| Les métabolites secondaires ou produits naturels - Le métabolisme secondaire |
| Tweet |
|
|
1. Présentation générale
2. Aperçu général des voies du métabolisme secondaire 3. Les polycétides ("polyketides") 4. Les polycétides synthétases (PKS) |
5. Mécanisme des polycétides synthétases de type I 6. Les peptides non ribosomiques ("Non-ribosomal peptides") 7. Les synthétases de peptides non ribosomiques ("Non-Ribosomal Peptide Synthetases" - NRPS) 8. Composés aromatiques - Composés phénoliques 9. Liens Internet et références bibliographiques |
1. Présentation générale a. Les métabolites secondaires Les métabolites primaires (métabolisme primaire) sont associés aux fonctions vitales pour la cellule : aucun organisme ne peut vivre sans la glycolyse ou une voie métabolique équivalente. Ensemble de cours sur le métabolisme primaire. Les métabolites secondaires (aussi appelés produits naturels) sont des molécules organiques non directement impliquées dans le développement ou la reproduction d'un organisme. Leur absence n'entraîne pas une mort immédiate mais peut limiter la survie, la fécondité ou l'apparence d'un organisme. Cette absence peut aussi n'avoir aucun effet. Les métabolites secondaires ont essentiellement pour rôle d'accroître la compétitivité de l'organisme qui les biosynthétise : les métabolites secondaires lui procure un avantage sur d'autres organismes. Malgré tout, les métabolites secondaires ont des fonctions biologiques qui peuvent s'avérer essentiels. Exemples:
|
Quelques produits naturels utilisés par l'homme depuis très longtemps Le plus ancien texte médical (Mésopotamie - vers 2600 avant JC) décrit environ 1000 substances d'origine végétale (huiles de cèdre, myrrhe - résine de Commiphora Myrrha, jus de la graine de Papaver somniferum (pavot), ...). Hippocrate de Cos (médecin grec - vers 460-377 avant JC) a recueilli plus de 400 agents naturels et décrit leur utilisation dans son "Corpus Hippocraticum". |
|||
| Canelle | Cinnamomum verum | cinnamaldéhyde, eugenol | phényl-propanoïdes |
| Thé | Camellia sinensis | caféine, théobromine, théophylline | phényl-propanoïdes, terpenes, alcaloïdes |
| Café | Coffea arabica | ||
| Tabac | Nicotiana tabacum | nicotine | alcaloïdes |
| Gingembre | Zingiber officinale | zingerone, gingeroles | sesquiterpenes, monoterpenes |
| Acide acétylsalicylique (aspirine) | Salix alba | --------- | phényl-propanoïdes |
| Pénicilline | Penicillium | --------- | peptide non ribosomique |
| Lovastatine | Pleurotus ostreatus | --------- | polycétide |
| Caroténoïdes | algues, plantes, champignons et bactéries | --------- | isoprenoïdes |
| Source : Ben Field - LGBP - UMR 7264 | |||
|
On peut citer bien d'autres exemples parmis les milliers de produits naturels estimés à ce jour : |
|||
Les métabolites secondaires sont souvent spécifiques d'une espèce. Exemples : les bactéries (Actinomyces, Streptomyces, ...), les champignons filamenteux (Penicillium, ...), les éponges marines (Invertébrés), ... Les plantes produisent plus de 200.000 métabolites secondaires ou spécialisés (produits bio-actifs naturels). |
| Composés phyto-chimiques | ||
| Alcaloïdes | Flavonoïdes | Phényl-propanoïdes |
| Alcaloïdes dérivés de l'ornithine | Flavonoïdes | Monolignols |
| Alcaloïdes dérivés de la lysine | Isoflavonoïdes | Lignanes |
| Alcaloïdes dérivés de l'acide nicotinique | Flavonoïdes complexes | Coumarines |
| Alcaloïdes dérivés de la tyrosine | ||
| Alcaloïdes dérivés du tryptophan et de l'acide anthranilique | Terpenoïdes | Polycétides |
| Alcaloïdes dérivés de l'histidine | Hemiterpenoïdes (C5) | Anthraquinones |
| Alcaloïdes dérivés par réactions d'amination | Monoterpenoïdes (C10) | Pyrones |
| Autres | Sesquiterpenoïdes (C15) | Autres |
| Diterpenoïdes (C20) | ||
| Skimate / composés issus de la voie de l'acétate - malonate | Sesterterpenoïdes (C25) | Autres |
| Stilbenoïdes | Triterpenoïdes (C30) | Naphthoquinones |
| Autres | Stéroïdes | Tannins et dérivés galloyl |
| Caroténoïdes et apo-caroténoïdes | ||
| Composés dérivés d'acides aminés | Autres | |
| Betalaines | ||
| Glucosides cyanogèniques | ||
| Glucosinolates | ||
Exemples de précurseurs de produits industriels ou pharmaceutiques synthétisés par des micro-organismes modifiés
|
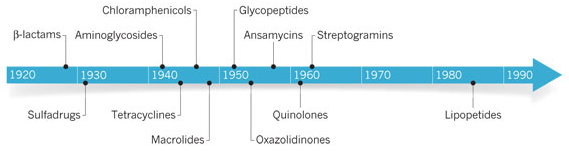
b. Recherche et découverte d'antibiotiques et d'antifongiques L'utilisation généralisée d'antibiotiques à partir des années 1940 (notamment la pénicilline - découverte en 1928 et la streptomycine - découverte en 1943) a transformé la médecine en fournissant des remèdes efficaces contre les maladies les plus fréquentes de l'époque. Les antibiotiques naturels ont évolué afin de rompre les barrières qui empêchent la pénétration dans les bactéries cibles et la plupart des antibiotiques introduits en médecine ont été découverts à partir de microorganismes du sol cultivables. Cependant, le développement de résistances limite la durée de vie utile des antibiotiques et exige de découvrir en permanence de nouveaux composés. Mais la découverte de médicaments anti-microbiens est d'une rare difficulté, principalement en raison de la faible pénétration des composés dans les bactéries. La surexploitation de cette ressource limitée dans les années 1960 a mis fin à la découverte d'antibiotiques et ce d'autant que les approches par synthèse chimique n'ont pas pu remplacer les antibiotiques naturels.
Source : Lewis K. (2012) |
| Classe chimique | Cible | Mode d'action | Exemples |
| Sulfonamides | synthèse du folate | bactériostatique | Sulfanilamide |
| β-Lactames | synthèse de la paroi | bactéricide | Penicilline Céphalosporine Carbapenemes |
| Aminoglycosides | synthèse protéique | bactéricide | Spectinomycine Kanamycine Neomycine |
| Tétracycline | synthèse protéique | bactériostatique | Tétracycline Doxycycline |
| Chloramphenicol | synthèse protéique | bactériostatique | Chloramphenicol |
| Macrolides | synthèse protéique | bactériostatique | Erythromycine Clarithromycine |
| Glycopeptides | synthèse de la paroi | bactéricide | Vancomycine Teicoplanine |
| Oxazolidinones | synthèse protéique | bactériostatique | Linezolide |
| Ansamycines | synthèse des ARN | bactéricide | Rifamycine |
| Quinolones | synthèse de l'ADN | bactéricide | Ciprofloxacine |
| Streptogramines | synthèse protéique | bactéricide | Pristinamycine |
Début 2015, des chercheurs ont annoncé que la teixobactine (isolée de la bactérie Eleftheria terrae) est le premier membre d'une nouvelle classe d'inhibiteurs de la synthèse de peptidoglycanes.
Source : Ling et al. (2015) La teixobactine se fixe à la région undecaprenyle-PP-sucre de deux cibles précurseurs différentes : le lipide II (précurseur du peptidoglycane) et le lipide III (précurseur de l'acide téichoïque). La teixobactine est très puissante contre une large gamme de bactéries Gram-positif, y compris Staphylococcus aureus résistant à la méthicilline et les entérocoques résistants à la vancomycine.
Source : Ling et al. (2015)
|
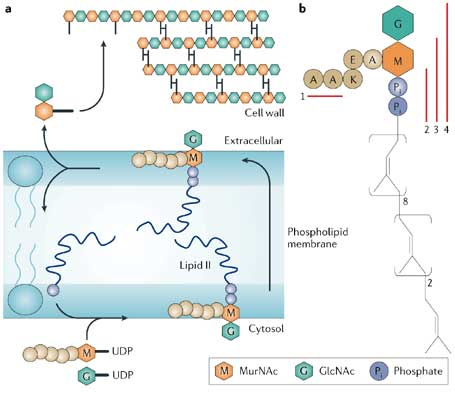
La paroi de toutes les bactéries contient un polymère de sucres aminés alternés : la N-acétylglucosamine (GlcNAc) et l'acide N-acétylmuramique (MurNAc) - figure ci-dessous - partie a en haut. Ces polymères de glycanes sont réticulées par un pentapeptide dont la séquence est généralement [L-Ala-γ-D-Glu-diaminopimelyl (ou L-Lys) -D-Ala-D-Ala] attaché au sucre MurNAc. Cette réticulation confère à la cellule sa rigidité et sa résistance mécanique.
Source : Breukink & Kruijff (2006) Le lipide II (plusieurs milliers de molécules par cellule bactérienne) est le transporteur d'isoprénoides C55 qui reçoit le muramyl-pentapeptide à la fin de la phase qui a lieu dans le cytoplasme de la biosynthèse du peptidoglycane (figure ci-dessus - partie a en bas ). Tout en restant sur la face interne de la membrane cytoplasmique, le muramyl-pentapeptidyl-lipide II est glycosylé. Puis le disaccharyl-pentapeptidyl-lipide II est transloqué par des flippases sur la face externe de la membrane cytoplasmique. Lorsqu'elle affleure à la surface externe de la membrane, la partie lipide II du disaccharyl-pentapeptidyl-lipide II est la cible de 2 types d'antibiotiques :
|
Nouveaux antibiotiques contre les bactéries Gram- La darobactine est un nouvel antibiotique produit par un opéron silencieux de Photorhabdus khanii (synthèse ribosomale). Il semble actif contre les bactéries Gram- in vitro (exemples : Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginos, Acinetobacter baumannii, …) et chez des modèles d'infection animale.
Une famille d'antibiotiques peptido-mimétiques chimériques (murepavadine et polymyxine B1) qui possèdent une activité antimicrobienne à large spectre contre les bactéries à Gram- a également été découverte. Ces antibiotiques ont un mécanisme d'action qui implique également la liaison au LPS et à BamA. Voir Imai et al. (2019) & Luther et al. (2019). |
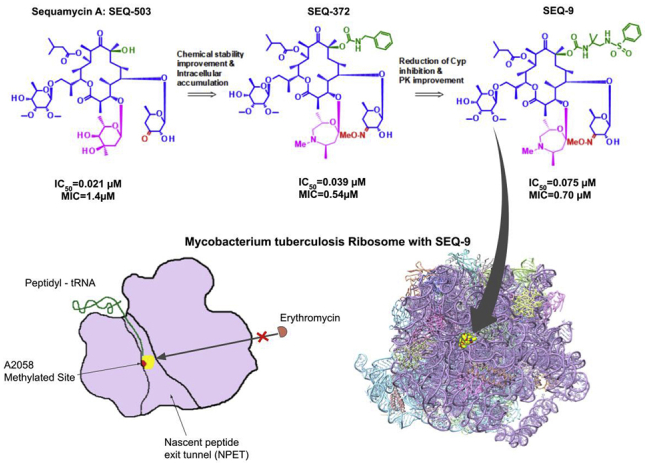
Nouvel antibiotique contre une bactérie Gram+ Des macrolides appelés séquanamycines sont des inhibiteurs exceptionnels (in vitro et in vivo) du ribosome de Mycobacterium tuberculosis. Les séquanamycines interagissent avec le ribosome de la même manière que l'érythromycine et la clarithromycine (des macrolides également). Cependant, les caractéristiques de liaison des séquanamycines surmontent la résistance aux macrolides de Mycobacterium tuberculosis. L'obtention de structures tridimensionnelles des séquanamycines fixés au ribosome ont permis de les optimiser et d'obtenir une molécule appelée SEQ-9 :
Source : Zhang et al. (2023) |
Recherche de nouveaux antibiotiques par intelligence artificielle Un réseau de neurones a été créé pour apprendre à reconnaître les particularités structurales des molécules qui inhibent la croissance d'Escherichia coli en utilisant une collection de 2335 molécules à activité anti-bactérienne connue, dont environ 300 antibiotiques et 800 produits naturels d'origine végétale, animale et microbienne (Stokes et al., 2020). Le modèle construit a ensuite été utilisé pour analyser plus de 100 millions de molécules de la bibliothèque "Drug Repurposing Hub" (entre autres molécules, cette bibliothèque contient environ 6000 molécules à l'étude pour des maladies chez l'homme) : environ 100 molécules candidates ont été sélectionnées pour des tests. Une molécule (étudiée pour un éventuel traitement du diabète) s'est avérée un antibiotique puissant : l'halicine ("c-Jun N-terminal kinase inhibitor", SU3327), dénommée ainsi d'après HAL (l'ordinateur du film "2001: A Space Odyssey").
Puis plus de 107 millions de structures de molécules de la base de données ZINC15 (base de données gratuite de près d'un milliard de composés disponibles dans le commerce pour le criblage virtuel) ont été analysées : 23 molécules candidates ont été testées et 8 molécules ont une une activité antibactérienne. En particulier, deux d'entre elles ont une activité contre un large éventail d'agents pathogènes et contre des souches d'Escherichia coli résistantes aux antibiotiques. |
Recherche de nouveaux antifongiques par combinaison d'algorithme et d'analyse bioinformatique de génomes Le décryptage de la voie des stérols a permis de développer des médicaments tels que les statines hypo-cholestérolémiantes et les azoles antifongiques.
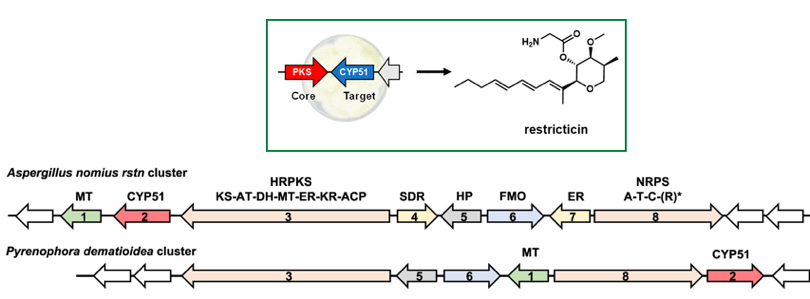
Penicillium restrictum fabrique la restricticine pour se défendre contre d'autres champignons (exemple, Candida albicans). Cet antifongique bloque une enzyme cruciale de ces champignons adverses et freine ainsi leur croissance. Tout champignon qui fabrique de la restricticine doit se protéger de cette molécule : le gène fongique qui code cet antifongique se trouve à proximité du gène codant un variant de son enzyme cible résistant à la restricticine. Le développement d'un algorithme d'exploration du génome pour rechercher des gènes codant pour une enzyme résistante à proximité de groupes de gènes susceptibles de synthétiser la restricticine a permis de découvrir la voie de biosynthèse de cet antifongique (synthétisé également chez d'autres espèces de champignons).
Source : Liu et al. (2021) |
c. Classifications des métabolites secondaires On estime à plusieurs centaines de milliers les métabolites secondaires, de structure et de fonction très diverses. Il existe donc un grand nombre de classification selon les sources. Il ressort que la classification la plus élémentaire des métabolites secondaires inclue trois groupes :
L'une des classifications les plus fiables et exhaustives est celle de KEGG qui s'appuie sur de très nombreuses informations (en particulier les voies de biosynthèse) concernant les produits naturels. |
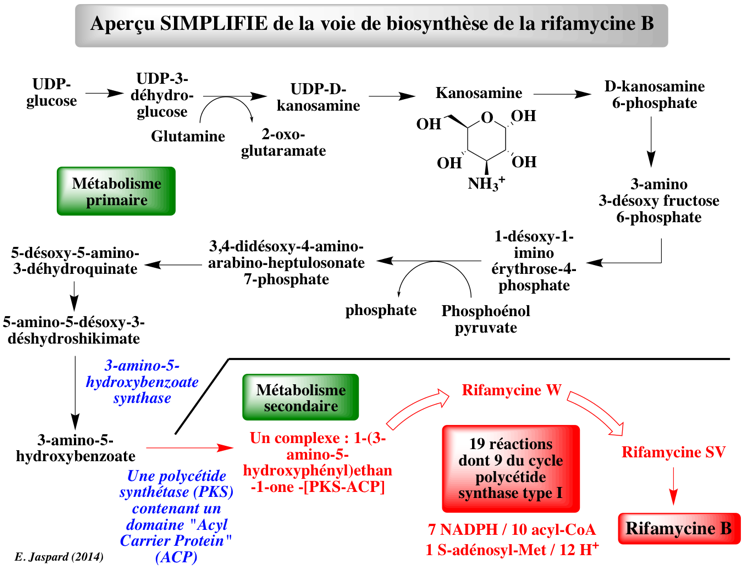
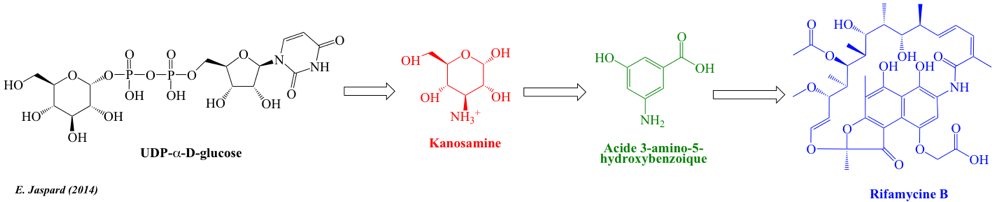
2. Aperçu général des voies du métabolisme secondaire En terme de complexité le métabolisme secondaire n'a de "secondaire" que le nom. Et ce d'autant plus que l'ensemble des molécules précurseurs du métabolisme secondaire sont issues du métabolisme primaire. Figure ci-dessous : exemple simplifié de l'extrême complexité de la synthèse d'un métabolite secondaire. Les enzymes et la plupart des molécules connexes aux réactions ne sont pas indiquées.
La distinction métabolisme primaire versus métabolisme secondaire n'est pas formelle. La partie métabolisme secondaire implique un très grand nombre de réactions. Notamment celles catalysées par le complexe multi-fonctionnel polycétide synthase : construction cyclique de l'armature pour la fin de la synthèse de la rifamycine B. Le plus étonnant est la transformation des métabolites du point de vue structural et donc chimique :
Les rifamycines (classification "Ansamycins and related polyketides") forment un groupe d'antibiotiques synthétisés par la bactérie Amycolatopsis rifamycinica. Ils sont une sous-classe de la famille des ansamycines. |
Voies du métabolisme secondaire Voir la carte générale du métabolisme secondaire. Les liens dans le tableau ci-dessous renvoient vers des fichiers ou des cartes interactives du métabolisme (exemple ci-dessous).
Ces liens permettent d'obtenir des informations sur les métabolites, les enzymes et les gènes codant ces enzymes qui sont impliqués dans chaque réaction d'une voie métabolique considérée. |
| Biosynthesis of secondary metabolites | Biosynthesis of other secondary metabolites | |
|
00940 Phenylpropanoid biosynthesis M00039 Monolignol biosynthesis M00137 Flavanone biosynthesis M00138 Flavonoid biosynthesis M00369 Cyanogenic glycoside biosynthesis M00370 Glucosinolate biosynthesis M00661 Paspaline biosynthesis |
00945 Stilbenoid, diarylheptanoid and gingerol biosynthesis 00941 Flavonoid biosynthesis 00944 Flavone and flavonol biosynthesis 00942 Anthocyanin biosynthesis 00943 Isoflavonoid biosynthesis 00901 Indole alkaloid biosynthesis 00403 Indole diterpene alkaloid biosynthesis 00950 Isoquinoline alkaloid biosynthesis 00960 Tropane, piperidine and pyridine alkaloid biosynthesis 01058 Acridone alkaloid biosynthesis |
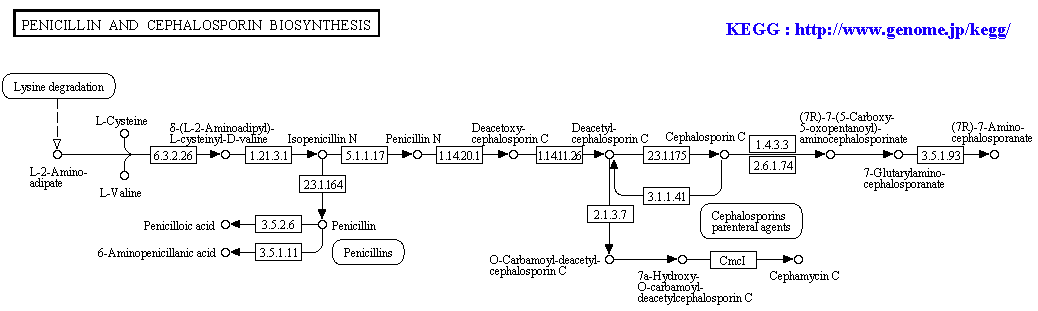
00232 Caffeine metabolism 00965 Betalain biosynthesis 00966 Glucosinolate biosynthesis 00402 Benzoxazinoid biosynthesis 00311 Penicillin and cephalosporin biosynthesis 00332 Carbapenem biosynthesis 00331 Clavulanic acid biosynthesis 00521 Streptomycin biosynthesis 00524 Butirosin and neomycin biosynthesis 00231 Puromycin biosynthesis 00401 Novobiocin biosynthesis 00254 Aflatoxin biosynthesis |
| Terpenoid backbone biosynthesis | Other terpenoid biosynthesis | Metabolism of terpenoids and polyketides |
|
M00095 C5 isoprenoid biosynthesis, mevalonate pathway M00096 C5 isoprenoid biosynthesis, non-mevalonate pathway M00364 C10-C20 isoprenoid biosynthesis, bacteria M00365 C10-C20 isoprenoid biosynthesis, archaea M00366 C10-C20 isoprenoid biosynthesis, plants M00367 C10-C20 isoprenoid biosynthesis, non-plant eukaryotes |
M00097 beta-Carotene biosynthesis M00371 Castasterone biosynthesis |
01052 Type I polyketide structures 00522 Biosynthesis of 12-, 14- and 16-membered macrolides 01051 Biosynthesis of ansamycins 01056 Biosynthesis of type II polyketide backbone 01057 Biosynthesis of type II polyketide products 00253 Tetracycline biosynthesis 00523 Polyketide sugar unit biosynthesis 01054 Nonribosomal peptide structures 01053 Biosynthesis of siderophore group nonribosomal peptides 01055 Biosynthesis of vancomycin group antibiotics |
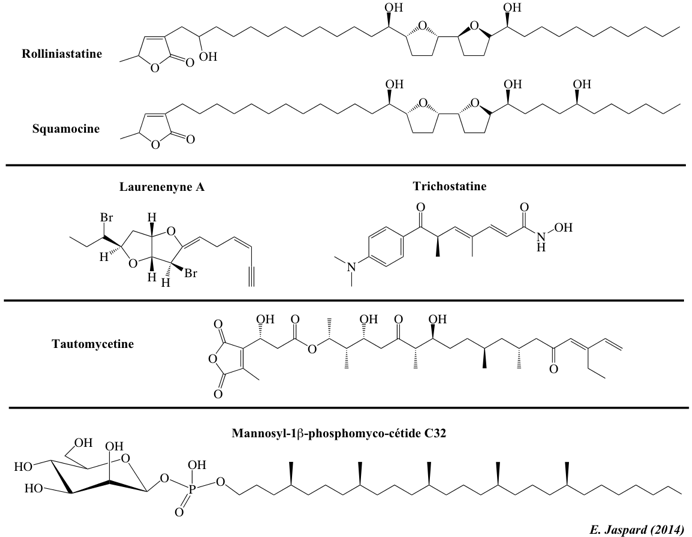
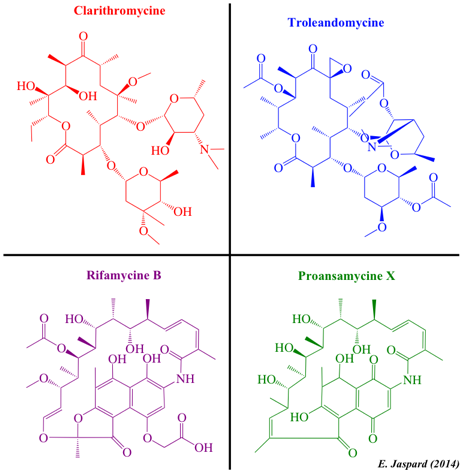
| Quelques polycétides - les liens renvoient vers les données physico-chimiques de la base de données PubChem | ||
| Acetogenines et polycétides aromatiques | Polycétides linéaires | Polycétides aromatiques et dérivés d'ansamycines |
| Rolliniastatine-1 | Trichostatine | Laurenenyne A |
| Squamocine | Tautomycetine | Rifamycine B |
| Hypericine | Mannosyl-1β-phosphomycoketide C30 à 34 | Pro-ansamycine X |
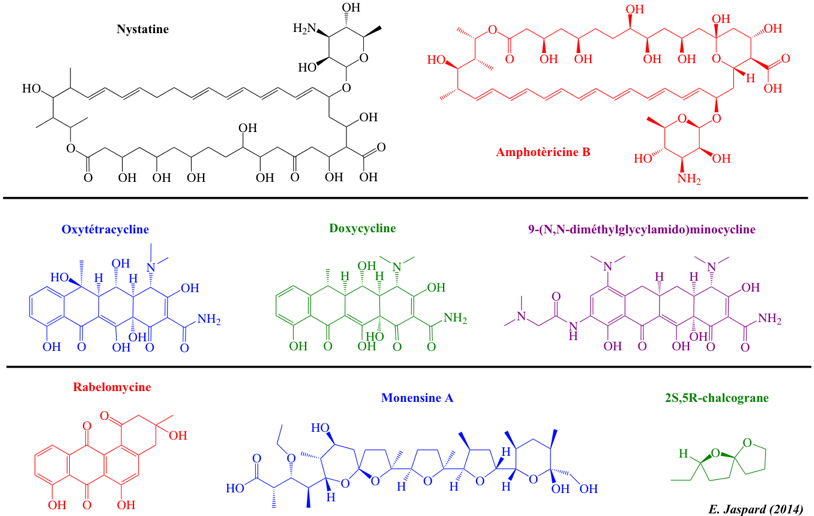
| Flavonoïdes | Polyènes et tétracyclines linéaires | Angucyclines et polycétides poly-éther |
| Cyanidine | Nystatine | Rabelomycine |
| Pelargonidine | Monensine A | |
| Petunidine | Oxytétracycline | 2S,5R-chalcograne |
| Rotenone | Doxycycline | |
| Folinine | 9-(N,N-diméthylglycylamido)minocycline | |
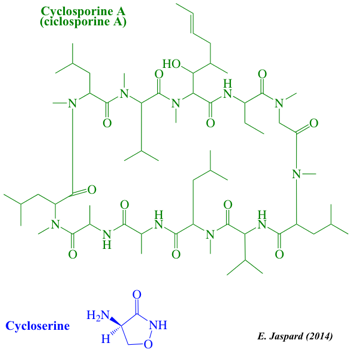
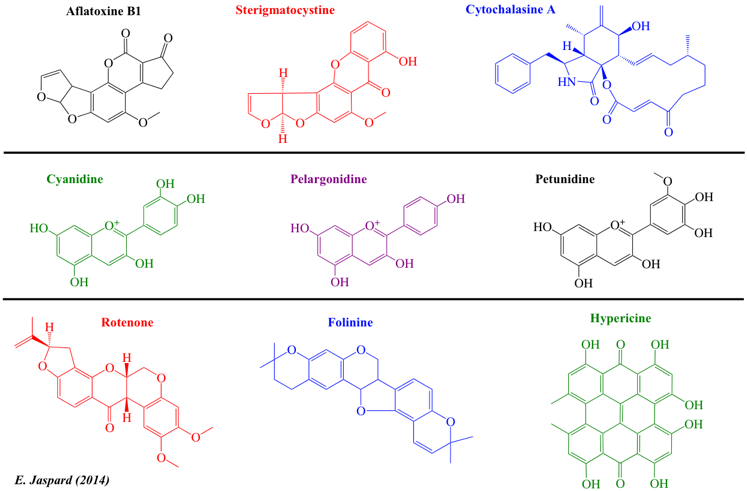
| Peptides non ribosomiques (polycétides hybrides) | Aflatoxines et cytochalasines | Macrolides et polycétides lactone |
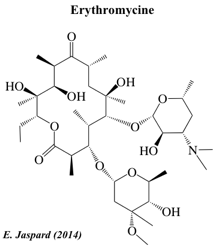
| Ciclosporine A | Aflatoxine B1 | Erythromycine |
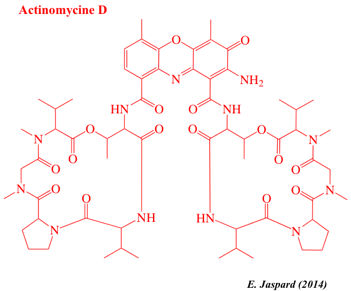
| Actinomycine D | Sterigmatocystine | Clarithromycine |
| Cycloserine | Cytochalasine A | Troleandomycine |
| Source : LIPID Metabolites and Pathways Strategy (LIPID MAPS) | ||
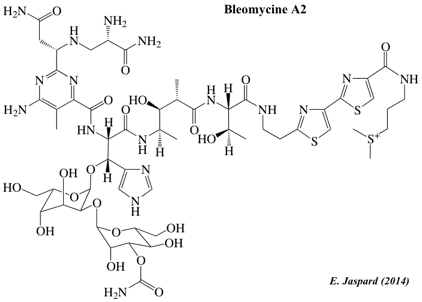
| Bleomycine A2 | ||
Les polycétides forment une grande famille de produits naturels (métabolites secondaires) synthétisés par les bactéries, les champignons et les plantes. Il existe une remarquable diversité de polycétides, tant en termes de structures que de fonctions. Ils possèdent une très large gamme d'activités pharmacologiques (antimicrobiens, antifongiques, antiparasitaires, anti-tumoraux) et agrochimiques.
Source des fichiers cdxml : LIPID Metabolites and Pathways Strategy (LIPID MAPS) En particulier, les polycétides incluent de nombreux médicaments importants tels que :
|
Les cyclosporines sont un groupe de macrolides isolés à partir de champignons. Classification : "Polyketides : Non-ribosomal peptide/polyketide hybrids".
La ciclosporine A a été initialement isolée à partir de Tolypocladium inflatum (Hans Frey - 1969 - Norvège). C'est un peptide non ribosomique cyclique de 11 acides aminés (undécapeptide). Il contient un acide aminé de configuration D (dextrogyre) rarement rencontré dans la nature. La ciclosporine A est un immunosuppresseur (J.F. Borel - 1976) puissant avec une action spécifique sur les lymphocytes T. L'érythromycine est un antibiotique utile dans le traitement d'un certain nombre d'infections bactériennes. L'érythromycine a un spectre antimicrobien à peu près semblable à celui de la pénicilline. Classification : "Macrolides and lactone polyketides".
L'actinomycine D (ou dactinomycine) est le plus important membre des actinomycines, classe d'antibiotiques polypeptidiques isolés de bactéries du genre Streptomyces. L'actinomycine D inhibe la transcription en se fixant au complexe d'initiation de la transcription, empêchant ainsi l'élongation de la chaîne d'ARN par l'ARN polymérase. Classification : "Non-ribosomal peptide/polyketide hybrids".
La bléomycine est un antibiotique polypeptidique isolé de bactéries du genre Streptomyces. La bléomycine rompt les brins d'ADN. La bléomycine méga-synthétase est constituée d'une synthétase de peptides non ribosomiques et de modules de polycétide synthétase de type I (voir ci-dessous). Voir : K.M. Fisch (2013). Classification : "Non-ribosomal peptide/polyketide hybrids".
|
Doxycycline (dérivé de l'oxytétracycline - Streptomyces rimosus) : antibiotique. Monensine A (Streptomyces cinnamonensis) : antibiotique (ionophore). |
Source des fichiers cdxml : LIPID Metabolites and Pathways Strategy (LIPID MAPS) |
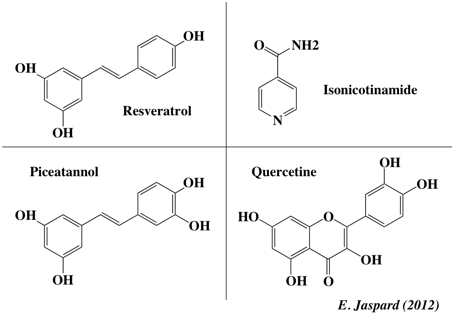
Le resvératrol (3,4′,5-trihydroxy-trans-stilbene) est un polycétide aromatique. C'est une phytoalexine (polyphénols de la classe des stilbènes) que l'on trouve dans de nombreuses plantes supérieures (notamment le raisin, l'arachide, la rhubarbe, …). Il augmente l'expression d'un microARN (miR-663) dans les cellules immunitaires humaines, diminuant ainsi l'expression du microARN miR-155 qui favorise la réaction inflammatoire. Figure ci-dessous : différents activateurs de la sirtuine 1.
Le resvératrol a un effet cardioprotecteur en inhibant l'agrégation des plaquettes et en atténuant l'expression de la P-selectine 35. Cet effet serait lié à la diminution de l'activité de la phospholipase C. Le resvératrol neutralise les radicaux libres (et autres oxydants) et inhibe l'oxydation des lipoprotéines LDL. |
4. Les polycétides synthétases (PKS) Les polycétides sont biosynthétisés à partir de précurseurs acyl-CoA par les polycétides synthétases ("Polyketides Synthases" - PKS). L'étape clé de leur biosynthèse est une condensation décarboxylante analogue à l'étape d'allongement des chaînes au cours de la biosynthèse des acides gras. Cependant, contrairement à la biosynthèse des acides gras au cours de laquelle chaque étape d'élongation de la chaîne est suivie d'une séquence fixe de réactions (céto-réduction, déshydratation et réduction du groupe énoyle), les différents intermédiaires de l'allongement des chaînes des polycétides subissent toutes, certaines ou aucune de ces modifications de groupes fonctionnels. Il en résulte un degré de complexité chimique remarquable de ces produits naturels. Un degré de complexité supplémentaire est atteint du fait :
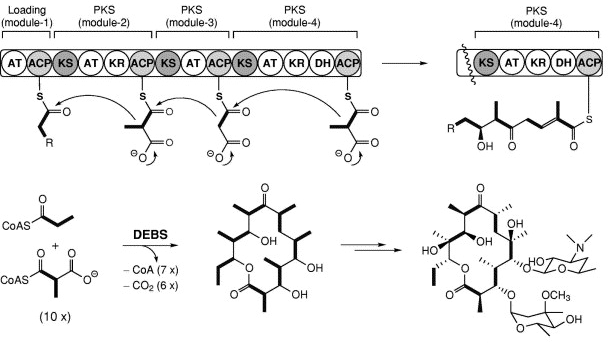
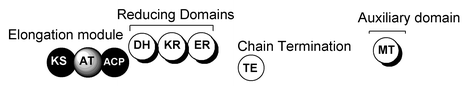
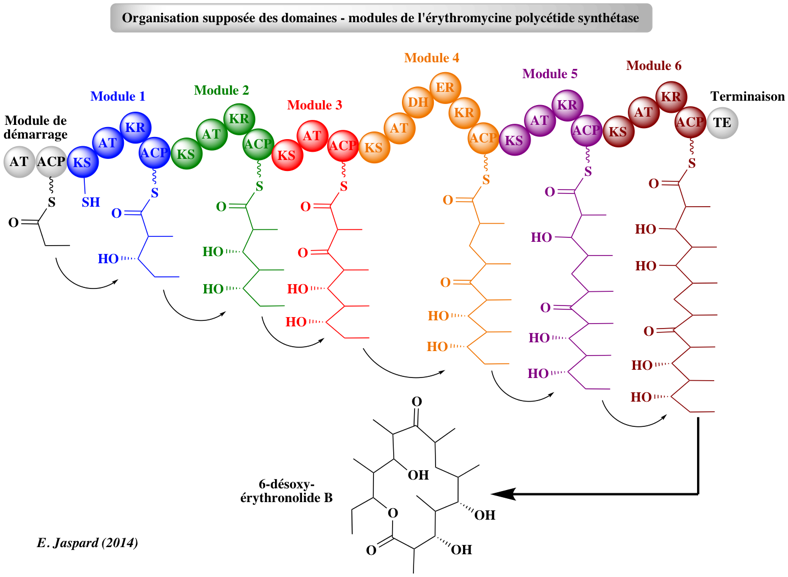
Les 3 types de PKS Les PKS de type I sont des enzymes multi-fonctionnelles organisées en modules : chaque module assure un ensemble distinct d'activités enzymatiques non-itératives responsables de la catalyse d'un cycle d'allongement des chaînes des polycétides. Elles utilisent la protéine "Acyl Carrier Protein" (ACP) pour activer les substrats acyl-CoA et canaliser la croissance des chaînes de polycétides.
Source : B. Shen (2003) Exemple : la 6-désoxy-érythromycine B synthétase ("6-deoxyerythromycin B synthase" - DEBS) pour la biosynthèse de polycétides réduits (c'est à dire les macrolides, les polyéthers et le polyène) tels que l'érythromycine A. Les acronymes des domaines : AT : Acyl-Transferase / ACP : Acyl Carrier Protein / KS : Keto-Synthase / KR : Keto-Reductase / DH : DeHydratase Les PKS de type II (aussi appelées "bacterial aromatic PKS") sont des complexes multi-enzymatiques qui catalysent de manière itérative un ensemble unique de réactions.
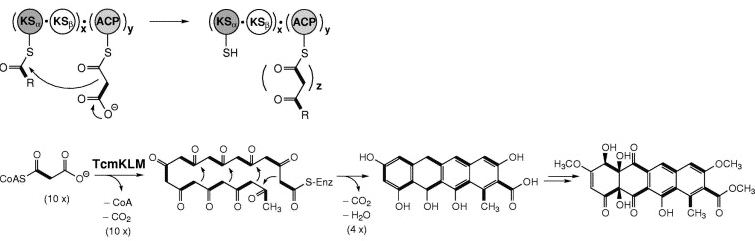
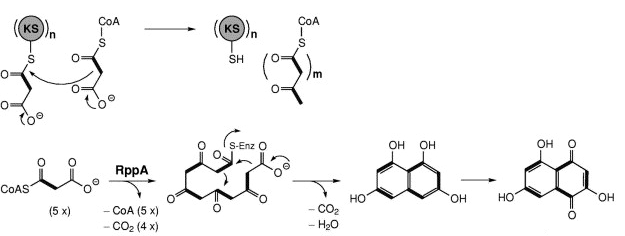
Source : B. Shen (2003) Les domaines minimaux requis des PKS de type II (mais aussi des PKS de type I) pour effectuer un tour de condensation décarboxylante sont : KS - AT - ACP. Elles contiennent des domaines catalytiques similaires à ceux des PKS de type I, à ceci près qu'il y a deux domaines KS : KSα et KSβ. Le premier est équivalent au domaine KS des PKS de type I. Le second contrôle la longueur du polycétide. La réduction des groupes β-cétone n'a lieu qu'après la synthèse complète du polycétide. Elles sont impliquées dans la synthèse des polycétides aromatiques (souvent polycycliques). Exemples : la tétracénomycine PKS pour la biosynthèse de la tétracénomycine C - les PKS impliquées dans la synthèse de la partie aglycone de l'actinorhodine et de la daunorubicine. Les PKS de type III (aussi appelées "chalcone synthase-like PKS") sont des enzymes homodimèriques.
Source : B. Shen (2003) Comme les PKS de type I et de type II, les PKS de type III (exemple : la chalcone synthase) condensent de manière itérative une unité de démarrage avec une série d'unités d'extension pour générer une chaîne poly-β-cétone. Mais, à la différence des deux autres types, les PKS de type III :
Exemples :
Malgré des différences structurales et mécanistiques, les 3 types de PKS catalysent la biosynthèse des polycétides par condensation décarboxylante séquentielle des précurseurs acyl-CoA. Le domaine céto-acyl synthétase (voir ci-dessous) des PKS de type I ou la sous-unité céto-acyl synthétase des PKS de type II et de type III catalysent l'étape de formation de la liaison C-C. |
5. Mécanisme des polycétides synthétases de type I Les PKS de type I sont des enzymes multi-fonctionnelles organisées en modules séparés par de courtes régions. L'ordre des modules de l'extrémité N-terminale vers C-terminale est :
Source : Salomon et al. (2004) Les acronymes des domaines: KS : Keto-Synthase / AT : Acyl-Transferase / ACP : Acyl Carrier Protein La chaîne de polycétide et les groupes de démarrage sont liés via leur groupe fonctionnel carboxyle au groupe SH de l'ACP et de KS (liaison thioester).
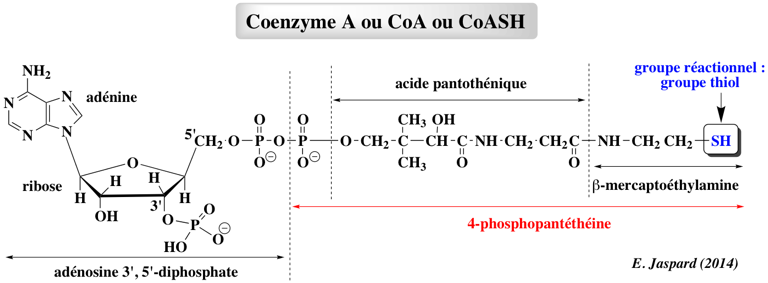
La chaîne en croissance est transférée d'un groupe thiol à un autre par trans-acylation et elle est libérée par hydrolyse ou par cyclisation. Les différents intermédiaires de l'allongement des chaînes des polycétides subissent toutes, certaines ou aucune de ces modifications de groupes fonctionnels. Le coenzyme A ou CoA ou CoASH est la molécule qui permet les réactions de transfert des groupes acyles (R-C=O). Ces groupes sont liés au coenzyme A par des liaisons thioester, liaisons à haut potentiel énergétique (ΔG°' = - 9 kcal/mol).
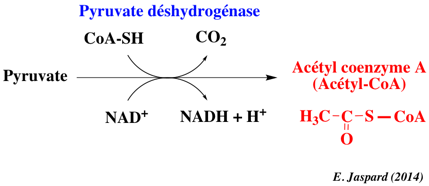
Le coenzyme A est un dérivé de l'acide pantoténique, vitamine de la famille des vitamines B. Remarque : l'adénosine 3', 5' -diphosphate n'est pas à confondre avec l'ADP = adénosine 5' -diphosphate. Le complexe multi-enzymatique (et multi co-facteurs) de la pyruvate déshydrogénase (ou pyruvate:NADP+ oxidoréductase ou PDH - EC 1.2.1.51) catalyse la transformation du pyruvate en acétyl-CoA.
|
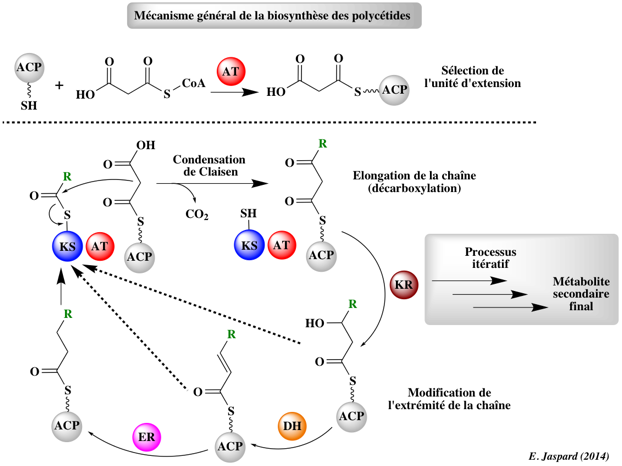
Phase de démarrage Le groupe de démarrage, généralement l'acétyl-CoA ou le malonyl-CoA, est chargé sur le domaine ACP du module de démarrage.
Cette réaction est catalysée par le domaine AT (Acyl-Transferase) du module de démarrage. Etapes d'allongement de la chaîne
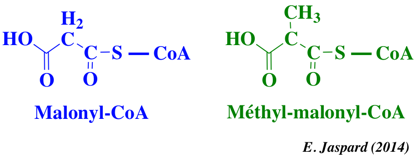
Figure adaptée de Staunton & Weissman (2001) La chaîne de polycétide est transférée du domaine ACP du module précédent au domaine KS (Keto-Synthase) du module en cours. Cette réaction est catalysée par le domaine KS. Le groupe d'élongation, généralement le malonyl-CoA ou le méthyl-malonyl-CoA, est chargé sur le domaine ACP du module en cours. Cette réaction est catalysée par le domaine AT (Acyl-Transferase) en cours. Le groupe d'allongement lié à l'ACP réagit via une condensation de Claisen avec la chaîne de polycétide fixée au domaine KS (dégagement de CO2) : le domaine KS est libéré et la chaîne de polycétide en cours d'allongement est fixée à l'ACP. La réaction a lieu à l'extrémité de la chaîne fixée au domaine KS : la chaîne se déplace d'une position et le groupe d'allongement devient le nouveau groupe fixé. Eventuellement, la chaîne de polycétide en cours d'allongement peut être modifiée pas à pas par des domaines supplémentaires :
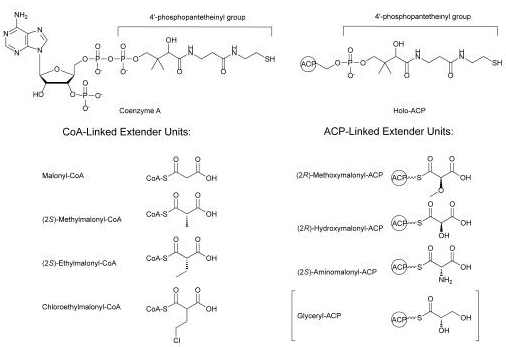
Ce cycle est répété pour chaque module d'allongement. Etape de terminaison Le domaine TE (Thio-Esterase) hydrolyse la chaîne de polycétide complète du domaine ACP du module précédent. Unités d'extension ("extender units") Les unités d'extension les plus couramment sélectionnées sont celles liées au CoA : malonyl-CoA, (2S)-méthylmalonyl-CoA, (2S)-éthylmalonyl-CoA et chloroéthylmalonyl-CoA.
Source : Chan & Thomas (2010) Les unités d'extension liées à l'ACP sont moins courantes : (2R)-méthoxymalonyl-ACP, (2R)-hydroxymalonyl-ACP et (2S)-aminomalonyl-ACP. Elles sont synthétisées sur le bras phosphopantéthéinyl d'une ACP séparée de la ligne d'assemblage PKS. La protéine porteuse d'acyle ("Acyl Carrier Protein " - ACP) L'ACP intervient dans la biosynthèse des acides gras et des polycétides. La chaîne en cours de croissance est liée sous forme d'un thiol-ester au groupement thiol distal de la 4'-phosphopantéthéine. L'ACP est biosynthètisée sous forme apo inactive. La 4'-phosphopantéthéine est ajoutée de manière post-traductionnelle à la sérine 36 conservée de l'ACP, réaction catalysée par la protéine holo-ACP synthase (transférase de groupe phosphopantéthéinyle). CoA-(4'-phosphopantéthéine) + apo-[ACP] ===> adénosine 3',5'-bis-phosphate + holo-[ACP] |
|
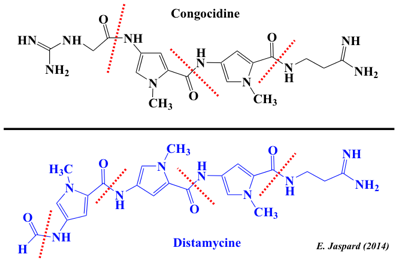
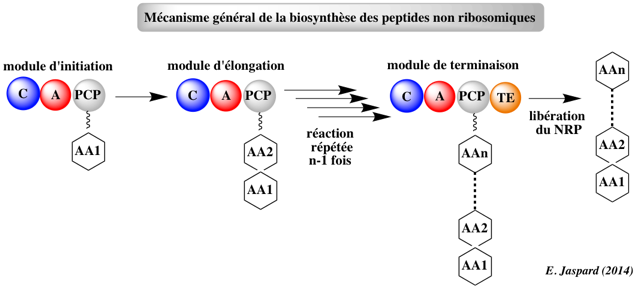
6. Les peptides non ribosomiques ("Non-ribosomal peptides" - NRP) Les peptides non ribosomiques (NRP) sont une classe majeure de métabolites secondaires peptididiques, généralement synthétisés par des bactéries et des champignons. Ils sont synthétisés par des synthétases de peptides non ribosomiques ("non-ribosomal peptide synthetases") : cette synthèse est donc indépendante d'un ARN messager. Chaque synthétase ne peut synthétiser qu'un seul type de peptide. Remarque : certaines cyanobactéries biosynthétisent des peptides avec des ribosomes et les modifient post-traductionnellement (exemples : NRP de la famille des patellamides, NRP de la famille des microviridines). Exemples de NRP Les pyrrolamides (congocidine, distamycine, anthramycine, kikumycines, pyrronamycines, noformycine, ...) constituent une famille de NRP synthétisés par Streptomyces ou d'autres actinobactéries (par exemple, Streptomyces ambofaciens pour la production industrielle de la spiramycine). Cette famille de pyrrole-amides (aussi appelées oligopyrroles ou oligopeptides) contient un ou plusieurs groupes pyrrole-2-carboxamides. Ils se fixent de manière réversible sur des séquences spécifiques (quatre ou plus paires [A-T] consécutives) du petit sillon de la double hélice d'ADN.
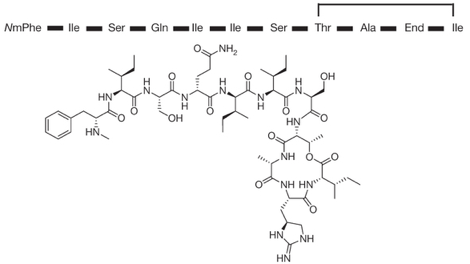
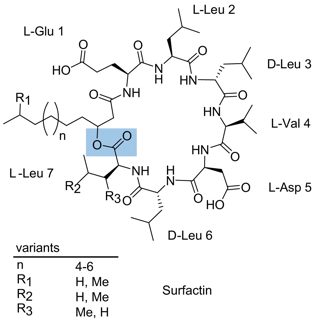
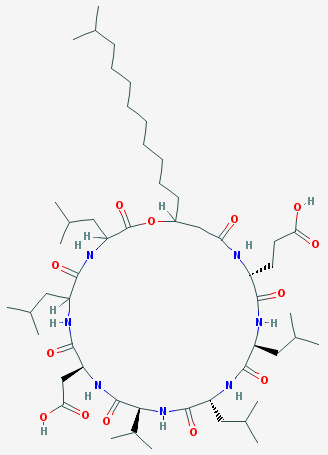
La surfactine (synthétisée par Bacillus subtilis) appartient à la classe des lipopeptides. Elle est couramment utilisée comme antibiotique. C'est aussi un agent tensio-actif très puissant utilisé dans certaines applications biotechnologiques. Sa bioactivité est fortement influencée par la partie acides gras. La surfactine est une lipo-heptapeptide lactone cyclique qui contient 2 acides aminés acides (Glu et Asp) à côté de 5 acides aminés non polaires et 1 acide gras 3-hydroxy. Il existe plusieurs variants de la surfactine. La liaison lactone est surlignée en bleu.
Source : Kraas et al. (2010) La chaîne de la partie acide gras est variable en longueur (13 à 15 carbones) et en ramification de la chaîne.
Source : PubChem |
Autres exemples de NRP Dipeptides cycliques, indigoidine, andrimide, moiramide, salinosporamide K, orphamide A, solonamides et dérivés, holomycine, skyllamycine, holomycine, saframycine, laspartomycine, thiomarinols, ... Les depsipeptides sont des peptides au sein desquels un ou plusieurs groupe(s) amide [-C(O)NH-R] est/sont remplacé(s) par un groupe ester [-C(O)O-R]. Exemples :
NRP biosynthétisés par les cyanobactéries :
|
| Module | Domaine | Activité enzymatique | |
| d'initiation | Ce module ne possède pas, en général, de domaine C | ------------ | |
| d'élongation | d'adénylation (A) | sélection et activation des acides aminés | |
de thiolation (T) |
maintient la structure intermédiaire et la transmet au domaine suivant (C) contient le co-facteur 4'-phospho-pantéthéine |
||
de condensation (C) |
formation de la liaison peptidique conjointement avec le domaine T | ||
| thioestérase (TE) | libération des NRP | ||
optionnels |
méthyltransférase (MT) |
N-méthylation | |
| de formylation (F) | formylation | ||
| d'épimérisation (E) | conversion de l'acide aminé chargé en son énantiomère D | ||
| d'oxydation (Ox) | oxydation de thiazolines ou oxazolines en thiazoles ou oxazoles | ||
| de réduction (Red) | réduction de thiazolines ou oxazolines en thiazolidines ou oxazolidines | ||
| de réduction (R) | réduction en aldéhyde ou alcool | ||
| de terminaison | possède, en général, un domaine TE | ------------ | |
Quelques structures de domaines de NRPS : module de terminaison ("surfactin synthetase" - 2007) : 2VSQ / bidomaines PCP-C ("tyrocidin synthetase" - 2007) : 2JGP / domaine réductase (2012) : 4DQV / domaine de condensation ("CDA peptide synthetase" - 2013) : 4JN5 |
|||
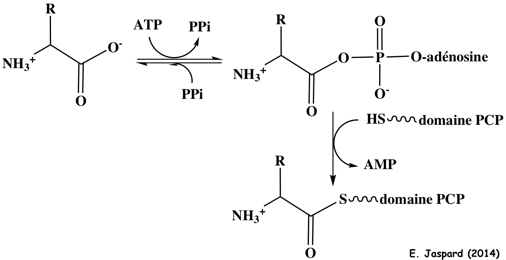
Le domaine A (environ 550 acides aminés - IPR010071) Il catalyse l'activation de l'acide aminé (figure ci-dessous) :
Le domaine A peut catalyser cette activation sans être relié à d'autres domaines. Le domaine PCP (environ 90 acides aminés) : une sérine conservée fixe le cofacteur 4'-phophopantétheine (forme apo => forme holo). Le domaine C (environ 450 acides aminés) ajoute les acides aminés via une réaction de condensation. Sa structure permet aux domaines PCP qui entourent le domaine C de positionner les substrats pour la condensation. Le domaine TE (environ 250 acides aminés) libère le produit peptidique par hydrolyse ou macro-cyclisation. |
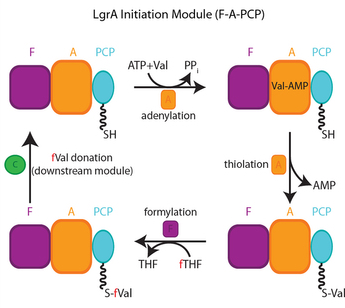
Illustration de la très grande dynamique des domaines des NRPS Voir une vidéo (source : Reimer et al., 2016) : animation du cycle de synthèse du module d'initiation de la gramicidine A synthétase linéaire (LgrA)
Source : Reimer et al. (2016)
Les structures observées expérimentalement sont annotées par des valeurs statistiques cristallographiques (parties désordonnées ou parties absentes en gris). Les détails de l'adénylate au sein du domaine A et de la valine dans le site actif (réaction de formylation) sont basés sur les co-complexes de domaines A et A-PCP et de protéines FT (voir la vidéo). |
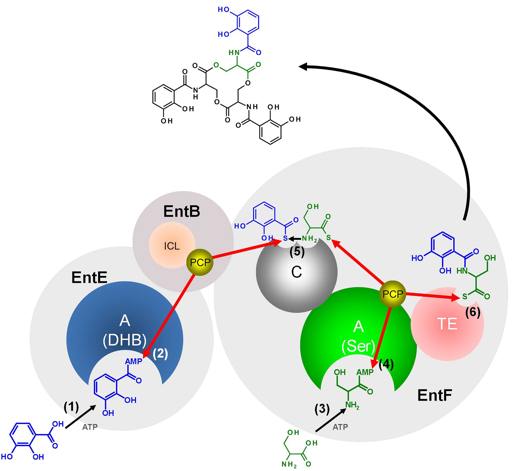
Exemple de l'entérobactine synthétase Figure ci-dessous : biosynthèse de l'entérobactine (chélateur du fer - siderophore) de Escherichia coli catalysée par deux modules de l'entérobactine synthétase (EC 6.3.2.14) qui correspond aux 4 protéines : EntD, EntE, EntB et EntF.
Source : Pfennig & Stubbs (2012) Réaction globale : 6 ATP + 3 L-serine + 3 2,3-dihydroxybenzoate <=> 6 AMP + enterobactine + 6 diphosphate + 3 H+
EntD (phosphopantétheinyl transférase) active EntB. EntE catalyse la condensation (ATP-dépendante) du 2,3-dihydroxybenzoate et de holo-EntB pour former forme arylée de EntB. EntB est bifonctionnelle :
EntF est une protéine constituées de quatre domaines qui catalyse :
|
Exemples d'autres NRPS et de leur produit
Il existe aussi des NRPS de type II constituées d'enzymes indépendantes ("standalone enzymes") : BlmI : synthèse de la bléomycine (Streptomyces verticillus ATCC 15003) |
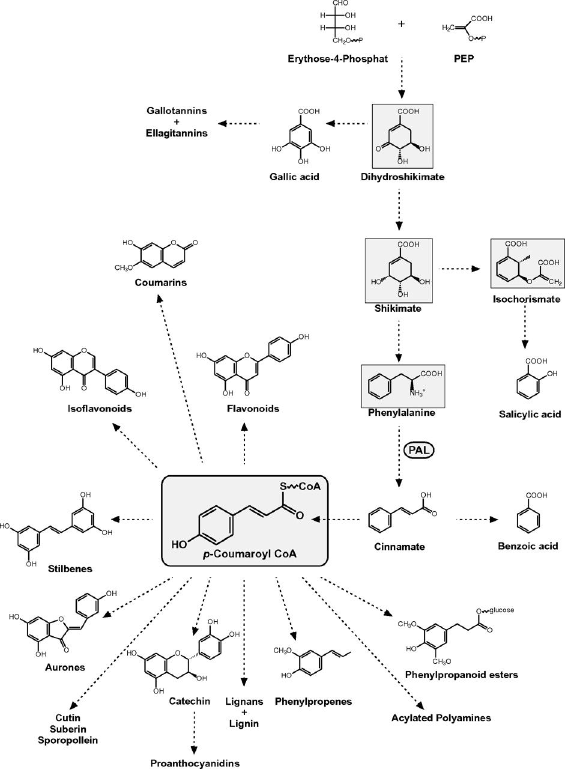
8. Composés aromatiques - Composés phénoliques Cette famille contient des composés non azotés qui possèdent des cycles aromatiques, sous forme de glycoconjugués. Ils sont en général solubles dans l'eau. Exemples : acide salicylique, isoflavones, des composés d'huiles essentielles (en association avec des terpènes), les tannins, la lignine (polymère formé à partir de dérivés de l'acide cinnamique). Les composés phénoliques des végétaux (plus de 8.000 structures) sont principalement dérivés de la voie des phénylpropanoides et de la voie de l'acétate. Ils représentent 40 % du carbone organique qui circule dans la biosphère. Ils ont des rôles dans les structures de la paroi cellulaire, les défenses des plantes, les caractéristiques du bois et des écorces, la couleur des fleurs, les saveurs ... Les composés phénoliques synthétisés et accumulés par certaines plantes (exemple : la chicorée industrielle - Cichorium intybus, Asteraceae) sont les antioxydants alimentaires les plus abondants. La chicorée industrielle produit et accumule l'acide chlorogénique, l'acide isochlorogénique, l'acide caftarique et l'acide chicorique (esters de caféoyle).
Ils dérivent de la phénylalanine, constituent l'essentiel des composés aromatiques.
Source : T. Vogt (2010)
Il s'ensuit une voie commune qui aboutit à la formation du shikimate puis du chorismate. La phénylalanine ammonia-lyase (PAL - EC 4.3.1.24) et la tyrosine ammonia-lyase (TAL - EC 4.3.1.23) catalysent la désamination non-oxydative de Phe en acide trans-cinnamique et orientent le flux de carbone de la voie du shikimate vers les diverses branches du métabolisme général des phenylpropanoïdes. Le 4-coumaroyl CoA est un métabolite carrefour clé dans la biosynthèse des phénylpropanoïdes chez les plantes. Il est le précurseur direct des flavonoïdes ou des lignines ou participe à la biosynthèse de monolignols. Une étape décisive est contrôlée par l'enzyme hydroxycinnamoyl CoA:shikimate / quinate hydroxy-cinnamoyl transférase (HCT). |
|
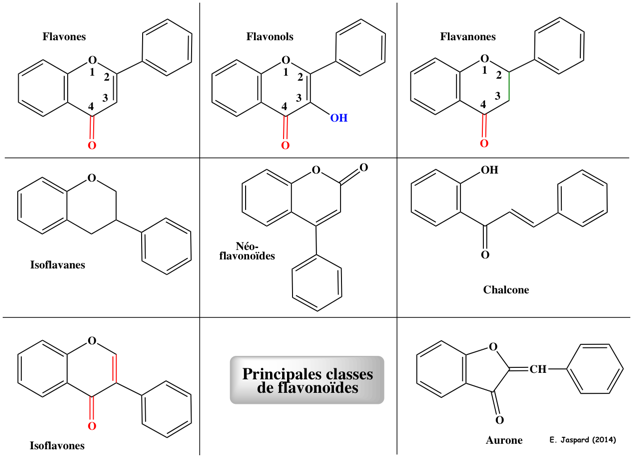
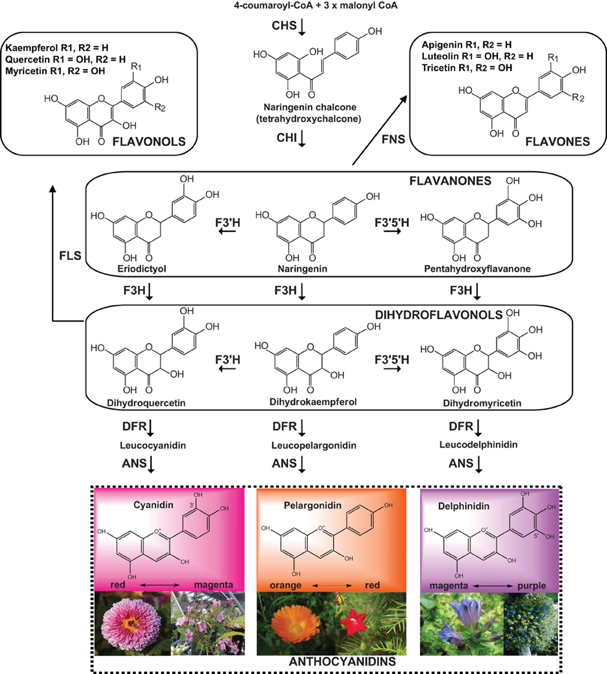
Ce sont des métabolites secondaires synthétisés par les plantes. Ce sont des pigments qui assurent de nombreuses fonctions chez les plantes. Leurs rôles physiologiques (antioxydant, antibactérien, , ...) sont très variés. Exemples de flavones (anthoxanthine) naturelles : apigenine, luteoline, tangeritin, chrysine, baicalein, wogonine.
On trouve un très grand nombre de flavonols (anthoxanthine) dans les fruits et les légumes (exemples : azaleatine, fisetine, galangine, kaempferide, kaempferol, morine, myricetine, pachypodol, rhamnetine). Le squelette de base des flavonoïdes peut subir un grand nombre de modifications : hydroxylation, glycosylation, prénylation, méthylation, acétylation. Voir la voie de biosynthèse des flavones et des flavonols. Découverte de l'interférence ARN En 1990, Napoli, Lemieux & Jorgensen ont été les premiers à décrire un phénomène d'interférence ARN, sans savoir qu'il s'agissait de ce mécanisme.
L'objectif de leur étude était de déterminer si la chalcone synthase (CHS), une enzyme clé de la biosynthèse des flavonoïdes, est l'enzyme limitante dans la voie de biosynthèse de l'anthocyane à l'origine de la coloration violette profonde du pétunia.
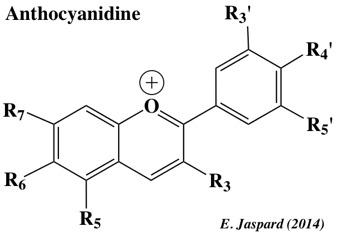
Les anthocyanes sont des pigments hydrosolubles vacuolaires rouges, violets ou bleus selon le pH. Ils appartiennent aux flavonoïdes synthétisées par la voie des phénylpropanoïdes. Les anthocyanes dérivent des anthocyanidines par addition d'oses (sur R3). Napoli, Lemieux & Jorgensen ont surexprimé la CHS en introduisant dans des pétunias un transgène codant une CHS exogène : étonnament, 42% des pétunias dans lesquels le gène CHS a été introduit, ont blanchi (seulement 9% des fleurs contrôle ont blanchi). Le niveau de [CHS endogène + CHS introduite] était 50 fois inférieur au niveau de CHS du pétunia de type sauvage. Ca a été la première mise en évidence du phénomène d'interférence ARN. Biosynthèse des anthocyanidines : cyanidine, pélargonidine et delphinidine.
Source
: Falcone Ferreyra et al. (2012) Enzymes :
|
| 9. Liens Internet et références bibliographiques |
|
LIPID Metabolites and Pathways Strategy NRPS/PKS substrate predictor DoBISCUIT : Database of BIoSynthesis cluster CUrated and InTegrated |
|
|
MAPSI : Management and Analysis for Polyketide Synthase type I PKS/NRPS Analysis Web-site The world of polyketides NORINE: a database of nonribosomal peptides |
|
|
NaPDos : a bioinformatic tool for the rapid detection and analysis of secondary metabolite genes ClusterMine360 : a database of microbial polyketide and non-ribosomal peptide gene clusters CSDB : a bioinformatics database containing genetic and biochemical information on natural products synthesised by Thiotemplate Modular Systems |
|
Staunton & Weissman (2001) "Polyketide biosynthesis: a millennium review" Nat. Prod. Rep. 18, 380-416 Admiraal et al. (2001). "The loading module of rifamycin synthetase is an adenylation-thiolation didomain with substrate tolerance for substituted benzoates" Biochemistry 40, 6116-6123 |
|
|
B. Shen (2003) "Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms" Curr. Opin. Chem. Biol. 7, 285-295 Lautru & Challis (2004) "Substrate recognition by nonribosomal peptide synthetase multi-enzymes" Microbiol. 150, 1629-1636 Koch et al. (2005) "Charting biologically relevant chemical space: A structural classification of natural products (SCONP)" PNAS 102, 17272–17277 Filippos et al. (2007) "Biotechnology of flavonoids and other phenylpropanoid-derived natural products. Part I: Chemical diversity, impacts on plant biology and human health". Biotechnol. J. 2, 1214-1234 |
|
Juguet et al. (2009) "An iterative nonribosomal peptide synthetase assembles the pyrrole-amide antibiotic congocidine in Streptomyces ambofaciens" Chem. Biol. 16, 421-431 Chan et al. (2009) "Biosynthesis of Polyketide Synthase Extender Units" Nat. Prod. Rep. 26, 90-114 Chan & Thomas (2010) "Recognition of (2S)-aminomalonyl-acyl carrier protein (ACP) and (2R)-hydroxymalonyl-ACP by acyltransferases in Zwittermicin A biosynthesis" Biochemistry 49, 3667-3677 Gao et al. (2010) "Engineered polyketide biosynthesis and biocatalysis in Escherichia coli" Appl. Microbiol. Biotechnol. 88, 1233-1242 T. Vogt (2010) "Phenylpropanoid Biosynthesis" Mol. Plant 3, 2-20 |
|
|
Strieker et al. (2010) "Nonribosomal peptide synthetases: structures and dynamics" Curr. Opin. Struct. Biol. 20, 234-240 Kraas et al. (2010) "Functional Dissection of Surfactin Synthetase Initiation Module Reveals Insights into the Mechanism of Lipoinitiation" Chem. Biol. 17, 872-880 Floss et al. (2011) "The biosynthesis of 3-amino-5-hydroxybenzoic acid (AHBA), the precursor of mC7N units in ansamycin and mitomycin antibiotics: a review" J. Antibiotics 64, 35-44 Kang et al. (2012) "Biosynthesis of 3,5-AHBA-derived natural products" Nat. Prod. Rep. 29, 243-263 |
|
Falcone Ferreyra et al. (2012) "Flavonoids: biosynthesis, biological functions, and biotechnological applications" Front. Plant Sci. 3, 222 Pfennig & Stubbs (2012) "Flexing and stretching in nonribosomal Peptide synthetases" Chem. Biol. 19, 167-169 Dias et al. (2012) "A Historical Overview of Natural Products in Drug Discovery" Metabolites 2, 303-336 A. Brakhage (2013) "Regulation of fungal secondary metabolism" Nat. Rev. Microbiol. 11, 21-32 K.M. Fisch (2013) "Biosynthesis of natural products by microbial iterative hybrid PKS–NRPS "RSC Adv. 3, 18228-18247 |
|
|
Lewis K. (2012) "Antibiotics: Recover the lost art of drug discovery" Nature 485, 439 - 440 Ling et al. (2015) "A new antibiotic kills pathogens without detectable resistance" Nature 517, 455 - 459 Reimer et al. (2016) "Synthetic cycle of the initiation module of a formylating nonribosomal peptide synthetase" Nature 529, 239–242 |
|
|
Huguenin-Dezot et al. (2018) "Trapping biosynthetic acyl-enzyme intermediates with encoded 2,3-diaminopropionic acid" Nature 565, 112 - 117 Luther et al. (2019) "Chimeric peptidomimetic antibiotics against Gram-negative bacteria" Nature 576, 452 - 458 Imai et al. (2019) "A new antibiotic selectively kills Gram-negative pathogens" Nature 576, 459 - 464 |
|
|
Torres et al. (2020) "Animal biosynthesis of complex polyketides in a photosynthetic partnership" Nat. Commun. 11, 2882 Liu et al. (2021) "Targeted Genome Mining Reveals the Biosynthetic Gene Clusters of Natural Product CYP51 Inhibitors" J. Am. Chem. Soc. 143, 6043 - 6047 Zhang et al. (2023) "Discovery of natural-product-derived sequanamycins as potent oral anti-tuberculosis agents" Cell 186, 1013 - 1025 |
![]()