de l'échantillon à analyser | déplacement de l'équilibre de liaison [molécule - ligand greffé] en faveur de l'équilibre [molécule - tierce molécule] |
|
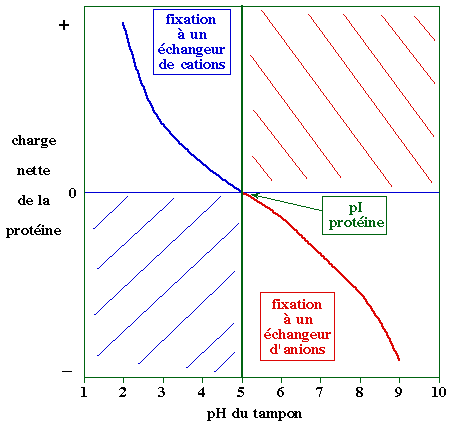
Les différents types d'échangeurs d'ions Les échangeurs d'ions sont des billes qui portent des groupements ionisables dont la charge est : |
| positive : résines échangeuses d'anions qui fixent des molécules chargées négativement -R+...M- | négative : résines échangeuses de cations qui fixent des molécules chargées positivement -R-...M+ | ||
| nature de la fonction ionisable : -R+ | nature de la fonction ionisable : -R- | ||
| base forte | base faible | acide fort | acide faible |
|
ammonium quaternaire : -NR3+ exemple : triméthylammonium -N(CH3)3+ |
forme protonnée d'une amine I, II ou III : -NHR2+ exemple : diéthylaminoéthylammonium
|
exemple : sulfonate -SO3- |
exemple : carboxyméthyl -O-CH2-CO2- |
Dans un premier temps, la résine est équilibrée dans un tampon dont le pH est tel que le groupement porté par l'échangeur d'ions soit ionisé :
Les molécules à séparer sont dans le même tampon (donc au même pH) et en fonction de leur point isoélectrique (pI), elle porte une charge nette :
Il y a deux manières d'éluer les molécules fixées sur la résine :
|
|
Chromatographie d'exclusion ou filtration sur gel Le principe de la chromatographie d'exclusion (appelée aussi filtration sur gel ou tamisage moléculaire ou chromatographie de perméation) repose en première approximation sur la masse molaire des molécules à séparer. En toute rigueur, le paramètre physico-chimique qui intervient est la dimension des molécules, donc leur volume et donc plus précisément leur rayon de Stokes. Mais cette relation univoque [masse molaire - rayon de Stokes] n'est valable que pour des protéines dites "globulaires", c'est-à-dire assimilées à des sphères. Différents paramètres influencent le volume des molécules et peuvent fausser la détermination de la masse molaire réelle par ce type de chromatographie :
La phase stationnaire est constituée de billes de polysaccharides (type "Sephadex") gonflées dans le solvant utilisé pour l'élution :
|
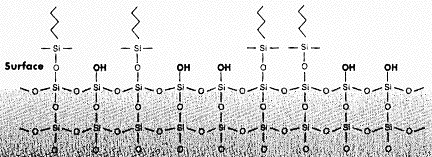
Chromatographie à polarité de phase inversée ou en phase reverse Les matrices pour chromatographie en phase reverse sont souvent à base de silice (SiO2), appelée aussi gel de silice car elle est plus ou moins hyratée. La silice provient de la déshydratation de l'acide silicique et se présente sous forme d'une poudre dont les grains (les billes) sont très fins.
On utilise fréquemment une silice sur laquelle des chaînes alkylées (voir les différents groupes dans le tableau ci-dessous) sont greffées au niveau des groupements silanols.
Les chaînes alkylées confèrent à la silice un caractère très hydrophobe :
|
|
La phase mobile Elle est donc constituée d'un mélange eau / solvant organique, en proportion variable. La phase aqueuse : l'eau est le solvant le plus polaire (le moins hydrophobe). On ajoute un sel pour tamponner le milieu. Cependant, on emploie souvent un acide fort (l'acide trifluoroacétique) et une amine quaternaire (la triéthylamine, par exemple) car ces deux composés ioniques forment des paires d'ions avec les molécules à séparer, neutralisant ainsi des charges portées par ces dernières. Celà a pour conséquence d'augmenter l'hydrophobicité des molécules et ainsi qu'elles s'adsorbent davantage sur la phase stationnaire. Le solvant organique :
Voir un cours sur les différentes techniques de chromatographie. |
|
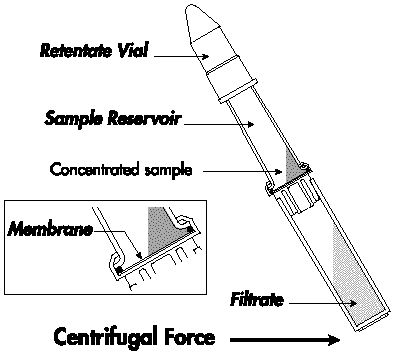
La concentration des protéines L'ultrafiltration : l'échantillon est mis dans un tube. Ce tube contient une membrane semi-perméable dont les pores ont un diamètre tel que seules l'eau et des molécules au-dessous d'une certaine masse molaire choisie ("cut-off") peuvent la franchir. Le tube est placé dans une centrifugeuse et, sous l'action de la force centrifuge, l'eau et les petites molécules traversent la membrane. Une autre méthode de concentration est la précipitation par un sel (exemple : le sulfate d'ammonium). La précipitation par un sel nécessite de dialyser l'échantillon après concentration. Ces techniques de concentration occasionnent souvent des pertes importantes de matériel.
Source : Millipore |
|
L'activité totale (AT) correspond à l'activité antibactérienne de la sapécine : 1 unité d'activité antibactérienne est la quantité de polypeptide qui entraine 50% d'inhibition de la croissance bactérienne (Escherichia coli) par rapport à un contrôle. |
|
AT
étape n R (%) = -------------- AT étape 1 |
AT
étape n AS (UA/mg) = -------------- Q étape n |
AS
étape n FP = -------------- AS étape 1 |
| Etapes | AT (unités arbitraires) | Q protéines (mg) | R (%) | AS (UA/mg) | FP |
| milieu de culture | 6675 | 3258 | 100 | 2 | 1 |
| chromatographie sur carboxyméthylcellulose | 4247 | 26,8 | 63,6 | 158 | 79 |
| concentration | 2912 | 15,8 | 43,6 | 184 | 92 |
| filtration sur gel | 1986 | 5,3 | 29,8 | 375 | 187,5 |
| chromatographie sur phase reverse | 1540 | 0,53 | 23,1 | 2906 | 1453 |
| R : rendement - AT : activité totale - Q : quantité de protéines - AS : activité spécifique - FP : facteur de purification | |||||
|
Le polypeptide peut être considéré comme "pur" car une forte activité totale (AT) est conservée pour une quantité minime de protéines en regard des valeurs initiales. Bien évidemment, à l'échelle moléculaire et compte-tenu du nombre d'Avogadro, aucun corps (hormis sous forme de cristal "pur") ne peut être considéré comme totalement pur. A fortiori en ce qui concerne les protéines extraites de milieux extrêmement composites et complexes comme la cellule. Cependant, l'enrichissement est tel (facteur de purification de 1453) que, macroscopiquement, on peut considérer que la solution ne contient "quasi exclusivement" que le polypetide purifié. |
|
Conditions d'hydrolyse totale acide des liaisons peptidiques pour la détermination de la composition minimale en acides aminés : réduction des ponts disulfure / HCl 6N / 110°C pendant 24 à 72 h.
Les acides aminés sont ensuite :
|
|
La composition en acides aminés est rapide (15 minutes) et nécessite environ 1 pmole de chaque acide aminé. On obtient une estimation du pourcentage de chaque acide aminé dans la chaîne polypeptidique. Remarque : cette technique est de moins en moins utilisée car il n'est pas nécessaire de déterminer la composition en acides aminés pour déterminer la séquence. En effet, de plus en plus de séquences peptidiques sont :
Les valeurs obtenues après hydrolyse acide sont multiples de 7,5 [Leu 45 / Thr 7,5 / Cys 45 / Glu 30 /Ile 7,5 / Asx 7,5 / Arg 30 / Ser 15 / His 7,5 / Lys 15 / Gly 7,5 / Tyr 15 / Ala 7,5 / Val 15].
|
|
L'iodoacétamide et l'iodoacétate sont des réactifs des groupements thiols (-SH) :
Il est donc capital de séparer les dérivés par électrophorèse en conditions natives afin que les différentes formes du polypeptide migrent seulement en fonction de leurs charges nettes respectives. Plus le polypeptide est chargé négativement, moins il migre : c'est le traitement au iodoacétate radioactif 100% => piste 5 des gels A et B. |
|
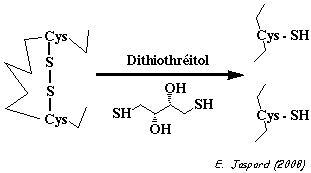
Le dithiothréitol (DTT - aussi appelé réactif de Wallace Cleland) réduit les ponts disulfures. Toutes les Cys sont accessibles aux réactifs.
Le dithioérythritol (épimère du DTT) est aussi un agent réducteur, mais moins efficace que le DTT. Enfin, l'iodoacétamide et l'iodoacétate étant radioactifs, les Cys qui réagissent sont elles aussi marquées au 14C. Lors de l'autoradiographie, la radioactivité impressionne le film aux endroits où migrent les différentes formes du polypeptide. |
|
Gel A : le polypeptide purifié est non réduit. Seules les Cys libres sont marquées par l'un et l'autre des deux réactifs.
|
Gel B : le polypeptide purifié est d'abord traité par le dithiothréitol. Toutes les Cys sont marquées.
|
Interprétation de la figure du gel natif A
Mais on ne peut pas visualiser de bandes car ces polypeptides sont à l'état de trace*. |
| Traitement | piste | polypeptide natif - gel A | polypeptide réduit - gel B |
|
iodoacétamide radioactif 100% |
1 |
1 bande qui migre le plus loin : le polypeptide est non chargé 1 forme => toutes les Cys sont sous la forme Cys-CO-NH2 La bande piste 1 - gel A migre aussi loin que la bande piste 1 - gel B. |
|
|
iodoacétamide radioactif 66% iodoacétate radioactif 33% |
2 |
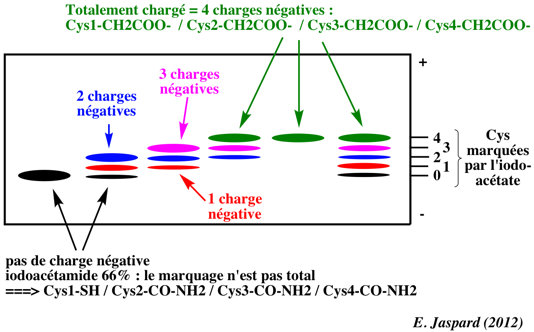
3 bandes = 3 formes variables du polypeptide selon le pourcentage respectif iodoacétamide - iodoacétate. Par exemple, pour 66% - 33 %, co-existence des formes principales* : Cys1-SH / Cys2-CO-NH2 / Cys3-CO-NH2 / Cys4-CO-NH2 : pas de charge, forme qui migre le plus loin (identique à la piste 1) Cys1-SH / Cys2-CO-NH2 / Cys3-CO-NH2 / Cys4-CH2COO- : 1 charge négative Cys1-SH / Cys2-CO-NH2 / Cys3-CH2COO- / Cys4-CH2COO- : 2 charges négatives |
4 bandes = 4 formes du polypeptide => 0 à 3 Cys marquées par l'iodoacétate |
|
iodoacétamide radioactif 50% iodoacétate radioactif 50% |
3 |
5 bandes = 5 formes du polypeptide => 0 à 4 Cys marquées par l'iodoacétate |
|
|
iodoacétamide radioactif 33% iodoacétate radioactif 66% |
4 | ||
|
iodoacétate radioactif 100% |
5 |
1 bande qui migre le moins loin : le polypeptide est totalement chargé négativement 1 forme => toutes les Cys sont sous la forme Cys-CH2COO- La bande piste 5 - gel A migre plus loin que la bande piste 5 - gel B donc le polypeptide natif est moins négatif que le polypeptide réduit. |
|
|
mélange : |
6 |
5 bandes = 5 formes du polypeptide : Cys1-CO-NH2 / Cys2-CO-NH2 / Cys3-CO-NH2 / Cys4-CO-NH2 |
7 bandes = 7 formes du polypeptide => 0 à 6 Cys marquées par l'iodoacétate |
|
Conclusions 1. Le polypeptide natif et non traité par les réactifs radioactifs migre exactement comme la bande 1 du gel A ou du gel B : la sapécine est un monomère. 2. Une bande apparaît dans la piste 1 du gel B et sept bandes apparaissent dans la piste 6 du gel B : la sapécine contient 6 cystéines. 3. Deux bandes additionnelles apparaissent dans la piste 6 du gel B par rapport à la piste 6 du gel A. La réduction par le DTT révèle 2 Cys supplémentaires : il y a 1 pont disulfure. Voir un cours sur les cystéines et la formation des ponts disulfure. |
|
5. Complément d'information sur la sapécine
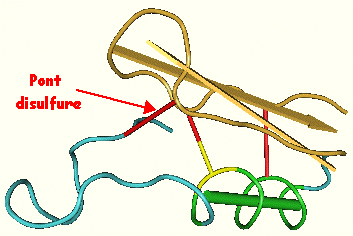
La sapécine mature contient 3 ponts disulfures (figure ci-contre) : Cys 3 - Cys 30 / Cys 16 - Cys 36 / Cys 20 - Cys 38 (numérotation par rapport à la forme mature - voir ci-dessous).
Sapécine de Sarcophaga peregrina - Hanzawa et al. (2002) - Code d'accès : MMDB : 19110 / PDB : 1L4V L'énoncé présente donc des profils électrophorètiques qui ne reflètent pas la réalité, et ce, dans un but pédagogique afin de mieux comprendre la démarche pour déterminer le nombre de cystéines et de ponts disulfures dans une protéine. Quels seraient les profils électrophorètiques avec 6 cystéines impliquées dans 3 ponts disulfures ? |
|
Visualisation de la sapecine de Sarcophaga peregrina - RMN en solution. Code PDB : 1L4V
|
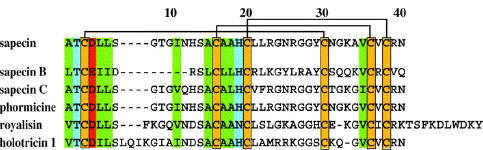
| La sapécine
appartient à la famille des défensines des insectes. Elle réprime la croissance des bactéries Gram+. La sapécine (P18313)
est synthétisée sous forme d'un précurseur qui contient un peptide signal (acides aminés 1 à 23) et un propeptide (acides aminés 24 à 54).
La forme mature de la sapécine correspond donc aux acides aminés 55 à 94, soit 40 acides aminés et une masse molaire de 4080 Daltons. Séquence de la sapécine mature : A1TC3DLLSGTGINHSAC16AAHC20LLRGNRGGYC30NGKAVC36VC38RN40
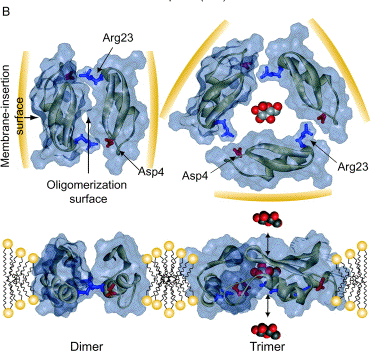
Source : Takeuch et al. (2004) C'est une protéine basique avec une charge nette cationique de +3 au pH physiologique. Des interactions électrostatiques s'établissent entre les acides aminés basiques et un phospholipide acide, la cardiolipine. Ces interactions entraînent la formation de pores puis la perméabilisation de la membrane des bactéries. Lors de cette interaction, la sapécine adopterait une structure oligomèrique.
Source : Takeuch et al. (2004) |
| 6. Références bibliographiques |
|
Matsuyama & Natori (1988) "Purification of three antibacterial proteins from the culture medium of NIH-Sape-4, an embryonic cell line of Sarcophaga peregrina" J. Biol. Chem. 263, 17112 - 17116 Kuzuhara et al. (1990) "Determination of the Disulfide Array in Sapecin, an Antibacterial Peptide of Sarcophaga peregrina (Flesh Fly)" J. Biol. Chem. 107, 514 - 518 Takeuch et al. (2004) "Channel-forming membrane permeabilization by an antibacterial protein, sapecin - Determination of membrane-buried and oligomerization surfaces by NMR" J. Biol. Chem. 279, 4981 - 4987 |
Synthèse de cette correction au format PDF (tous droits réservés biochimej). |
![]()