| Les membranes : structure des lipides membranaires et des protéines membranaires |
| Tweet |
|
|
1. Introduction 2. Structure et thermodynamique des membranes
3. Les lipides membranaires 4. Les (glycéro)phosphatidylinositols et les phosphoinositides
5. Relation entre lipides membranaires et protéines transmembranaires
|
6. Environnement biophysique et caractéristiques des protéines transmembranaires
7. Les protéines membranaires et transmembranaires
8. Les aspartyl protéases intramembranaires 9. Un cas très particulier : le site actif de la diacylglycérol kinase bactérienne 10. Les protéines MAPEG 11. Les récepteurs transmembranaires 12. Liens Internet et références bibliographiques |
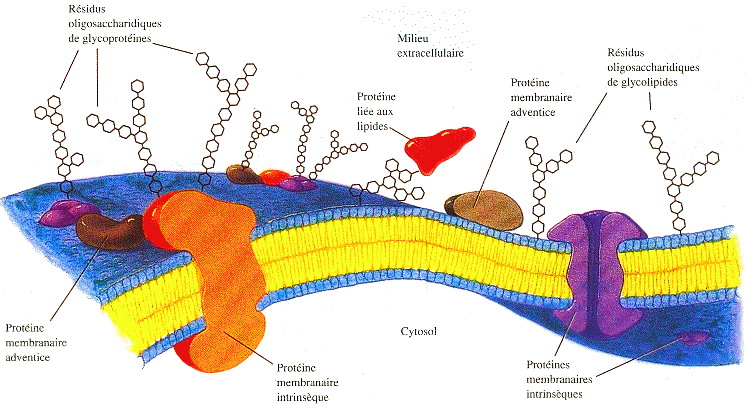
1. Introduction On conçoit difficilement la vie, donc l'entité cellule, sans que l'ensemble des molécules biologiques soient rassemblées et puissent, dés lors, interagir. Le premier rôle des membranes biologiques est donc évident : comme les murs d'une maison, les membranes créent un milieu intérieur (intracellulaire) qui, de facto, est séparé d'un milieu extérieur (extracellulaire). Mais ces murs possèdent des "fenêtres" et des "portes". Le second rôle des membranes est donc de laisser entrer et/ou sortir de manière hautement sélective les molécules biologiques par différents mécanismes de transport. Qu'elles appartiennent à un organisme uni- ou pluri-cellulaires, les cellules ne vivent pas dans un monde isolé : elles doivent donc s'adapter à leur environnement. Le troisième rôle des membranes est de percevoir les caractéristiques du milieu extérieur et transmettre cette information à l'ensemble de la cellule, via les récepteurs et autres protéines transmembranaires. Les cellules ainsi constituées ne sont pas isolées au sein d'un organisme. Le quatrième rôle des membranes est d'établir des relations (communications / échanges) avec d'autres cellules de même type ou autre. En particulier, certains phospholipides modifient les caractéristiques chimiques des membranes et marquent ainsi une cellule afin qu'elle soit reconnue par le système immunitaire pour être détruite. Les membranes biologiques sont un assemblage de protéines, de (phospho)lipides et d'oses en une structure dite en "double feuillet".
Source : "Principes de Biochimie" Horton et al. (1994) Certaines membranes cellulaires sont asymétriques : les deux feuillets de la bicouche lipidique ont une composition différente en phospholipides. Cette différence est maintenue par des transports unidirectionnels ou bidirectionnels de lipides entre les 2 feuillets. Cette asymétrie est primordiale pour de nombreuses fonctions cellulaires (par exemple, la stabilité mécanique de la membrane, la modulation de l'activité de protéines membranaires, la génération de vésicules de sécrétion et l'apoptose). |
|
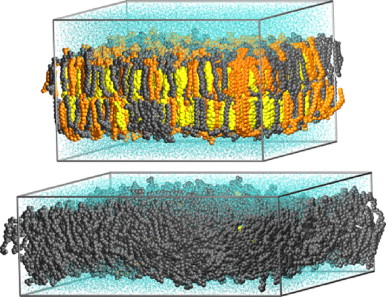
2. Structure et thermodynamique des membranes 1915 : découverte que les membranes sont composées de lipides et de protéines. 1935 : premier modèle de membrane de Danielli et Davson selon lequel une membrane est composée d'une bicouche lipidique avec un "revêtement" hydrophile de protéines des deux côtés. Ce modèle a été rejeté dans les années 1960 par l'emploi de microscopes électroniques. "Davson–Danielli tri-layer (protein–lipid–protein) model" : Danielli & Davson (1935) "A contribution to the theory of permeability of thin films" J. Cell. Comp. Physiol. 5, 495 - 508 1959 : "Unit Membrane Model" Robertson (1959) "The ultrastructure of cell membranes and their derivatives" Biochem. Soc. Symp. 16, 3 - 43 1972 : S. J. Singer et G. L. Nicholson ont proposé que les protéines sont enchâssées dans la bicouche avec leurs extrémités hydrophyles exposées au milieu aqueux externe et leurs régions hydrophobes intégrées dans la membrane. Ce modèle porte le nom de "mosaïque fluide". a. Fluidité des membranes - phases thermodynamiques Les membranes sont des entités dynamiques : leur fluidité (mouvements ondulatoires - voir une vidéo) leur assure une grande souplesse structurale et une grande résistance mécanique. Elles peuvent ainsi s'adapter à des changements des caractéristiques physico-chimiques de leur environnement. La fluidité est le principal déterminant des autres propriétés de la membrane comme la perméabilité des petites molécules. Les phospholipides des membranes se déplacent dans le plan de la membrane avec une vitesse de diffusion latérale très élevée (coefficient de diffusion D ∼ 10-8 cm2.s-1). La fluidité des bicouches lipidiques est en grande partie dictée par la force des interactions de Van der Waals entre les lipides adjacents. Les lipides dont la chaîne carbonée est longue ont plus de surface d'interaction (ce qui augmente la force de cette interaction) et ont donc une mobilité moindre : à une température donnée, un lipide à chaîne carbonée courte sera donc plus mobile (fluide). Tous les lipides ont une température caractéristique à laquelle ils subissent une transition : phase gel <=> phase liquide. En phase liquide, un lipide donné peut échanger sa position avec ses voisins des millions fois par seconde : cet échange aléatoire permet aux lipides de diffuser sur toute la surface de la membrane. En conséquence, l'un des paramètres capitaux pour décrire les propriétés d'une bicouche lipidique est sa phase thermodynamique, qui peut varier de gel ordonné à basse température à liquide désordonné à haute température. Figure ci-dessous, exemples de 2 phases thermodynamiques des bicouches lipidiques : phase liquide ordonné (en haut) et phase liquide désordonné (en bas). Ces images ont été obtenues par simulation par dynamique moléculaire d'une bicouche lipidique "simplifiée" avec 3 composants : la sphingomyéline (en orange), la phosphatidylcholine (en gris) et le cholestérol (en jaune).
Source : Niemela et al. (2009) Par ailleurs, la phase thermodynamique d'une bicouche lipidique dépend de sa composition en lipides :
|
![]()
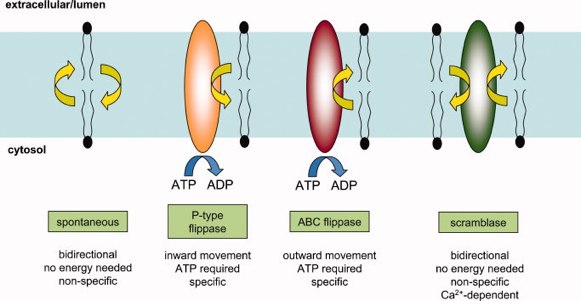
b. Asymétrie des feuillets membranaires et transport des lipides d'un feuillet à l'autre - flippases et scramblases Le mouvement des têtes polaires/chargées des phospholipides à l'intérieur fortement hydrophobe d'une bicouche lipidique est très défavorable du point de vue énergétique. Ainsi, la vitesse de diffusion transversale entre les 2 feuillets est très faible (∼ 10-15 s-1), avec un temps de demi-phénomène de l'ordre de plusieurs heures à plusieurs semaines pour les phospholipides et les glycolipides. Les membranes cellulaires sont asymétriques : les deux feuillets d'une bicouche lipidique d'une membrane ont une composition différente en phospholipides. Cette différence est maintenue par des transports de lipides entre les 2 feuillets.
Source : Adam Steinberg Figure ci-dessous : différents mécanismes de mouvements de lipides membranaires entre les 2 feuillets.
Source : Sharom (2011)
|
![]()
α. Les flippases Les flippases catalysent le transfert net de phospholipides spécifiques ou non d'un feuillet de la membrane vers l'autre feuillet. Les mouvements catalysées par ces enzymes nécessitent un apport d'énergie via l'hydrolyse de l'ATP car les lipides sont transportés contre leur gradient de concentration :
Les flippases sont donc impliquées dans la création et le maintien de l'asymétrie des 2 feuillets de certaines membranes comme la membrane plasmique ou le réseau trans-Golgi des Eucaryotes. Les flippases - ATPases de type P4 catalysent la translocation de lipides spécifiques. Par exemple, la flippase d'amino-phospholipides des érythrocytes :
Source : Niemela et al. (2009) En revanche, les flippases - transporteurs de type ABC (ou floppases) sont relativement peu spécifiques et catalysent plus lentement la translocation des lipides. |
![]()
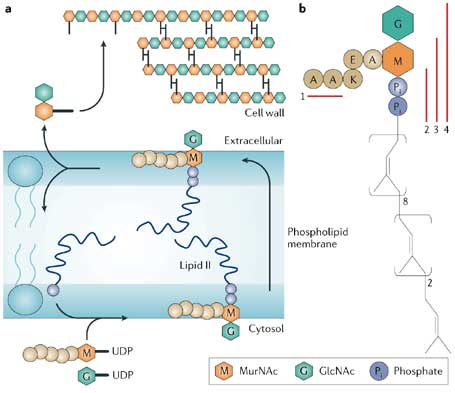
β. Rôle des flippases dans la biosynthèse de glycoconjugués tels que les lipopolysaccharides bactériens Figure a ci-dessous en haut : la paroi de toutes les bactéries contient un polymère de sucres aminés alternés : la N-acétylglucosamine (GlcNAc) et l'acide N-acétylmuramique (MurNAc). Ces polymères de glycanes sont réticulées par un pentapeptide dont la séquence est généralement [L-Ala-γ-D-Glu-diaminopimelyl (ou L-Lys) -D-Ala-D-Ala] attaché au sucre MurNAc. Cette réticulation confère à la cellule sa rigidité et sa résistance mécanique.
Source : Breukink & Kruijff (2006) Figure a en bas : le lipide II (plusieurs milliers de molécules par cellule bactérienne) est le transporteur d'isoprénoides C55 qui reçoit le muramyl-pentapeptide à la fin de la phase qui a lieu dans le cytoplasme de la biosynthèse du peptidoglycane. Tout en restant sur la face interne de la membrane cytoplasmique, le muramyl-pentapeptidyl-lipide II est glycosylé. Puis le disaccharyl-pentapeptidyl-lipide II est transloqué par des flippases sur la face externe de la membrane cytoplasmique. Lorsque qu'elle affleure à la surface externe de la membrane, la partie lipide II du disaccharyl-pentapeptidyl-lipide II est la cible de 2 types d'antibiotiques :
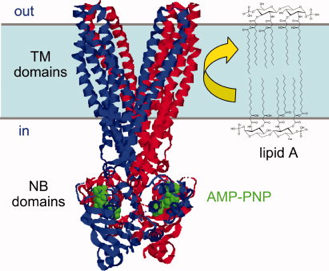
Le lipide A est une glucosamine disaccharide hexa-acylée. C'est un intermédiaire dans l'assemblage du lipopolysaccharide qui est le composant prédominant du feuillet externe de la membrane externe des bactéries Gram-négatives. La flippase MsbA (transporteur de type "ATP-binding cassette") transloque le lipide A de son site de synthèse situé dans le feuillet cytoplasmique (face interne de la membrane interne) vers le feuillet périplasmique où ont lieu les autres modifications de la structure de la tête du lipide A.
Source : Sharom (2011) Figure ci-dessous : structure du dimère de la flippase de lipides MsbA de Salmonella typhimurium (PDB : 3B60) et de son substrat, le lipide A bactérien. Deux molécules de nucléotides AMP-PNP (en vert) sont fixées au domaine de fixation des nucléotides.
Source : Sharom (2011) |
![]()
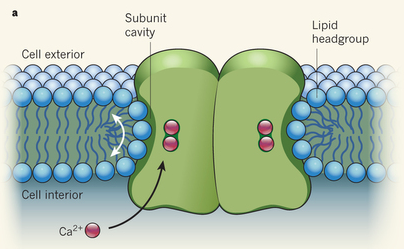
γ. Les scramblases Le mécanisme de ces enzymes est indépendant de l'énergie (Interpro : scramblase - IPR005552). Les scramblases ré-équilibrent la distribution des phospholipides communs chargés négativement entre les deux feuillets de la bicouche lipidique : on obtient une distribution symétrique. Elles jouent aussi un rôle dans la biosynthèse de glycoconjugués comme les glycosphingolipides, les protéines N-glycosylées et les protéines ancrées par une ancre glycosylphosphatidylinositol (GPI). Toutes les scramblases contiennent un domaine de fixation du calcium avec un motif de type "EF-hand". L'activité enzymatique de la scramblase dépend de la concentration de calcium intracellulaire. Dans des conditions normales, cette concentration est très faible : la scramblase a donc une activité faible en conditions de repos. La redistribution des phospholipides est déclenchée par une augmentation du calcium cytosolique. Cette redistribution semble dépendante de la scramblase : elle a pour résultat une distribution symétrique des phospholipides chargés négativement entre les deux feuillets de la bicouche lipidique.
Brunner et al. (2014) ont déterminé la première structure de nhTMEM16 (champignon Nectria haematococca), une scramblase activée par le calcium. Environ 40% de la séquence en acides aminés de nhTMEM16 est identique à celles de ses homologues chez les mammifères : il est possible qu'elles partagent une même structure de base et un mécanisme d'action commun. La scramblase nhTMEM16 est organisée en homodimère. Chaque sous-unité contient :
Source : Brunner et al. (2014) |
|
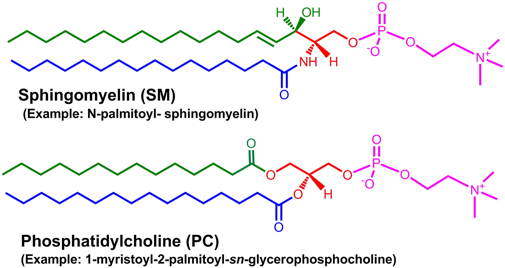
Les membranes plasmiques contiennent des combinaisons [glycosphingolipides / protéines récepteurs] qui forment des micro-régions de glyco-lipo-protéines appelées radeaux lipidiques ("lipid rafts"). Les radeaux lipidiques ont une composition en lipides différente de celle de la bicouche environnante (par exemple, ils contiennent 3 à 5 fois plus de cholestérol). En conséquence, les radeaux lipidiques ont une structure plus ordonnée et compacte que celle de la bicouche environnante mais ils "flottent" librement dans la bicouche. Ces micro-régions spécialisées des membranes interviendraient dans des processus cellulaires (bien que leurs rôles - et l'existence même de ces radeaux - ne soient pas encore définitivement démontrés) :
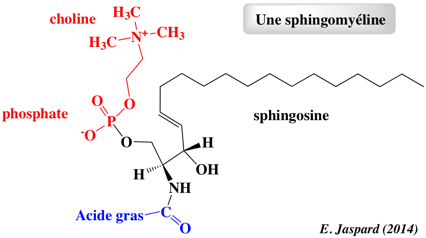
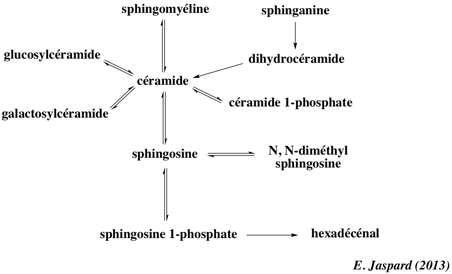
Chez les Eucaryotes, le principal composant des radeaux lipidiques sont les sphingomyélines qui représentent environ 10 à 15 % des phospholipides totaux (ce sont les sphingolipides membranaires les plus abondants) et davantage encore dans le cerveau et les tissus du système nerveux périphérique (gaînes de myéline). Les sphingomyélines sont absentes chez les végétaux et les micro-organismes. Les sphingomyélines sont composées d'une tête phosphocholine (rouge - figure ci-dessous), d'une sphingosine (noir) et d'un acide gras (bleu). Les sphingomyélines prédominantes contiennent l'acide palmitique ou l'acide stéarique N-acylés sur le carbone 2 de la sphingosine.
Les sphingomyélines et leurs métabolites sont des messagers secondaires dans la transduction de signaux lors du développement, de la différenciation et de la réponse immunitaire. Les sphingomyélines sont essentielles pour l'activité de certains récepteurs (exemples : récepteur nicotinique α7, récepteurs NMDA, récepteurs neurotrophiques, récepteur tyrosine kinase de type 2, récepteur sérotonine1A, récepteur de l'urokinase). |
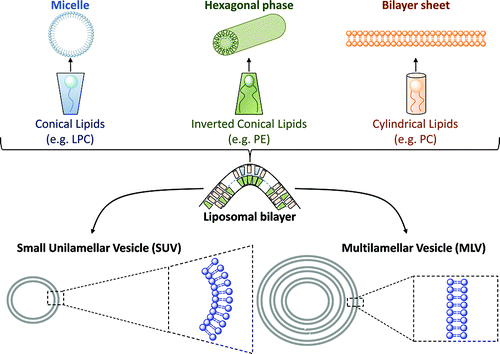
d. Liposomes et transport ciblé de médicaments Les lipides peuvent former des liposomes (figure ci-contre) : vésicule artificielle, composée d'une bicouche lipidique qui entoure un compartiment aqueux, tout comme les membranes cellulaires. Les liposomes diffèrent des micelles qui sont constitués d'une monocouche lipidique. Les liposomes sont souvent constituées de phospholipides enrichis en phosphatidylcholine et peuvent également contenir des chaînes lipidiques mixtes aux propriétés tensioactives comme la phosphatidyl-éthanolamine. Les nanoparticules peuvent véhiculer un médicament, un gène… au sein d'un tissu, d'une cellule, d'un compartiment cellulaire. Les nanoparticules sont en général constituées de polymères biodégradables. Les liposomes ressemblent à des cellules (biomimétiques) : le médicament est dissous dans le compartiment aqueux (figure à droite ci-dessous) s'il est hydrophile ou dans la bicouche s'il est lipophile.
|
| Avantages des liposomes | Inconvénients des liposomes |
| Liposomes increased efficacy and therapeutic index of drug (actinomycin-D) | Low solubility |
| Liposome increased stability via encapsulation | Short half-life |
| Liposomes are non-toxic, flexible, biocompatible, completely biodegradable, and non-immunogenic for systemic and non-systemic administrations | Sometimes phospholipid undergoes oxidation and hydrolysis-like reaction |
| Liposomes reduce the toxicity of the encapsulated agent (amphotericin B, Taxol) | Leakage and fusion of encapsulated drug/molecules |
| Liposomes help reduce the exposure of sensitive tissues to toxic drugs | Production cost is high |
| Site avoidance effect | Fewer stables |
| Flexibility to couple with site-specific ligands to achieve active targeting | ------------------------------ |
| Intérêts de véhiculer des médicaments via liposomes | Exemples |
| Improved solubility of lipophilic and amphiphilic drugs | Amphotericin B, porphyrins, minoxidil, some peptides, and anthracyclines, respectively; hydrophilic drugs, such as anticancer agent doxorubicin or acyclovir |
| Passive targeting to the cells of the immune system, especially cells of the mononuclear phagocytic system | Antimonials, amphotericin B, porphyrins, vaccines, immunomodulators |
| Sustained release system of systemically or locally administered liposomes | Doxorubicin, cytosine arabinoside, cortisones, biological proteins or peptides such as vasopressin |
| Site-avoidance mechanism | Doxorubicin andamphotericin B |
| Site-specific targeting | Anti-inflammatory drugs, anti-cancer, anti-infection |
| Improved transfer of hydrophilic, charged molecules | Antibiotics, chelators, plasmids, and genes |
| Improved penetration into tissues | Corticosteroids, anesthetics, and insulin |
| Source : Akbarzadeh et al. (2013) | |
La taille du liposome va dépendre de la technique utilisée pour le fabriquer. Le diamètre peut varier entre quelques dizaines de nanomètres et quelques dizaines de microns, c'est à dire d'un rapport de 1 à 1000.
Source : Paige et al. (2011) |
| Classification des vésicules lipidiques selon leurs structures et/ou préparation | |
| archeosomes | vésicules consistant en lipides d'origine archebactérie |
| cochleates | dérivés de liposomes suspendus dans une solution aqueuse de polymère |
| dendrosomes | famille de nanoparticules non toxiques, neutres, biodégradables, covalentes ou auto-assemblées, hyperbranchées, dendritiques et sphériques |
| vésicules séchées reconstituées | petites vésicules unilamellaires, contenant différents lipides ou des mélanges de lipides |
| éthosomes | systèmes beaucoup plus efficaces pour délivrer la substance au niveau de la peau (en termes de quantité et de profondeur) que les liposomes classiques ou les solutions hydroalcooliques |
| immunoliposomes | liposomes modifiés par des anticorps |
| immunosomes | préparés par l'ancrage de glycoprotéines pour préformer les liposomes |
| complexe immunitaire stimulant | assemblages sphériques, micellaires d'environ 40 nm. Ils sont faits d'un mélange Quil A, cholestérol et phospholipides |
| lipoplexes | complexes [ADN-lipides cationiques], vecteurs efficaces pour la transfection de cellules mais qui présentent certains inconvénients en raison de leur toxicité |
| LUVET ("Large Unilamellar VEsicles") | préparés par la technique d'extrusion (systèmes à haute pression) |
| niosomes ou novasomes | petites vésicules unilamellaires préparées avec des agents surfactants non-ioniques |
| liposomes sensibles au pH | il en existe 4 classes |
| liposomes polymérisés | vésicules de phosphatidyl-choline polymérisées (35-140 nm) |
| proliposomes | particules sèches et fluides qui forment immédiatement une dispersion liposomale au contact de l'eau |
| protéosomes | vésicules d'origine bactérienne solubilisées, puis précipitées au sulfate d'ammonium et dialysées contre un détergent. Les protéines et les peptides sont complexés à la membrane de manière non covalente, ce qui les rend hautement immunogènes |
| vésicules d'évaporation en phase inverse | vésicules formées par évaporation de l'huile dans une émulsion d'eau ce qui aboutit à de grands liposomes unilamellaires |
| liposomes "furtifs" / cachés | le revêtement des liposomes avec du polyéthylène glycol (polymère hydrophile synthétique) améliore leur stabilité et augmente leur demi-vie en circulation, rendant l'utilisation des glycolipides obsolètes. Le revêtement par le PEG inhibe l'adsorption de protéines et l'opsonisation des liposomes (recouvrement par l'opsonine) |
| liposomes sensibles à la température | considérés comme vecteur prometteur pour délivrer de manière spécifique le médicament au bon endroit dans la cellule. Ils sont préparés avec des lipides qui subissent une transition de phase gel => liquide cristallin à quelques degrés au-dessus de la température physiologique |
| transfersomes | composés de phosphatidylcholine et de cholate - vésicules ultra-déformables avec des propriétés de pénétration dans la peau améliorées |
| virosomes | petites vésicules unilamellaires contenant l'hémagglutinine de la grippe qui leur permet de fusionner avec les membranes d'endocytose |
| Source : Wagner & Vorauer-Uhl (2011) | |
|
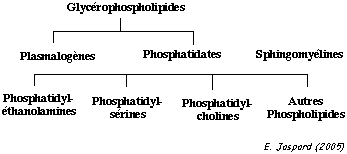
La structure en double couche de la membrane plasmique est due aux propriétés amphiphiles (ou amphipathiques) des multiples types de lipides qui la constituent. Il existe 3 classes principales de lipides membranaires : les phospholipides, les glycolipides et les stérols.
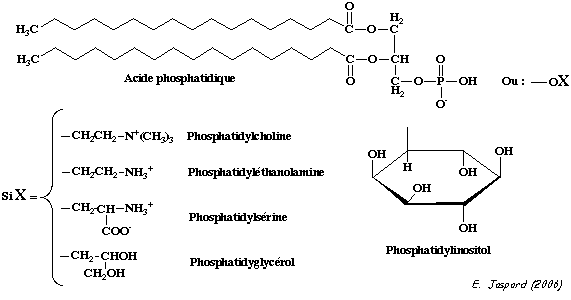
Parmi les les phospholipides, les glycérophospholipides sont les plus abondants. Leur structure est la suivante (figure ci-dessous):
Les cellules Eucaryotes contiennent des compartiments sub-cellulaires ou organites. C'est la composition spécifique [protéines / (phospho)lipides / oses] de chacune des membranes des organites qui est à l'origine d'une grande partie de leur propriétés et fonctions biologiques. |
| type de membrane (foie de rat) | cholestérol | PC | SM | PE | PI | PS | PG | DPG | PA | GL |
| plasmique | 30 | 18 | 14 | 11 | 4 | 9 | --- | --- | 1 | --- |
| RE rugueux | 6 | 55 | 3 | 16 | 8 | 3 | --- | --- | --- | --- |
| RE lisse | 10 | 55 | 12 | 21 | 7 | --- | --- | 2 | --- | --- |
| mitochondie - interne | 3 | 45 | 2,5 | 24 | 6 | 1 | 2 | 18 | 0,7 | --- |
| mitochondie - externe | 5 | 50 | 5 | 23 | 13 | 2 | 2,5 | 3,5 | 1,3 | --- |
| membrane nucléaire | 10 | 55 | 3 | 20 | 7 | 3 | --- | --- | 1 | --- |
| appareil de golgi | 7,5 | 40 | 10 | 15 | 6 | 3,5 | --- | --- | --- | --- |
| lysosome | 14 | 25 | 24 | 13 | 7 | --- | --- | 5 | --- | --- |
| myéline | 22 | 11 | 6 | 14 | --- | 7 | --- | --- | --- | 12 |
| érythrocyte | 24 | 31 | 8,5 | 15 | 2 | 7 | --- | --- | 0,1 | 3 |
| membrane plasmique - E. coli | 0 | 0 | --- | 80 | --- | --- | 15 | 5 | --- | |
| PC : phosphatidylcholine / SM : sphingomyèline / PE : phosphatidyléthanolamine / PI : phosphatidylinositol / PS : phosphatidylsérine / PG : phosphatidylglycérol / DPG : diphosphatidylglycérol (cardiolipine) / PA : acide phosphatidique / GL : glycilipides / RE : réticulum endoplasmique | ||||||||||
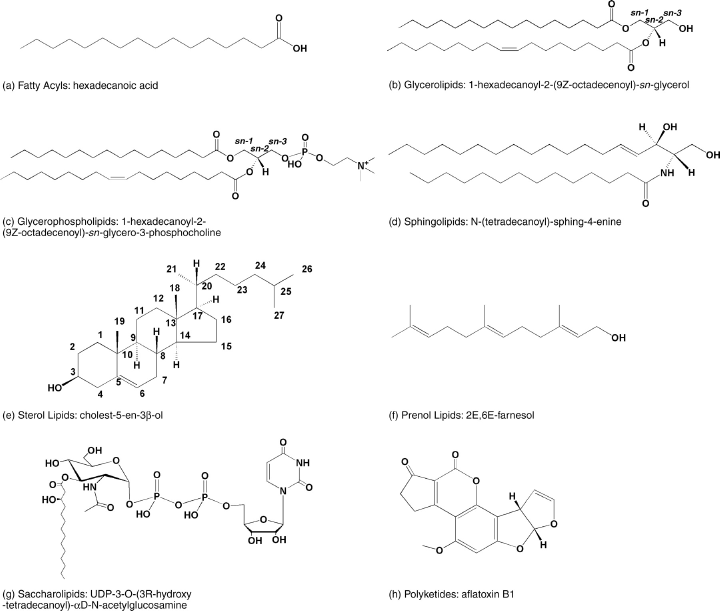
Base de données LIPID MAPS Structure Database (LMSD) Exemple : Glycerolipids -> Diradylglycerols -> 1,2-diacyl-sn-glycerol Remarques :
|
| Catégories | Exemples de sous-catégories | Exemple de lipide et de nomenclature |
| Acyls gras |
acide gras et conjugués |
acide dodécanoïque |
| Glycérolipides |
Monoradylglycérols, diradylglycérols, triradylglycérols Glycosylmonoradylglycérols Glycosyldiradylglycérols |
1-hexadécanoyl-2-(9Z-octadecenoyl)-sn-glycérol |
Glycéro- phospho- lipides |
glycérophosphocholine, glycérophosphoéthanolamine, glycérophosphoserine, glycérophosphoglycérol, glycérophosphate |
1-hexadécanoyl-2-(9Z-octadecenoyl)-sn-glycéro-3-phosphocholine |
| Sphingolipides | sphing-4-enine (sphingosine) |
|
| Stérol lipides |
cholestérol et dérivés cholesteryl esters phytosterols et dérivés stéroides C18 (estrogens) et dérivés, stéroides C19 (androgens) et dérivés, stéroides C21 (gluco/mineralocorticoides, progestogines) et dérivés secostéroides vitamines D2, D3 et dérivés [ST0301] acides biliares et dérivés glucuronides conjugués de la glycine, de la taurine hopanoides |
cholest-5-en-3β-ol |
| Prénol lipides |
C5 isoprenoides C5, C10 (monoterpenes), C15 (sesquiterpenes), C20 (diterpenes), C25 (sesterterpenes), C30 (triterpenes), C40 (tetraterpenes) polyterpenes quinones, hydroquinones, ubiquinones vitamines E, K polyprenols bactoprenols, bactoprenol monophosphates, bactoprenol diphosphates phytoprenols, phytoprenol monophosphates, phytoprenol diphosphates Dolichols, dolichol monophosphates, dolichol diphosphates |
2E,6E-farnesol |
| Saccharo lipides |
ose monoacylamino à ose heptaacylamino ose acylamino glycanes acyltrehaloses acyltréhalose glycanes |
UDP-3-O-(3R-hydroxy-tétradécanoyl)-αd-N-acétylglucosamine |
| Polyketides | polyketides aromatiques |
aflatoxine B1 |
| Source : LIPID MAPS Structure database (LMSD) | ||
Classification des lipides et exemple de chaque classe.
Source : Fahy et al. (2005) |
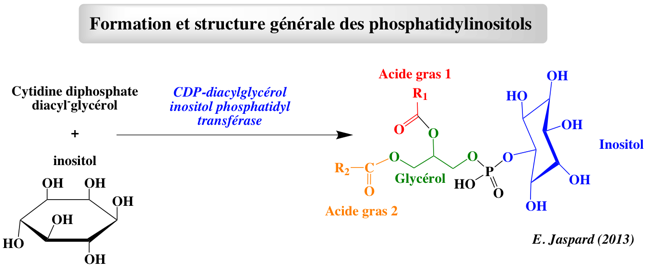
4. Les (glycéro)phosphatidylinositols et les phosphoinositides a. Formation et composition des phosphatidylinositols et des phosphoinositides Les phosphatidylinositols (abbréviation : PtdIns) font partie de la classe des glycérophospholipides.
Ils sont composés :
L'inositol peut-être phosphorylé de manière réversible sur les carbones 3, 4 et 5 et former les (glycéro)phosphatidylinositols phosphates (anioniques), également appelés inositides ou phosphoinositides :
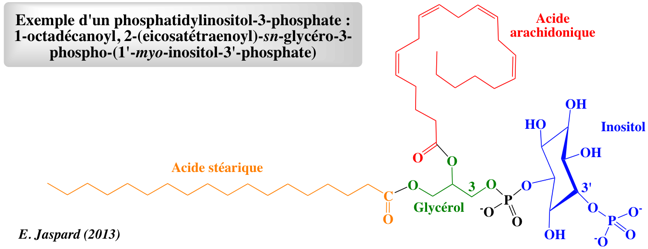
Exemple d'un phosphatidylinositol-3-phosphate qui joue un rôle important dans les membranes : sn-1-stearoyl-2-arachidonoyl phosphatidylinositol-3-phosphate.
Nom systématique : 1-octadécanoyl-2R-(5Z,8Z,11Z,14Z-eicosatétraenoyl)-sn-glycéro-3-phospho-(1'-myo-inositol-3'-phosphate). |
Les phosphoinositides sont des messagers secondaires très importants. Ils régulent des processus biologiques fondamentaux (croissance cellulaire, trafic membranaire, dynamique du cytosquelette, ...). La quantité et la distribution dans la cellule et dans le temps des phosphoinositides sont très variables. Les phosphoinositides sont situés exclusivement sur le feuillet cytosolique de la bicouche lipidique des membranes. Ils sont facilement accessibles aux phosphoinositides kinases, aux phosphoinositides phosphatases et aux phospholipases qui coupent les acides gras. Les enzymes de modification des phosphoinositides sont localisées de manière hétérogène dans la cellule.
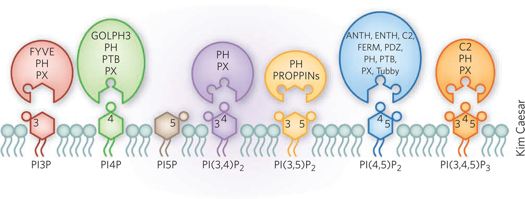
Source : Kutateladze (2010) Les phosphoinositides sont identifiés individuellement par un grand nombre de protéines effectrices qui contiennent des domaines structuraux spécialisés. Ces protéines effectrices reconnaissent l'arrangement unique de groupe(s) phosphate sur le cycle inositol. On dénombre actuellement 11 domaines de fixation des phosphoinositides qui couvrent un large éventail d'affinités et de sélectivité pour les lipides des membranes.
Source : Kutateladze T.G. (2010) Domaines des protéines effectrices (figure ci-dessus) : pleckstrin homology domain, ANTH (AP180 N-terminal homology), C2 (conserved region-2 of protein kinase C), ENTH (epsin N-terminal homology), FERM (4.1, ezrin, radixin, moiesin), FYVE (Fab1, YOTB, Vac1 and EEA1), GOLPH3 (Golgi phosphoprotein 3), PDZ (postsynaptic density 95, disk large, zonula occludens), PROPPINs (β-propellers that bind PIs), PTB (phosphotyrosine binding), PX (Phox homology), Tubby modules. Le domaine FYVE fixe avec une affinité et une spécificité élevées les PtdIns(3)P des endosomes précoces puis ponte ces PtdIns(3)P à des protéines cytosoliques.
|
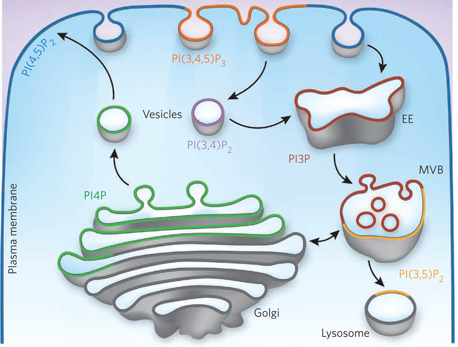
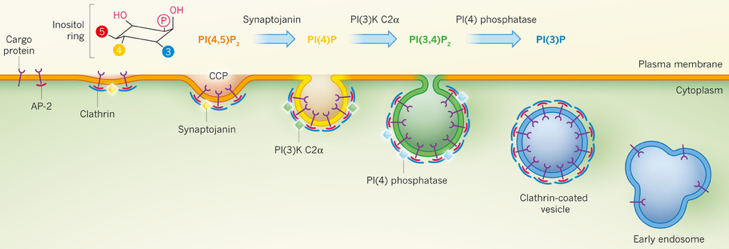
b. Interconversion de phosphatidylinositol phosphates et maturation des vésicules Afin que la cellule distinguent les vésicules qui doivent sortir (sécrétion) de celle qui assurent le traffic intracellulaire, les vésicules naissantes doivent acquérir instantanément une identité de membrane distincte de celle de la membrane dont elles sont issues. Les phosphatidylinositol phosphates (PIP) assurent ce marquage membranaire spécifique. Il existe un mécanisme d'interconversion spatiale et temporelle régulée de ces lipides au cours de la maturation des vésicules qui sont impliquées dans l'endocytose médiée par la clathrine, la voie principale pour l'internalisation des nutriments et des récepteurs impliqués dans la signalisation. Les différents types de phosphatidylinositol phosphate sont en interconversion rapide via des kinases de lipides et des phosphatases de lipides. Les kinases ajoutent des groupements phosphate sur les carbones 3, 4 et 5 du cycle de l'inositol d'un PIP et les phosphatases les enlèvent. La membrane plasmique est riche en phosphatidylinositol-4, 5-bisphosphate (PI(4,5)P2), dont la concentration est maintenue par l'activité de phosphatidylinositol-5-kinases. PI(4,5)P2 est essentiel pour l'endocytose médiée par la clathrine. De nombreux composants impliqués dans cette voie d'endocytose, y compris les protéines AP-2, se lient spécifiquement à PI(4,5)P2 avant de déclencher l'assemblage de la principale protéine d'enrobage, la clathrine, pour générer des "puits" recouverts de clathrine qui s'invaginent et forment les vésicules d'endocytose. Par contre, les endosomes (vésicules intracellulaires avec lesquelles les vésicules recouvertes de clathrine fusionnent éventuellement) sont riches en un autre PIP, le PI(3P), qui joue un rôle essentiel dans le traffic via les endosomes en recrutant plusieurs composants de la machinerie de fusion des endosomes. La maturation des "puits" recouverts de clathrine est accompagnée par la conversion progressive de PI(4,5)P2 en PI(4)P puis en PI(3,4)P2. Ce processus est médié par le recrutement séquentiel et l'activation de la phosphatidylinositol-5-phosphatase appelée synaptojanine et la phosphatidylinositol 3-kinase appelée PI(3)KC2α.
Source : Schmid & Mettlen (2013) Les surfaces des vésicules recouvertes de clathrine naissantes et les endosomes précoces avec lesquels elles peuvent fusionner, sont riches en PI(3)P générés par l'activité de la phosphatase PI(4). Le substrat préférentiel de la kinase PI(3)KC2α est PI(4)P, le produit de l'activité de la synaptojanine. La kinase PI(3)KC2α s'accumule progressivement sur les "puits" recouverts de clathrine tout au long de leur maturation. De plus, PI(3,4)P2 est une sous-population de ces "puits" qui s'enrichit. Un appauvrissement en kinase PI(3)KC2α induit une inhibition de l'endocytose médiée par la clathrine et prolonge la durée de vie des "puits" recouverts de clathrine et leur accumulation à des étapes intermédiaires de leur maturation. Ces résultats indiquent qu'il y a coordonnation entre la conversion de PI(4,5)P2 en PI(4)P puis en PI(3,4)P2 et la maturation des "puits" recouverts de clathrine. |
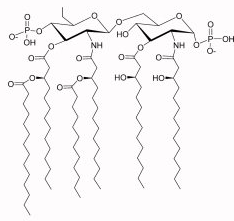
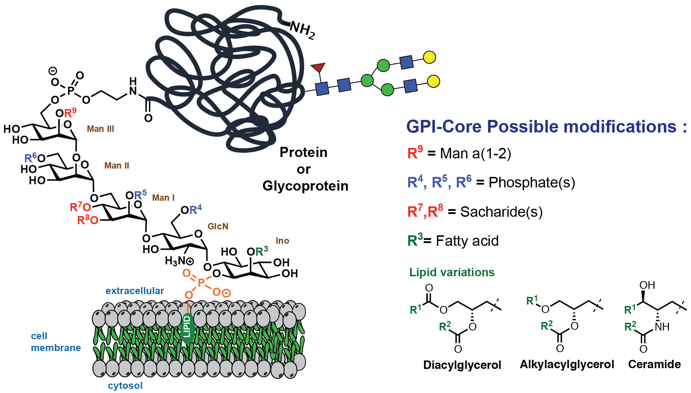
c. Les protéines attachées à la membrane par une ancre glycosylphosphatidylinositol Environ 20% des protéines sont attachées au feuillet externe de la membrane cellulaire par une unité glycosylphosphatidylinositol (GPI) dans toutes les cellules Eucaryotes. L'ancre GPI est une modification post-traductionnelle. Ce glycolipide contient :
Source : Max Planck Institute Cette structure de base GPI peut avoir des modifications supplémentaires selon le type de cellule, les plus courantes étant la phosphorylation, la glycosylation ou l'acylation (laision covalente entre un acide gras sous forme acyl-CoA et un acide aminé). L'extrémité réductrice de la glucosamine (GlcN) est liée à un groupement phosphatidylinositol. Ce phosphatidylinositol est alors ancré par une autre liaison phosphodiester à la membrane. Beaucoup de protéines ancrées via une ancre GPI sont des enzymes hydrolytiques ou ont un rôle de récepteurs, d'antigènes de surface des cellules ou de molécules d'adhérence cellulaire. De plus, les ancres GPI sont facilement incorporées dans les régions de la membrane enrichies en cholestérol et en sphingolipides, les radeaux lipidiques. Cela suggère que les protéines-GPI jouent un rôle important dans la transduction du signal. |
|
5. Relation entre lipides membranaires et protéines transmembranaires a. Corrélation entre nombre de lipides fixés et structure des protéines transmembranaires Les lipides de "première couche" qui entourent les protéines transmembranaires s'échangent avec ceux de la bicouche environnante. Ces lipides de "première couche" peuvent être caractérisés par spectroscopie de résonance paramagnétique électronique (RPE) parce que leur mobilité rotationnelle est significativement perturbée par la surface transmembranaire des protéines avec lesquelles ils interagissent. Thermodynamiquement, la "première couche" de lipides se caractérise par :
|
| Protéines | Nb | Masse molaire (x10-3) | nα : nombre prédit ou réel |
| Cytochrome c oxidase | 56 ± 5 | 204 | 28 |
| 55 ± 4 | 28 | ||
| 40 ± 7 | 250 | 20 | |
| 42 ± 7 | 268 | 20 | |
| Cytochrome c réductase | 38 ± 3 | 230 | 13 |
| [Na+,K+]-ATPase (rein) | 31.5 ± 1.5 | 157 | 12 |
| [Na+,K+]-ATPase (requin) | 33 ± 3 | 147 | 12 |
| Ca2+-ATPase | 22 ± 2 | 115 | 10 |
| 24 ± 5 | 10 | ||
| Rhodopsine (boeuf) | 25 ± 3 | 39 | 7 |
| 22 ± 2 | 7 | ||
| Rhodopsine (grenouille) | 23 ± 2 | 39 | 7 |
| transporteur [ADP - ATP] | 25 | 32.8 | 6 |
| protéolipide de la myeline | 10 ± 2 | 25 | 4 |
| protéolipide 16 kDa | 5–6 | 17.5 | 4 |
| M13 phage coat protein | 4–5 | 5.2 | 1 |
| Phospholambane (PLB) | 5.7 ± 0.7 | 6.12 | 3.5 ± 0.4 |
| PLB - A37 | 11.8 ± 0.6 | 6.08 | 1.15 ± 0.15 |
| PLB - A36,41,46 | 7.1 ± 0.2 | 6.11 | 1.9 ± 0.1 |
| 4.0 ± 0.1 | 4.9 ± 0.1 | ||
| 11.3 | 1.0 | ||
| 7.8 ± 0.3 | 2.2 ± 0.5 | ||
| PLB - Δ1-25A36,41,46 | 4.0 ± 0.2 | --- | 5.1 ± 0.6 |
| 4.0 ± 0.2 | 5.0 ± 0.5 | ||
| 10.1 ± 1.3 | 1.2 ± 0.2 | ||
| 6.0 ± 0.6 | 3.3 ± 0.4 | ||
| Gramicidine A | 3.6 ± 0.3 | 1.88 | β6.3-helix |
| K27, K-channel peptide | 2.2 | 3.16 | β-sheet |
| K27-Δ2 | 2.5 | 3.04 | β-sheet |
| M13 phage coat protein | 4 | 5.2 | β-sheet |
| OmpA | 11 | 18.90 | β-barrel |
| OmpG | 20 | 32.78 | β-barrel |
| FomA | 23 | 40.28 | β-barrel |
| FhuA | 32 | 80.09 | β-barrel |
| Source : Marsh D. (2008) | |||
| b. Sélectivité des protéines transmembranaires vis-à-vis des lipides constitutifs des membranes |
| Energies libres d'association (par rapport à la phosphatidylcholine ou PC - dernière colonne) de divers phospholipides interagissant avec des protéines ou peptides transmembranaires | |||||||
| Protéine | CL | PA | SA | PS | PG | PE | PC |
| PLP | – | − 2.34 | − 1.87 | − 0.79 | − 0.59 | – | 0.0 |
| DM-20 | – | − 0.83 | − 0.69 | − 0.10 | 0.0 | – | 0.0 |
| myelin proteolipide | − 0.41 | − 1.06 | − 1.95 | − 0.34 | − 0.10 | + 0.69 | 0.0 |
| − 1.10 | − 0.88 | − 1.06 | − 0.34 | − 0.69 | − 0.53 | 0.0 | |
| [Na+,K+]-ATPase | − 1.34 | − 0.41 | − 0.53 | − 0.53 | + 0.11 | + 0.11 | 0.0 |
[Na+,K+]-ATPase digéréeée |
– | – | −0.41 − 1.03 | − 0.64 | – | – | 0.0 |
| SERCA Ca2+-ATPase | – | 0.0 | + 0.7 | 0.0 | – | + 0.8 | 0.0 |
| cytochrome c oxidase | − 1.69 | − 0.64 | – | 0.0 | 0.0 | 0.0 | 0.0 |
| transporteur [ADP - ATP] | − 1.34 | − 1.46 | − 1.41 | − 0.88 | + 0.22 | – | 0.0 |
| − | − 0.99 | − 1.41 | + 0.36 | − | − 0.10 | 0.0 | |
| − | − 0.10 | −0.41 − 0.92 | − 0.79 | − | – | 0.0 | |
| − 1.63 | – | − 1.59 | − 0.99 | − 0.53 | + 0.69 | 0.0 | |
| M13 phage coat protein | − 1.44 | − 1.44 | − 0.83 | − 0.74 | − 0.47 | + 0.11 | 0.0 |
| – | − 0.47 | − 0.18 | − 0.18 | − 0.10 | 0.0 | 0.0 | |
| cytochrome c reductase | − 0.34 | − 0.88 | − 0.92 | − 0.59 | − 0.53 | − 0.26 | 0.0 |
| protéolipide 16 kDa | – | – | − 0.88 | − 0.41 | − 0.34 | – | 0.0 |
| K27, K-channel peptide | – | − 1.19 | − 0.69 | − 0.69 | − 0.10 | – | − 0.10 |
| K27-Δ2 | – | − 1.46 | − 1.46 | − 0.92 | 0.0 | – | 0.0 |
phospholambane - |
– | − 0.26 | − 1.03 | 0.0 | 0.0 | + 0.11 | 0.0 |
phospholambane |
– | − 0.10 | − 0.59 | 0.0 | 0.0 | + 0.22 | 0.0 |
| GalP | – | 0.0 − 1.06 | −0.64 − 1.46 | − 0.01 | + 0.11 | 0.0 | 0.0 |
| Rhodopsine | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Gramicidine A | – | − 0.18 | + 0.36 | − 0.10 | + 0.22 | + 0.22 | 0.0 |
| OmpAj | – | − 0.92 | + 1.6 | +.60 | 0.0 | + 0.69 | + 0.51 |
| OmpG | – | − 0.17 | + 0.4 | – | − 0.13 | + 0.09 | 0.0 |
| FomA | – | − 0.19 | − 1.41 | − 0.46 | − 0.13 | − 0.06 | 0.0 |
| FhuAj | – | − 0.41 | − 1.10 | − 0.34 | 0.0 | + 0.51 | − 0.10 |
| OmpF | – | – | – | – | + 0.6 | + 0.2 | 0.0 |
| KcsA | + 0.9 | − 0.3 | – | + 0.2 | − 0.23 | + 0.6 | 0.0 |
| MscL | − 0.6 | − 0.6 | − | − 0.8 | − 0.6 | − 0.1 | − 0.0 |
CL : cardiolipine - PA : acide phosphatidique - SA : acide stéarique - PS : phosphatidylserine - PG : phosphatidylglycérol - PE : phosphatidyléthanolamine - PC : phosphatidylcholine |
|||||||
|
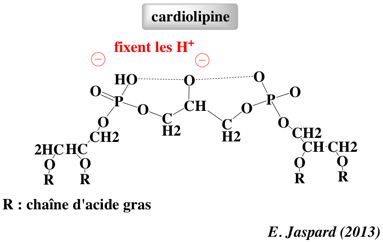
La cardiolipine (diphosphatidylglycérol) est un glycérophospholipide des membranes des bactéries et des mitochondries. La cardiolipine représente 18 à 20% des phospholipides de la membrane interne des mitochondries.
La cardiolipine est un lipide caractéristique des membranes qui développent un potentiel électrochimique via le transport des électrons pour la synthèse d'ATP. La cardiolipine a de nombreux rôles :
|
6. Environnement biophysique et caractéristiques des protéines transmembranaires a. Différence entre membranes réelles et conditions expérimentales pour la détermination de la structure des protéines transmembranaires
La principale raison est la très grande difficulté à solubiliser les protéines transmembranaires. L'autre difficulté est de reproduire expérimentalement les conditions biophysiques des membranes biologiques quand on purifie/isole une protéine transmembranaireet pour en déterminer la structure. Quelques exemples historiques :
Une caractéristique qui distingue les phospholipides des autres molécules amphiphiles est la concentration critique pour l'auto-assemblage extrêmement faible dans le cas des phospholipides :
|
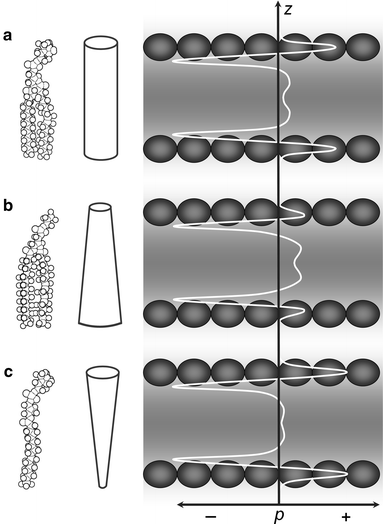
c. Origine physique de la pression latérale subie par les protéines transmembranaires L'une des différences majeures entre les protéines membranaires et les protéines cyto-solubles est la pression à laquelle elles sont exposées. Une protéine libre en suspension en solution aqueuse subit une pression isotrope, alors que la pression que subit une protéine membranaire varie considérablement le long de la bicouche lipidique. Les pressions locales peuvent s'élever à plusieurs centaines d'atmosphères.
De plus, les têtes hydrophiles subissent une répulsion électrostatique entre les groupements chargés et la perte de molécules d'eau d'hydratation du fait de cette compression. En ce qui concerne les chaînes hydrophobes d'hydrocarbures, le raidissement des chaînes acyles (pénalisée du point de vue de l'entropie) contribue à cette pression latérale positive. Formes schématiques de phospholipides (à gauche) et profils de pression latérale (à droite) dans les bicouches lipidiques.
Source : Fiedler et al. (2010) La pression latérale (p) est reportée en fonction de la profondeur (z) de la bicouche.
La composante négative de la pression latérale est due à la tension à l'interface entre les têtes hydratées et les chaînes acyles des lipides et reste donc pratiquement constante. A l'équilibre, l'intégrale de la pression au travers de la bicouche est toujours nulle : les surfaces cumulées sous la courbe p(z) sont égales pour les composantes de la pression positive et de la pression négative. |
|
7. Les protéines membranaires et transmembranaires Dans le génome humain, on estime qu'il y a environ 5500 gènes qui codent une protéine avec une région transmembranaire en hélice, soit environ 26 % des gènes codant des protéines. Le plus grand nombre de ces protéines membranaires n'aurait qu'une région transmembranaire prédite, mais il y a aussi beaucoup de protéines avec 7 régions transmembranaires prédites (notamment les récepteurs couplés aux protéines G).
Source : Jacso et al. (2012) En revanche, les protéines membranaires dont les structures ont été déterminéesne représentent environ que 2% de l'ensemble des structures connues (recensées dans la PDB) : un grand nombre de repliements caractéristiques des protéines membranaires sont donc encore inconnus.
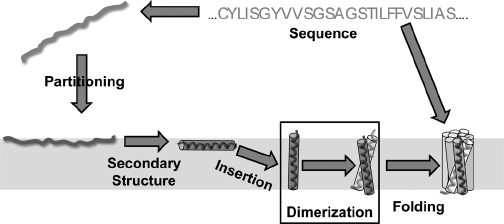
Cependant, les protéines membranaires sont soumises à des contraintes biophysiques, imposées par l'architecture de la bicouche lipidique, auxquelles ne sont pas soumises les protéines cytosolubles. Ces contraintes changent probablement la nature du processus du repliement des protéines membranaires qui, actuellement, est décrit par cinq étapes :
Source : Li et al. (2012)
La grande majorité des protéines membranaires sont synthétisées dans le réticulum endoplasmique, puis elles sont incorporées dans les membranes des organites où elles doivent fonctionner. Aller à la base de données "Orientation of Proteins in Membranes - OPM". |
| Quelques exemples de protéines membranaires | |||
| Fonctions - Rôles | Catégorie | Exemple - commentaires | Code PDB |
| Canaux et pores | Canaux ioniques trimèriques | "Degenerin/epithelial sodium channel" Récepteurs P2X "Acid sensing ion channels" |
|
| Canaux ioniques tétramèriques | "Voltage-gated ion channels" | ||
| "Pentameric ligand-gated ion channels" | Récepteur nicotinique de l'acétylcholine | ||
| Canaux ioniques hexamèriques | Jonctions gap ("Gap junctions") : complexe constitué de 2 semi-canaux (les connexons) chacun constitué de 6 connexines | ||
| Canaux ioniques viraux | "M2 channel of influenza A virus" | ||
| "Mechanosensitive channels" | MscL ("large conductance"), MscS ("small conductance"), MscM ("mini conductance") et MscK (regulées par la concentration du potassium) de Escherichia coli | ||
| Aquaporines | ------------ | 2B60 | |
| "Ammonia channel" | ------------ | 1U7G | |
| Toxines formant des pores | ClyA / α-HL | 2WCD / 7AHL | |
| Transports actifs | Transports primaires | ATPase de type P | |
| "Light-driven pumps" - famille de la bactériorhodopsine | 1FBB / 1FBK | ||
| "ATP-binding cassette (ABC) transporters" : glycoprotéine P / flippase MsbA | 3G61 | ||
| Transports secondaires : l'une des familles les plus vastes de protéines membranaires | Transporteurs [ATP/ADP] | 1OKC | |
| Transporteurs dépendant du sodium | 2NWX | ||
| Protéines de translocation | Canaux de translocation des protéines : SecYEβ / SecA–SecYEG | 1RHZ / 3DIN | |
| Chaîne de transport des électrons | ------------ | Complexe I - NADH:ubiquinone réductase | |
| Complexe II - succinate:ubiquinone réductase | 1ZOY | ||
| Complexe III - ubiquinol:cytochrome c réductase | 2FYU | ||
| Complexe IV - cytochrome c oxydase (aa3) | 1V54 | ||
| Photosynthèse | ------------ | Centre réactionnel | 1PRC |
Complexe LHC1 - Centre réactionnel |
1PYH | ||
| Complexe du photosystème I (PSI) | 2O01 | ||
| Complexe du photosystème II (PSII) | 3BZ1 | ||
| Réceptors couplés aux protéines G | ------------ | Rhodopsine | 1GZM |
| Récepteur β1-adrénergique | 2VT4 | ||
| Récepteur β2-adrénergique | 2RH1 | ||
| Récepteur adénosine A2a | 3EML | ||
| Enzymes membranaires | Protéases intramembranaires | Rhomboides GlpG (protéases à sérine à 6 domaines transmembranaires) | 2IC8 |
| Protéase site 2 | 3B4R | ||
| Thiol oxidases | |||
| Protéines MAPEG | "Membrane associated proteins in eicosanoid and glutathione metabolism" | 2H8A | |
| Méthane mono-oxygénase | ------------ | ||
| Protéines membranaires monotopiques | Carnitine palmitoyl transférase |
||
| Source : Vinothkumar & Henderson (2010) | |||
|
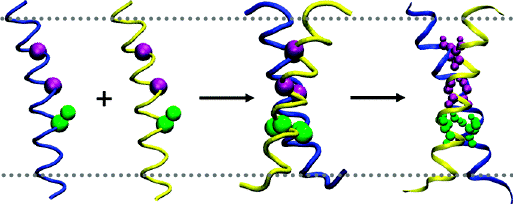
b. Les hélices transmembranaires Voir un cours sur les structures secondaires des protéines. La découverte (Lemmon et al., 1992) d'un motif (LIxxGVxxGVxxT) de dimérisation d'hélices transmembranaires de la glycophorine A (GpA) composé exclusivement d'acides aminés petits et apolaires a attiré l'attention sur l'importance des forces de van der Waals dans le repliement des protéines transmembranaires.
Ainsi, les interactions hélices - hélices initiales puis la formation de dimères seraient des événements clés du repliement des protéines membranaires polytopiques.
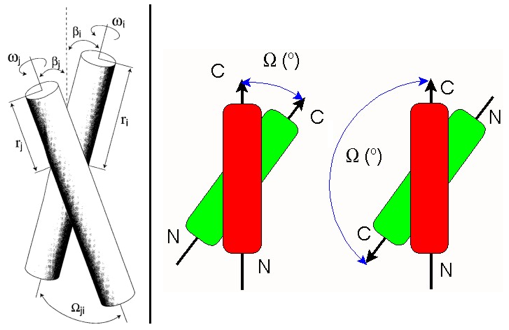
Source : Psachoulia et al. (2010) L'obtention d'un nombre croissant de structure de protéines membranaires et transmembranaires a apporté un grand nombre d'informations quant à la topologie adoptée par les hélices membranaires et transmembranaires : elle peuvent être courtes, longues, déformées, interrompues au milieu de la membrane, elles peuvent traverser la membrane de manière oblique, reposer à plat sur la surface de la membrane, traverser une partie de la membrane puis retourner en arrière (formant des boucles appelées "re-entrant loops"). Figures ci-dessous : représentation schématique des paramètres géométriques décrivant la conformation d'un faisceau d'hélices :
Figure adaptée de : Torres et al. (2002) et Zloh et al. L'angle d'intersection est un paramètre qui traduit la compaction d'un assemblage d'hélices. Les valeurs de cet angle (Ω dans la figure ci-contre) sont calculées sur la base d'un dièdre défini par 4 points : 1 point à l'extrémité de chaque axe d'hélice et les 2 points (sur l'axe des hélices) de plus grande approche des hélices.
|
| Structures et propriétés d'hélices dimèriques transmembranaires (Cymer et al., 2012) | ||||
| Protéine | PDB | Méthode | Acides aminés impliqués dans l'assemblage des hélices | angle d'intersection |
| GpA | 1afo | RMN | LIxxGVxxGVxxT | − 40 |
| ErbB2 | 2jwa | RMN | LTxxISAxVGI | − 41 |
| ErbB1/ErbB2 | 2ks1 | RMN | ErbB1: IxTGMxGAxLLxxV | − 46 |
| ErbB2 : TxxISAxVGIxLV | ||||
| ErbB3 | 2l9u | RMN | IxxLVxIFxxLxxxFLxxR | + 24 |
| αIIbβ3 | 2k9j | RMN | αIIb : GxxxGxxLL | − 25 |
| β3: VMxxxILxxG | ||||
| EphA1 | 2k1k | RMN | AVxxGLxxGAxxLL | − 39 à – 48 |
| EphA2 | 2k9y | RMN | LAxIGxxAVxxVVxLVxxxxxFF | + 15 |
| BNIP3 | 2j5d | RMN | FxxxFxxxLxxSHxxAxxxGxxIG | − 45 |
| BNIP3 | 2ka1, 2ka2 | RMN | SHxxAIxxGIxxG | − 34 |
| ζζ | 2hac | RMN | CxxxDxxLxxYxxxLTxxFxxV | + 23 |
| DAP12 | 2l34 | RMN | LxxIVxxDxxLT | + 20 |
| Sx1A/Syb2 | 3hd7 | Diffraction rayons X | Syb2 : MxxILxxIxxxIxxIIxxY | + 18 |
| Sx1A : IxxCxxILxxIxxxT | ||||
|
Visualisation de la bactériorhodopsine de Halobacterium salinarum. Code PDB : 2BRD |
c. Différences de composition en acides aminés entre les hélices transmembranaires et les hélices des protéines hydrosolubles Bien que reconnus comme casseurs d'hélice, Gly et Pro sont très abondants tout le long des hélices transmembranaires.
Voir un cours sur les acides aminés. En d'autres termes, la stabilité des hélices (résultant de la faible constante diélectrique et du manque d'eau dans le coeur hydrophobe des membranes) est amoindrie par Gly et Pro, mais ce phénomène est largement compensée par un accroissement de la stabilité de la structure tertiaire. Les hélices transmembranaires sont enrichies en Phe et Trp :
Le nombre d'acides aminés chargés (Asp, Glu, Arg, Lys - figure a ci-contre) ou très polaires (His, Gln et Asn - figure b ci-contre) est réduit d'un facteur 3 - 4 dans les hélices transmembranaires.
Source : Zhou & Cross (2013) Quand ils sont présents, ces acides aminés sont plus souvent situés aux extrémités (N- et C-) des hélices transmembranaires où les chaînes latérales interagissent avec les têtes hydrophiles des phospholipides. Les chaînes latérales faiblement polaires (Ser et Thr) sont parfois trouvés à la surface hydrophobe des protéines transmembranaires. |
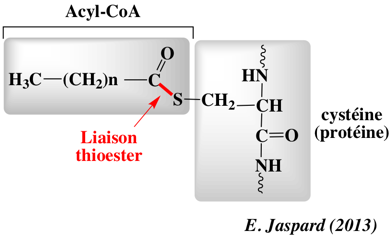
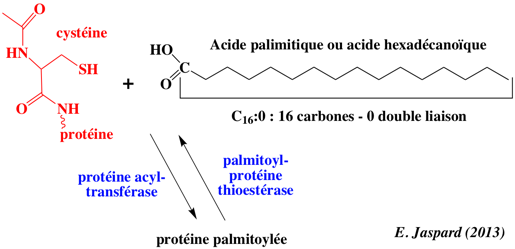
d. Ancrages à la membrane des protéines membranaires par S-palmitoylation La S-palmitoylation (ou S-acylation) est une modification post-traductionnelle qui ajoute, dans la grande majorité des cas, un acide palmitique (acide gras saturé en C16) sur 1 à 4 cystéine(s) d'une protéine membranaire, via une liaison thioesther. palmitoyl-CoA + [protéine]-L-cysteine <=> [protéine]-S-palmitoyl-L-cysteine + CoA
Les protéines sont souvent modifiées de manière séquentielle par des lipides différents :
La palmitoylation a lieu dans le cytoplasme et n'utilise pas les voies de sécrétion du réticulum endoplasmique. La palmitoylation augmente l'hydrophobicité de la protéine et donc son aptitude à s'associer aux membranes. La palmitoylation est essentielle pour le repliement des protéines membranaires impliquées dans la signalisation et le trafic intracellulaires comme les kinases de la famille Src, les GTPases de la famille Ras, les protéines G et les récepteurs couplés aux protéines G . La palmitoylation est aussi impliquée dans le métabolisme des lipoprotéines, l'apoptose et d'autres processus cellulaires. |
| Exemples d'acides gras | |||||
| Carbone | Doubles liaisons | Position des doubles liaisons | Nom commun | Nom IUPAC | Point de fusion |
| 12 | 0 | C12:0 | laurate (acide laurique) |
dodécanoate (acide dodécanoique) |
43°C |
| 14 | 0 | C14:0 | myristate | tétradécanoate | 52°C |
| 16 | 0 | C16:0 | palmitate | hexadécanoate | 63°C |
| 18 | 0 | C18:0 | stéarate | octadécanoate | 70°C |
| 20 | 0 | C20:0 | arachidate | icosanoate | 75°C |
| 22 | 0 | C22:0 | béhénate | docosanoate | 81°C |
| 24 | 0 | C24:0 | lignocérate | tétracosanoate | 84°C |
| 16 | 1 | C16:1 Δ9 | palmitoléate | cis-Δ9-hexadécénoate | - 0,5°C |
| 18 | 1 (végétaux) | C18:1 Δ9 (acide gras ω-9) | oléate | cis-Δ9-octadécénoate | 13°C |
| 18 | 2 (végétaux) | C18:2 Δ9,12 (acide gras ω-6) | linoléate | cis,cis-Δ9,12-octadécadiénoate | - 9°C |
| 18 | 3 (végétaux) | C18:3 Δ9,12,15 (acide gras ω-3) | linolénate | all-cis-Δ9,12,15-octadécatriénoate | - 17°C |
| 18 | 4 (végétaux) | C18:4 Δ6,9,12,15 (acide gras ω-3) | stéaridonate | all-cis-Δ6,9,12,15-octadécatétraénoate | ------ |
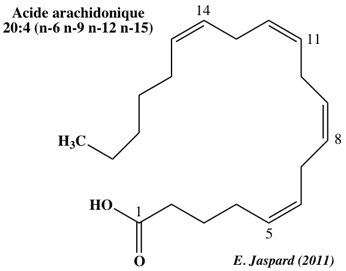
| 20 | 4 (animaux) | C20:4 Δ5,8,11,14 (acide gras ω-6) | arachidonate | all-cis-Δ5,8,11,14-eicosatétraénoate | - 49°C |
| 20 | 5 (poisson) | C20:5 Δ5,8,11,14,17(acide gras ω-3) | eicosapentaénoate | all-cis-Δ5,8,11,14,17-eicosapentaénoate | ------ |
| 22 | 5 (animaux) |
C22:5 Δ4,7,10,13,16 (acide gras ω-6) C22:5 Δ7,10,13,16,19 (acide gras ω-3) |
acide osbond clupanodonate |
all-cis-Δ4,7,10,13,16-docosapentaénoate all-cis-Δ7,10,13,16,19-docosapentaénoate |
------ |
| 22 | 6 (animaux) | C22:6 Δ4,7,10,13,16,19 (acide gras ω-3) | cervonate | all-cis-Δ4,7,10,13,16,19-docosahexaenoate | - 44°C |
| 24 | 6 | C24:6 Δ6,9,12,15,18,21 (acide gras ω-3) | nisinate | all-cis-Δ6,9,12,15,18,21-tétracosahexaénoate | ------ |
|
La S-palmitoylation est catalysée par les palmitoyl S-acyltransférases fixées à la membrane. A l'inverse de la prénylation et de la myristoylation, la palmitoylation est en général réversible du fait de la nature chimique de la liaison thioester établie entre l'acide gras et l'acide aminé. La dé-palmitoylation est catalysée par les palmitoyl-protéines thioesterases (EC 3.1.2.22).
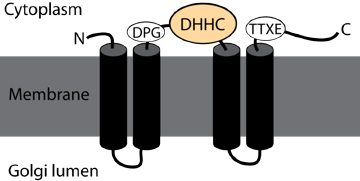
Ce processus de [palmitoylation - dépalmitoylation] est la seule modification par des qui soit réversible. Cette dynamique d'états modifiés ou pas est un élément clé de la régulation des protéines membranaires palmitoylées, au même titre que la [phosphorylation (kinases) - déphosphorylation (phosphatases)] des protéines. Palmitoyl-protéines thioesterases : PDB 3GRO. Les palmitoyl S-acyltransférases (EC 2.3.1.225) contiennent un domaine conservé riche en cystéine avec un motif [DHHC] nécessaire à l'activité enzymatique, raison pour laquelle on les appelle aussi protéines DHHC. Ce motif est lui-même inclus dans une région plus grande d'environ cinquante résidus d'acides aminés très conservés, dont le le motif général est [C-x(2)-C-x(9)-HC-x(2)-C-x(4)-DHHC-x(5)-C-x(4)-N-x(3)-F].
Source : PF01529 Les protéines S-acyltransférases s'auto-acylent. Les palmitoyl acyltransférases DHHC5, DHHC6 et DHHC8 sont S-acylées sur 3 résidus Cys au niveau d'un motif [CCX(7-13)C (S/T)] situé en aval du domaine DHHC conservé. Cette auto-acylation est un mécanisme qui régule probablement la spécificité et/ou l'activité des acyltransférases. |
|
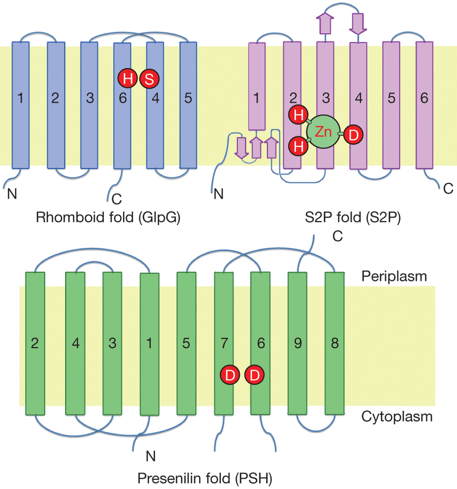
8. Les aspartyl protéases intramembranaires L'intérieur des membranes est un environnement très hydrophobe. La découverte de protéases utilisant des molécules d'eau pour l'hydrolyse d'autres protéines à l'intérieur des membranes fût donc une surprise. Cependant, la signalisation via la protéolyse intramembranaire est un mécanisme conservé des bactéries à l'homme. L'étape centrale de ce type de signalisation est l'hydrolyse de protéines cibles situées dans la bicouche lipidique par des protéases elles-mêmes enchâssées dans la membrane. Ces protéases intramembranaires se répartissent en 3 familles distinctes :
La préséniline est le composant catalytique de la γ-sécrétase (complexe protéolytique très hydrophobe constitué de 4 protéines intégrales membranaires : préséniline, PEN-2, APH-1 et nicastrine) responsable de la production du peptide amyloïde-β à partir de la protéine précurseur de l'amyloïde. Dans ce complexe, PEN-2 est supposé interagir directement avec la préséniline, qui contient 2 fragments d'endo-protéolyse appelées NTF (segments transmembranaires 1 à 6) et FCT (segments transmembranaires 7 à 9).
Source : Li et al. (2013)
|
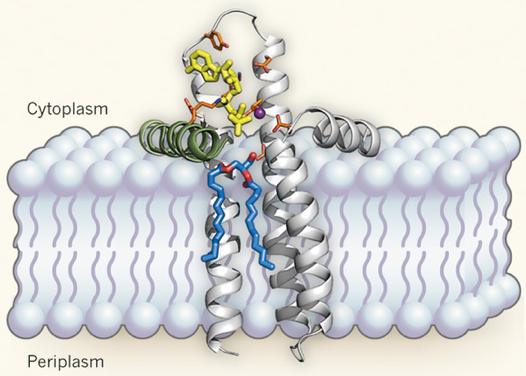
9. Un cas très particulier : le site actif de la diacylglycérol kinase bactérienne La diacylglycérol kinase bactérienne est une protéine intégrale de la membrane qui catalyse une étape clé dans la synthèse d'oligosaccharides (la conversion du diacylglycérol en acide phosphatidique). Elle forme un homotrimère et elle est fonctionnellement et structuralement distincte des autres kinases. De plus sa petite taille (seulement 121 acides aminés) la rend encore plus intrigante : comment une si petite enzyme peut-elle traverser la membrane tout en structurant un site actif qui doit accommoder un substrat lipidique encombrant et une molécule d'ATP hydrophile ? Les données cristallographiques récentes (Li et al., 2013) sont en faveur de l'hypothèse que chaque monomère "emprunte un composant structural" au monomère voisin et peut ainsi créer un site actif composite. Cet élément emprunté est une hélice amphiphile (hydrophobe et hydrophile) N-terminale. Cette hélice est stratégiquement située à l'interface entre la membrane et le cytoplasme et fournit un résidu catalytique crucial. Figure ci-dessous : schéma de la diacylglycérol kinase dans la membrane interne de Escherichia coli.
Source : Zheng & Jia (2013) bâtonnets oranges : acides aminés catalytiques - sphère pourpre : ion magnésium L'hypothèse formulée par Li et al. (2013) est que le site actif qui lie l'ATP (jaune) et le substrat (bleu et rouge), comprend à la fois la membrane et une hélice N-terminale d'un autre monomère (vert). La poche de fixation de l'ATP est dans la partie cytoplasmique de la protéine : elle est formée par les trois hélices transmembranaires d'un monomère et l'hélice N-terminale du monomère voisin. La poche de liaison du substrat est formée en grande partie par la membrane. Ainsi la membrane constituerait une partie du site actif de cette enzyme. |
|
Les protéines MAPEG ("Membrane Associated Proteins in Eicosanoid and Glutathione metabolism") sont impliquées dans la biosynthèse de dérivés de l'acide arachidonique.
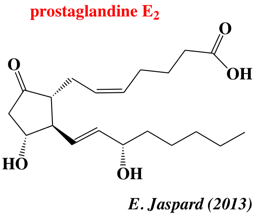
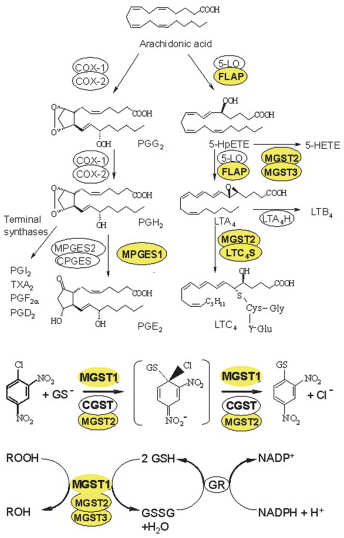
Ces dérivés sont des médiateurs de la douleur, de la fièvre et de l'inflammation. Les protéines MAPEG sont aussi impliquées dans la biotransformation et la détoxification des substances électrophiles. Résumé de la voie de biosynthèse des eicosanoides à partir de l'acide arachidonique :
|
Les protéines MAPEG constituent une superfamille de protéines transmembranaires apparentées sur le plan structural mais avec des fonctions diverses. L'analyse bioinformatique a permis d'établir plusieurs classes des protéines MAPEG des Eucaryotes : (i) les 6 familles de glutathion transférases des microsomes; (ii) la leucotriène C4 synthase; (iii) la protéine d'activation de la 5-lipoxygénase; (iv) la prostaglandine E synthase. Figure ci-dessous : schéma des réactions catalysées par les protéines MAPEG chez les organismes supérieurs.
Source : Hebert & Jegerschold (2007) Abbreviations :
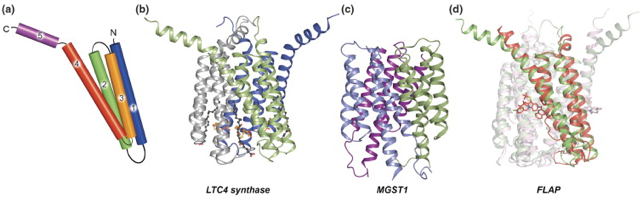
α. Voie de gauche - figure ci-dessus MPGES1 et COX-2 sont induites dans des conditions d'inflammation et catalysent la formation de quantités accrues de PGE2, qui aboutit aux symptômes de l'inflammation comme l'enflure, la douleur et la fièvre. En revanche, COX-1 en relation avec CPGES et MPGES2 assurent un niveau homéostatique de PGE2. La partie soluble de MPGES2 (PDB : 1Z9H) est structuralement similaire aux CGST solubles, mais elle est ancrée dans la membrane de l'appareil de Golgi. Les structures des autres synthases terminales des voies ont été déterminées. β. Voie de droite - figure ci-dessus LTA4 est formé par l'action conjuguée de 5-LO et FLAP. LTC4S catalyse spécifiquement la conjugaison de GSH sur LTA4 formant LTC4 et est impliquée dans des symptômes d'hypersensibilité comme l'asthme et l'allergie. En-dessous : MGST1 fait partie de la Phase II de la détoxification et catalyse la conjugaison de GSH aux substances endogènes électrophiles ou xénobiotiques dans le but d'être excrétées (exemple dans la figure du 1-chloro-2, 4-dinitrobenzène - CDNB). Le complexe central est un intermédiaire transitoire, le complexe Meisenheimer. En bas : réactions de réduction. MGST2 et MGST3 catalysent les réactions indiquées et ont (comme MGST1) une spécificité de substrat et une distribution de tissus relativement large. Elles sont pour l'instant moins bien caractérisées que les autres protéines MAPEG. Figure ci-dessous : structures de certaines protéines MAPEG.
Source : Molina et al. (2008) (a) représentation schématiques de la topologie des hélices transmembranaires. Les séquences en acides aminés de l'hélice transmembranaire 2 sont significativement conservées, en particulier le motif RX(3)NX(2)[D/E]. Quelques caractéristiques structurales des protéines MAPEG :
|
| 12. Liens Internet et références bibliographiques |
|
"Chronological history of lipid science" ("Cyberlipid.org") "An introduction to sphingolipids and membrane rafts" "Membrane proteins of known 3D structure" Base de données "Orientation of Proteins in Membranes" "Docking of Transmembrane Helices into Four Helix Bundles in the High Affinity IgE Receptor" - Zloh et al. CAMPS2.0 (Computational Analysis of the Membrane Protein Space database) "Mechanosensitive Channel MscS" Annotation des séquences de protéines transmembranaires "Prediction of palmitoylation site" |
|
|
Singer & Nicolson (1972) "The fluid mosaic model of the structure of cell membranes" Science 175, 720 - 731 Nicolson G.L. (November 2013) "The Fluid - Mosaic Model of Membrane Structure: Still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years" Biochimica et Biophysica Acta (BBA) - Biomembranes Breukink & Kruijff (2006) "Lipid II as a target for antibiotics" Nat. Rev. Drug Discover. 5, 321 - 323 Pomorski & Menon (2006) "Lipid flippases and their biological functions" Cell. Mol. Life Sci. 63, 2908 - 2921 Sharom (2011) "Flipping and flopping-lipids on the move" IUBMB Life 63, 736 - 746 |
|
|
Jacso et al. (2012) "Characterization of Membrane Proteins in Isolated Native Cellular Membranes by Dynamic Nuclear Polarization Solid-State NMR Spectroscopy without Purification and Reconstitution" Angew. Chem. Int. Ed. Engl. 51, 432 - 435 Schmid & Mettlen (2013) "Cell biology: Lipid switches and traffic control" Nature 499, 161 - 162 Li et al. (2013) "Structure of a presenilin family intramembrane aspartate protease" Nature 493, 56 - 61 |
|
|
Marsh, D. (2008) "Protein modulation of lipids, and vice-versa, in membranes" Biochim. Biophys. Acta - Biomembranes 1778, 1545 - 1575 Shao & Hegde (2011) "Membrane Protein Insertion at the Endoplasmic Reticulum" Annu. Rev. Cell. Dev. Biol. 27, 25 - 56 Wagner & Vorauer-Uhl (2011) "Liposome Technology for Industrial Purposes" J. Drug Deliv. 591325 Hicks et al. (2012) "Lipid rafts and Alzheimer's disease: protein-lipid interactions and perturbation of signaling" Front. Physiol. 3, 189 Akbarzadeh et al. (2013) "Liposome: classification, preparation, and applications" Nanoscale Res. Lett. 8, 102 |
|
|
Fahy et al. (2005) "A comprehensive classification system for lipids" J. Lipids Res. 46, 839 - 862 Niemela et al. (2009) "Bioinformatics and computational methods for lipidomics" J. Chromato. B (Lipidomics: developments and applications) 877, 2855 - 2862 Yang et al. (2010) "Proteome Scale Characterization of Human S-Acylated Proteins in Lipid Raft-enriched and Non-raft Membranes" Mol. Cell Proteomics 9, 54 - 70 Kutateladze (2010) "Translation of the phosphoinositide code by PI effectors" Nat. Chem. Biol. 6, 507 - 513 Paige et al. (2011) "Phospholipase D: Enzymology, Functionality, and Chemical Modulation" Chemical Rev. 111, 6064–6119 |
|
|
Zheng & Jia (2013) "Crystal structure of the integral membrane diacylglycerol kinase" Nature 497, 445 - 446 Li et al. (2013) "Crystal structure of the integral membrane diacylglycerol kinase" Nature 497, 521 - 524 |
|
|
Torres et al. (2002) "Contribution of Energy Values to the Analysis of Global Searching Molecular Dynamics Simulations of Transmembrane Helical Bundles" Biophys J. 82, 3063 - 3071 Vinothkumar & Henderson (2010) "Structures of membrane proteins" Q. Rev. Biophys. 43, 65 - 158 Fiedler et al. (2010) "Protein folding in membranes" Cell. Mol. Life Sci. 67, 1779 - 1798 Psachoulia et al. (2010) "Molecular dynamics simulations of the dimerization of transmembrane alpha-helices" Acc. Chem. Res. 43, 388 - 396 Li et al. (2012) "Transmembrane Helix Dimerization: Beyond the Search for Sequence Motifs" Biochim. Biophys. Acta. 1818, 183 - 193 Cymer et al. (2012) "Transmembrane helix–helix interactions are modulated by the sequence context and by lipid bilayer properties" Biochim. Biophys. Acta (BBA) - Biomembranes 1818, 963 - 973 Zhou & Cross (2013) "Influences of Membrane Mimetic Environments on Membrane Protein Structures" Annu. Rev. Biophys. 42, 361 - 392 |
|
|
Hebert & Jegerschold (2007) "The structure of membrane associated proteins in eicosanoid and glutathione metabolism as determined by electron crystallography" Curr. Opin. Struct. Biol. 17, 396 - 404 Molina et al. (2008) "Catalysis within the lipid bilayer - structure and mechanism of the MAPEG family of integral membrane proteins" Curr. Opin. Struc. Biol. 18, 442 - 449 Sjogren et al. (2013) "Crystal structure of microsomal prostaglandin E2 synthase provides insight into diversity in the MAPEG superfamily" Proc. Natl. Acad. Sci. U S A. 110, 3806 - 3811 M. Whorton (2014) "Structural biology: Calcium-activated proteins visualized" Nature Brunner et al. (2014) "X-ray structure of a calcium-activated TMEM16 lipid scramblase" Nature |
![]()