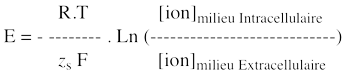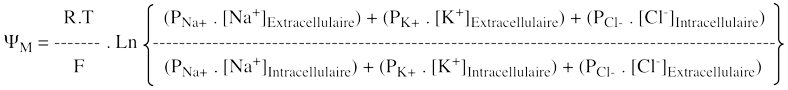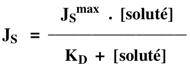| Les mécanismes de transports membranaires : passifs, actifs / pores, pompes, canaux |
| Tweet |
|
|
1. Introduction et définitions 2. Quelques lois thermodynamiques du transport membranaire 3. La diffusion simple ou diffusion libre (transport passif) 4. Le transport facilité (transport passif)
5. Le transport actif primaire ou direct
|
6. Les pompes - transport actif primaire
7. Transport actif secondaire
8. Les transports au travers des membranes mitochondriales 9. Détection du toucher et de la chaleur 10. Liens Internet et références bibliographiques |
1. Introduction et définitions Les transports membranaires déplacent des ions ou des molécules au travers des membranes biologiques séparant le milieu intra-cellulaire du milieu extra-cellulaire ou séparant deux compartiments sub-cellulaires.
Le flux net d'un soluté S (Js) au travers d'une membrane signifie qu'il existe une force qui agit sur ce soluté et qui détermine le sens de ce flux. Cette force résulte de gradients transmembranaires de potentiels thermodynamiques : gradients de pression osmotique, gradients chimiques ou électrochimiques, gradients de température.
Remarques concernant les gradients de température
|
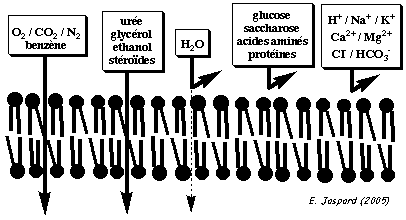
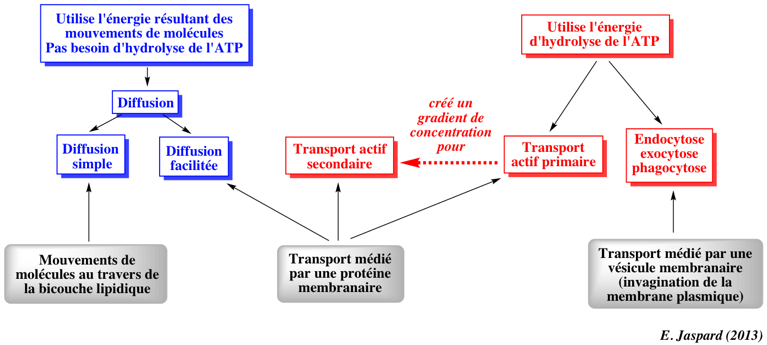
| Les différents types de transport |
| Les transports passifs | Les transports actifs | |
| Ils ne nécessitent pas d'énergie car ils s'effectuent dans le sens du gradient de concentration ou du gradient électrochimique. | Ils nécessitent de l'énergie car ils s'effectuent contre le gradient de concentration (transport non spontané). | |
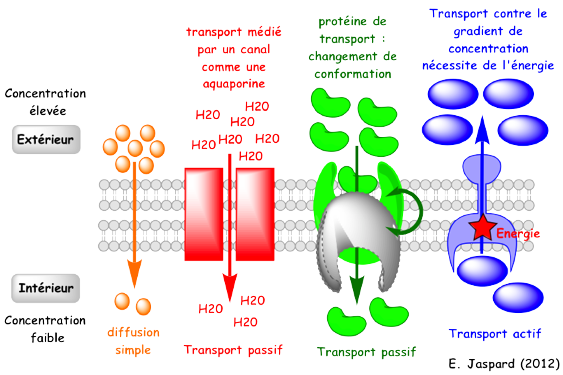
La diffusion directe (ou diffusion simple ou diffusion libre) : les molécules "liposolubles" diffusent au travers de la membrane biologique. La diffusion facilitée : les molécules utilisent une protéine de transport. L'osmose : mouvement net de molécules de solvant au travers d'une membrane semi-perméable vers un compartiment contenant une concentration plus importante d'un soluté. Ce mouvement tend à égaliser la concentration de ce soluté des 2 côtés de cette membrane. La différence de concentration engendre une différence de pression (gradient de pression) : c'est la pression osmotique qui provoque ce mouvement. |
Transport actif primaire (ou direct) : l'énergie est fournie par l'hydrolyse d'un nucléotide triphosphate (exemple : pompes à sodium et hydrolyse de l'ATP).
Transport actif secondaire (ou couplé) : l'énergie est fournie par une différence de potentiel électrochimique (exemple : un gradient de concentration de sodium). Le terme "secondaire" signifie que cette différence de potentiel électrochimique résulte d'un transport actif primaire (exemple : pompe à sodium et hydrolyse de l'ATP). |
Figures ci-dessous : illustration schématique des types de transports transmembranaires.
Figure ci-dessous : résumé des différents types de transports membranaires.
Le passage de matériel biologique trop volumineux ou trop réactif s'effectue par deux autres processus : l'endocytose et l'exocytose. Dans les deux cas, ce transport s'effectue par l'intermédiaire d'une vésicule lipidique, résultat d'une invagination de la membrane plasmique. Voir le cours : "Le trafic vésiculaire - les protéines SNARE". |
| Type de transport | Type de protéine de transport | saturable par le substrat transporté | production d'un gradient de concentration | dépendant de l'énergie |
| passif - diffusion simple | --------- | non | non | non |
| passif - diffusion facilitée | transporteur / perméase : diffusion d'une espèce chimique | oui | non | non |
| canal ionique / pore : diffusion d'une espèce chimique | non | non | indirectement | |
| actif primaire | les pompes :
|
oui | oui | oui |
| actif secondaire |
|
oui | oui | oui |
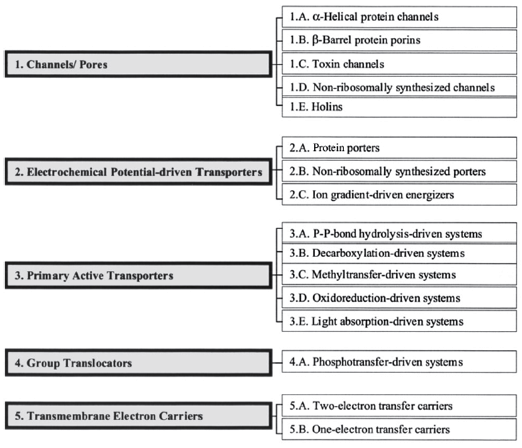
Figure ci-dessous : "The Transporter Classification system". Plus de 400 familles de transporteurs ont été recensées.
Source : Busch & Saier (2002) |
| "Transporter Classification database" : système de classification des protéines de transport membranaire | |
|
Class 1. Channels/Pores
Class 2. Electrochemical Potential-Driven Transporters
Class 3. Primary Active Transporters
|
Class 4. Group Translocators Class 5. Transmembrane Electron Carriers
Class 8. Accessory Factors
Class 9. Incompletely Characterized Transport Systems
|
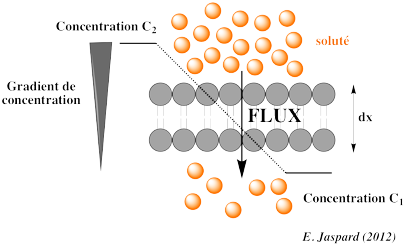
2. Quelques lois thermodynamiques du transport membranaire a. Les lois de la diffusion de Fick Considérons le flux Js d'un soluté S en solution. A est la surface (en m2) à travers laquelle le flux se dirige. Remarque : la diffusion a lieu dans les 3 dimensions mais le flux n'est considéré que dans 1 dimension par simplification. La diffusion a lieu le long d'un gradient de concentration : des fortes concentrations vers les faibles concentrations.
Les lois de la diffusion ont été développées par Adolf Fick en 1855 : 1ère loi de Fick : Js = - Ds . (∂Cs / ∂x) = - b . Cs . (∂μs / ∂x) 2ème loi de Fick : ∂Cs / ∂t = Ds . (∂2Cs / ∂x2)
Voir une démonstration des équations de Fick. |
b. Equation de Ussing-Teorell Elle permet de déterminer si le transport d'un soluté est passif (sans interaction avec d'autres substances) et ne met donc en jeu aucun type de transport actif. |
Mesure expérimentale des flux transmembranaires d'un soluté S :
On calcule le rapport de ces flux : ƒ = influxs / effluxs |
Equation de Ussing-Teorell : ƒ = exp (- Δμs / R.T)
Si le soluté S n’est pas chargé : f = CExtracellulaires / CIntracellulaires |
Conclusion : si la valeur expérimentale de ƒ est égale à la valeur calculée avec l'équation de Ussing-Teorell, le transport est passif. |
|
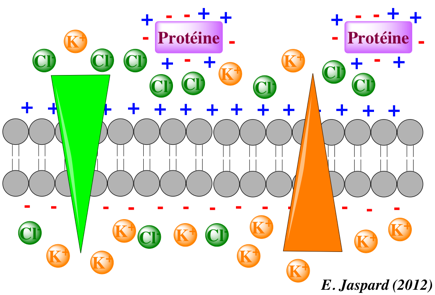
c. Définitions préalables au transport de solutés chargés ou non Plusieurs facteurs entraînent une distribution différente des ions ou des molécules de part et d'autre d'une membrane :
Les conséquences sont :
Ce potentiel de membrane influe sur le mouvement des molécules chargées : les molécules positives se dirigent vers le milieu intracellulaire (négatif) et inversement pour les molécules négatives. En conséquence, le déplacement des molécules chargées au travers des membranes est donc régi par deux forces :
La combinaison de ces deux forces est appelée gradient électrochimique. Une protéine de transport qui engendre un potentiel de membrane est une pompe électrogènique (exemple : pompe [Na+/K+]). |
d. Expression de la force résultant d'une différence de potentiel chimique ou électrochimique transmembranaire α. Si le soluté S n'est pas chargé, le potentiel chimique s'écrit : ΔG' = RT . Ln (CsIntracellulaire / CsExtracellulaire) β. Si le soluté S est chargé, il faut tenir compte aussi de la différence de potentiel électrique entre les deux faces de la membrane (la face de la membrane du côté du milieu intracellulaire est négative), proportionnelle au potentiel de membrane ΔΨM. Le potentiel électrochimique s'écrit : ΔG' = [RT . Ln (CsIntracellulaire / CsExtracellulaire)] + (zs . F . ΔΨM)
Voir le formalisme de la force proton-motrice de l'ATP synthase générée par la chaîne respiratoire. Remarque : l'ATP synthase synthétise l'ATP. Les pompes ATPases hydrolysent l'ATP. |
e. Perméabilité d'une membrane et potentiel de Nernst Le milieu extracellulaire E et le milieu intracellulaire I sont séparés par la membrane M de la cellule. Le saut de concentration d'un soluté S entre la phase membranaire M et les phases aqueuses I et E, est caractérisé par un coefficient de partage : ks= CMs / Cs Si le flux du soluté S au travers de la membrane est constant, alors : Js= - Ds . (∂Cs / ∂x) = [ks. Ds / h] . (CIs - CEs)
A l'équilibre thermodynamique, en absence de flux ou de potentiel électrostatique, le passage d'un ion au travers de l'interface [milieu aqueux / membrane], nécessite de l'énergie.
Pour un soluté chargé, il faut aussi tenir compte du potentiel électrique car le rapport des activités d'un soluté (as) de charge zs entre les deux phases est : asM / asI = ks . exp (-zs . F . ΔΨM-I / RT)
En absence de différence de potentiel entre les phases, le rapport d'activité est déterminé par le coefficient de partage. Dans le cas des solutions idéales (solutions diluées), l'activité se réduit à la concentration : asM / asI = CsM / CsI La différence de potentiel interfacial s'écrit alors : ΔΨM-I = ΨM - ΨI = - [RT / zs . F] . ln (CsM / ks.CsI) Pour le passage entre le milieu intracellulaire I et le milieu extracellulaire E au travers d'une membrane M, l'application aux 2 interfaces I->M et M->E pemet d'obtenir le potentiel de Nernst : ΔΨM = ΨI - ΨE = - [RT / zs . F] . ln (CsIntracellulaire / CsExtracellulaire) Ce résultat n'est autre que celui obtenu pour ΔG' = 0 (force du potentiel électrochimique / voir le paragraphe "d." ci-dessus), c'est-à-dire pour un processus à l'équilibre. |
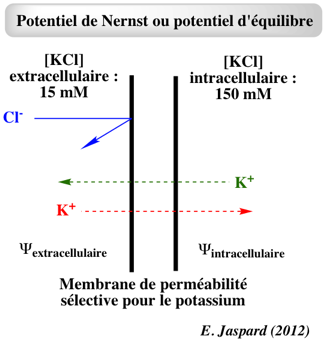
f. Equations pour le calcul du potentiel de membrane α. Potentiel d'équilibre pour un ion ou potentiel de Nernst pour un ion
Si la membrane n'est perméable qu'à K+, cet ion diffuse du milieu intracellulaire (concentration plus élevée) vers le milieu extracellulaire (concentration plus faible). L'ion Cl- ne peut pas diffuser.
Très rapidement, la différence de potentiel électrique déplace K+ vers le milieu intracellulaire à la même vitesse que la vitesse de déplacement de K+ vers le milieu extracellulaire en raison du potentiel chimique (le gradient de concentration de K+).
Remarque : cette expression est équivalente à celle décrite ci-dessus. La valeur du potentiel d'équilibre d'un ion dépend donc du gradient de concentration de cet ion au travers de la membrane :
|
β. Equation de David Goldman, Alan Hodgkin et Bernard Katz (1943 et 1949) Cette équation est une généralisation de l'équation de Nernst dans le cas de plusieurs types d'ions. Cette équation décrit le potentiel d'une membrane M (ou potentiel transmembranaire ΨM) qui sépare 2 solutions contenant, par exemple, NaCl et KCl.
On obtient :
Voir une démonstration de l'équation de Goldman-Hodgkin-Katz. Voir un article qui retrace l'historique et détaille les équations de Goldman (1943) et de Hodgkin & Katz (1949). |
γ. Equation de David Goldman Cette équation décrit le potentiel de membrane quand il y a des différences de concentrations de divers ions de part et d'autre de la membrane.
Il s'ensuit un déséquilibre des charges entre les 2 faces de la membrane : ce potentiel ralentit à son tour le mouvement des ions. Quand la différence nette des charges atteint un équilibre, un potentiel de diffusion est généré qui dépend de la perméabilité des différents ions. Le potentiel de membrane est alors calculé par l'équation de Goldman :
Voir une démonstration de l'équation de Goldman. Aller aux sites "Goldman-Hodgkin-Katz Equation" (applet java). |
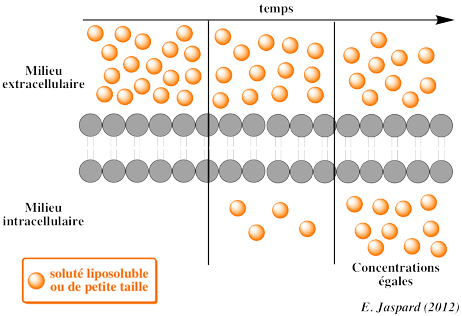
3. La diffusion simple ou diffusion libre (transport passif) La diffusion simple est la diffusion d'un soluté au travers d'une membrane dans le sens fortes concentrations vers les faibles concentrations (gradient de concentration), jusqu'à atteindre un équilibre des concentrations des 2 côtés de la membrane.
Ce type de diffusion nécessite que le soluté soit soluble dans la bicouche de phospholipides constitutive de la membrane :
Les caractéristiques de ce transport sont :
|
| Soluté | Concentration intracellulaire (mM) | Concentration extracellulaire (mM) | Perméabilité de la membrane (molécules qui diffusent) | Imperméabilité de la membrane |
| Na+ | 12 - 15 | 145 |
|
|
| K+ | 120 - 140 | 4 - 4,5 | ||
| Mg2+ | 0,8 | 1,5 | ||
| Ca2+ | 1 10-4 (≈ 100 nM) | 1,2 - 2 | ||
| Cl- | 5 - 20 | 110 - 116 | ||
| HCO3- / PO43- / protéines / acides nucléiques ... | 148 | 42 |
La concentration de H+ dans les fluides corporels est d'environ 40 10-9 moles / L (<=> pH 7,4). Par ailleurs, chez un être humain de 70 kg à l'état basal, le taux de production d'acide :
Le flux d'H+ au travers de l'organisme en 24 heures est 108 fois plus élevé que la quantité totale d'H+ libre dans toute l'eau du corps (< 2 μmoles). Cette homéostasie extraordinaire est assurée par :
|
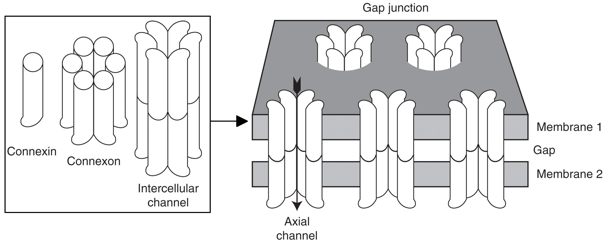
4. Le transport facilité (transport passif) a. Types de protéines du transport facilité Le moteur de la diffusion facilitée est le gradient de concentration (comme la diffusion passive). Mais la diffusion facilitée s'opère via des protéines transmembranaires intrinsèques qui sont spécialisées dans le transport de solutés spécifiques. α. Les canaux ioniques qui permettent la communication entre cellules. Exemple : les jonctions gap (ou jonctions communicantes ou "gap junction").
Figure ci-dessous : les niveaux multiples de la structure des canaux jonctions gap. Ils sont composés de protéines intrinsèques de taille moyenne : les connexines (vertébrés, PFAM PF00029) ou les innexines (invertébrés).
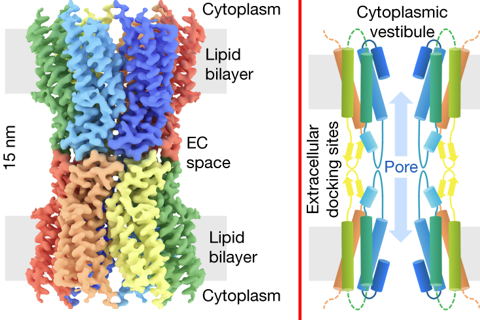
Source : Goodenough & Paul (2009) Les connexines contiennent 4 domaines trans-membranaires. Elles sont synthétisées au niveau du réticulum endoplasmique rugueux puis insérées au réticulum endoplasmique au cours de leur traduction. Les connexines s'assemblent à l'intérieur de la cellule sous forme d'hexamères, appelés connexons qui sont dirigés vers la surface de la cellule. Il s'y associent à d'autres connexons de cellules adjacentes : cela forme un canal axial (qui traverse les 2 membranes plasmiques) et un orifice extracellulaire étroit (le "gap"). Figure ci-dessous, à gauche : reconstruction 3D par cryo-microscopie électronique du co-assemblage (hauteur = 15 nm) des connexines dans des canaux de jonction ("lens gap junction channels") de la lentille de l'oeil composés de connexine 46 et de connexine 50 (Cx46/50). Les douze sous-unités sont colorées différemment. Les régions correspondant à la bicouche lipidique, au cytoplasme et à l'espace extracellulaire (EC) sont indiquées.
Source : Myers et al. (2018) Figure ci-dessus, à droite : modèle du repliement de la section transversale du canal intercellulaire assemblé, avec les domaines TM1 à 4, EC1, EC2 et l'hélice N-terminale NTH. Les domaines extra-cellulaires EC1 et EC2 forment des sites d'amarrage qui constituent un pore / canal continu de 1,4 nm de diamètre reliant les cytoplasmes des cellules voisines.
|
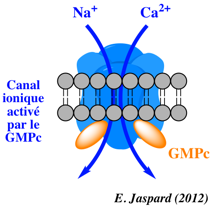
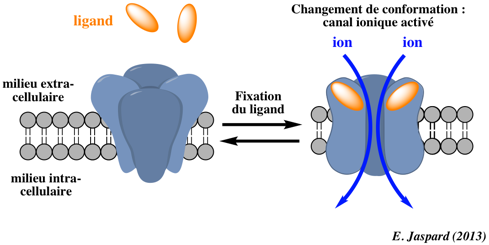
β. Les canaux ioniques qui s'ouvrent en réponse à un stimulus chimique ou électrique précis ("Ligand-Gated Ion Channels").
Exemples de canaux ioniques et de ligands activateurs
|
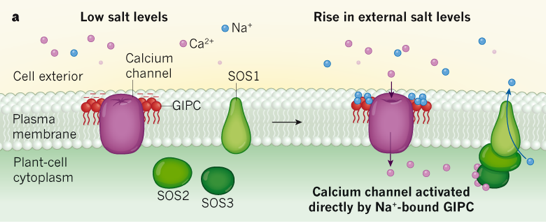
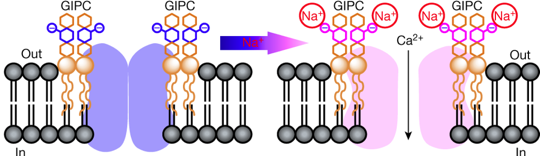
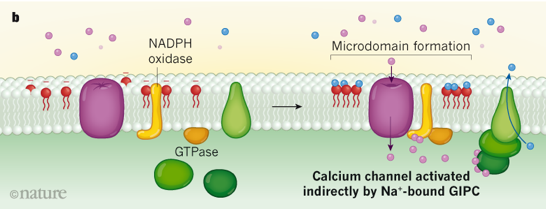
Illustration de l'activation d'un canal ionique Modèles expliquant une manière qu'ont les plantes de percevoir la teneur en sel et d'activer certains canaux calciques. Modèle a. Lorsque les ions Na+ sont détectés à l'extérieur d'une cellule végétale, un canal calcique est activé et des ions Ca2+ pénètrent dans la cellule.
Source : Steinhorst & Kudla (2019)
Source : Jiang et al. (2019) Modèle b
Source : Steinhorst & Kudla (2019) |
γ. Les pores sélectifs comme les porines ou les aquaporines. On les trouve dans la membrane externe des bactéries (Gram - et certaines Gram +), de la mitochondrie et du chloroplaste.
Voir un cours sur les porines et les aquaporines. δ. Les protéines de transport ("carrier") ou perméases Voir chapitre suivant : "b. Caractéristiques du transport facilité". |
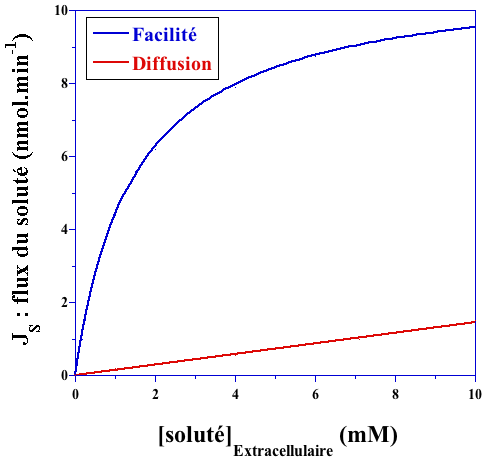
b. Caractéristiques du transport facilité Le transport facilité augmente considérablement la vitesse de transport d'un soluté par rapport à la diffusion simple (figure ci-dessous).
Remarque : certains solutés transportés par transport facilité sont aussi transportés, à un flux beaucoup plus faible, par diffusion simple. La droite en rouge (diffusion simple) est donc parallèle à la fin de la courbe en bleu (transport facilité). Il y a une spécificité de reconnaissance entre la protéine de transport facilité et le soluté transporté (comme les enzymes vis-à-vis de leur substrat), d'où une saturation quasi-hyperbolique (courbe en bleu - figure ci-dessus).
On peut déterminer une constante de dissociation KD du soluté transporté :
La structure du site de fixation du substrat de GLUT5 (transporteur spécifique du fructose) est très proche de celles de GLUT1 (transporteur spécifique du glucose) de l'homme et de XylE de Escherichia coli.
Le transport facilité peut être inhibé (inhibition compétitive ou non compétitive / inactivation). Exemple : la perméase GLUT2 transporte le glucose, le fructose et le galactose. Le galactose étant un analogue structural du glucose, certains sites du transporteur sont occupés par le galactose (inhibition compétitive). |
|||||||||||||||
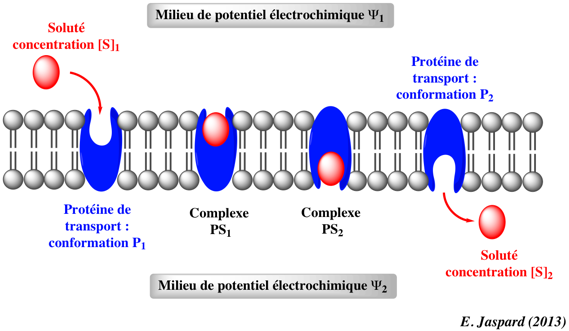
c. Formalisme thermodynamique du transport facilité La force motrice de la diffusion passive d'un soluté S de charge zs au travers d'une membrane est la différence de potentiel électrochimique de ce soluté entre les 2 côtés de la membrane. Potentiel électrochimique d'un soluté S de charge zs : μs = μ0s + (RT log as) + (zs . F . Ψ)
Considérons le transport facilité d'un soluté S de charge zs, médié par un transporteur P, d'un milieu de potentiel électrique Ψ1 vers un milieu de potentiel électrique Ψ2.
On fait les approximations suivantes :
L'expression de la force motrice de la diffusion passive devient : ΔμsElectroChimique = μ2s - μ1s = RT (log [S2] - log [S1]) + [zs . F . (Ψ2 - Ψ1)] Remarque : dans le cas du transport de plusieurs solutés, l'expression est la même en sommant les différences de potentiels électrochimiques pour obtenir la force nette globale.
|
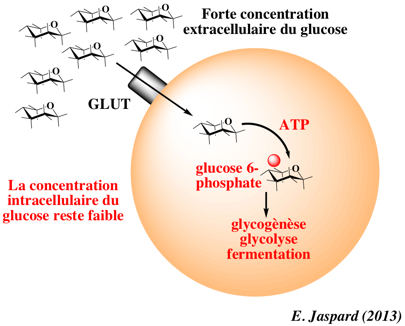
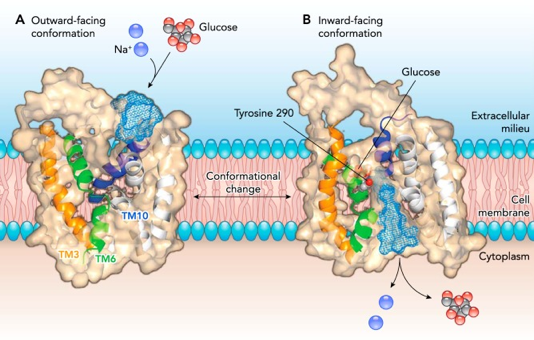
e. Exemple du transport facilité du glucose par les perméases GLUT Les perméases du glucose de la famille GLUT ("GLUcose Transporter") assurent le transport facilité de divers oses, notamment celui du glucose. Les transporteurs GLUT existent à la surface de toutes les cellules. Les transporteurs GLUT fonctionnent dans les 2 sens (entrée et sortie du glucose) en fonction du gradient de concentration :
Certaines cellules où a lieu la diffusion facilitée peuvent maintenir un état éloigné de l'équilibre en maintenant la concentration d'une molécule à une concentration très faible. C'est le cas du glucose qui entre dans la cellule car il est immédiatement phosphorylé en glucose 6-phosphate (hexokinase ou glukokinase).
Il existe de nombreuses isoformes qui constituent une famille de 14 transporteurs d'hexoses chez l'homme (GLUT1 à GLUT12, GLUT14, et HMIT) subdivisée en 3 classes. |
| Quelques transporteurs d'oses GLUT : 12 hélices α transmembranaires. |
| GLUT1 (gène SLC2A1) : transcrit dans divers tissus mais à un faible niveau dans le foie et les muscles squelettiques. C'est le principal transporteur du glucose dans les érythrocytes (voir ci-dessous). |
| GLUT2 (gène SCL2A2) : transcrit essentiellement dans le foie, les cellules pancréatiques β, les reins et les intestins. Transport bi-directionnel du glucose au travers de la membrane. Il transporte aussi le fructose et le galactose. |
| GLUT3 (gène SCL2A3) : transcrit essentiellement dans les neurones. De tous les transporteurs du glucose, c'est celui qui a la valeur de KD la plus faible. |
|
| GLUT5 (gène SCL2A5) : transcrit essentiellement dans les entérocytes et les intestins. Il transporte spécifiquement le fructose. |
| GLUT6 (gène SCL2A6) : transcrit essentiellement dans le cerveau et les leukocytes. Il transporte le glucose. Attention : il a aussi comme appellation GLUT9 mais il est différent du suivant. |
| GLUT9 (gène SCL2A9) : transcrit essentiellement dans les reins, le foie et le placenta. Il transporte le glucose à une vitesse faible. Il transporte aussi l'urée et le fructose. |
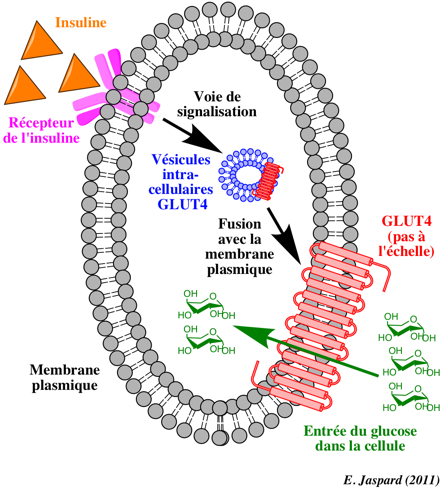
Transport insulino-dépendant du glucose par GLUT4 Dans une cellule non stimulée ou quand la concentration en insuline est faible, le transporteur insulino-dépendant du glucose GLUT4 ("GLUcose Transporter 4") est localisé dans des vésicules de stockage des cellules hépatiques et musculaires.
Quand le niveau de glucose circulant est élevé, l'insuline est libérée par les ilots de Langerhans :
|
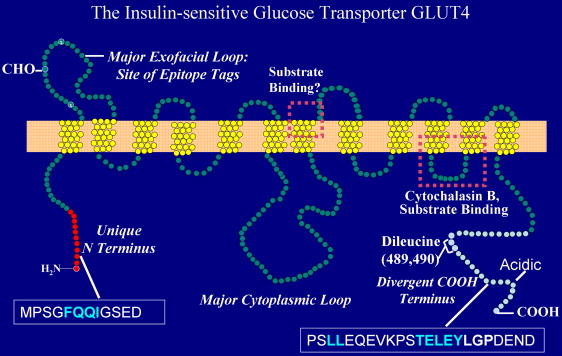
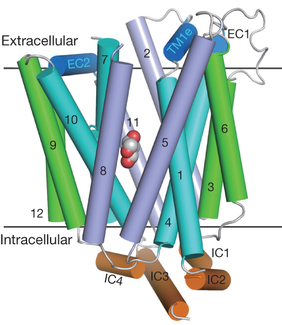
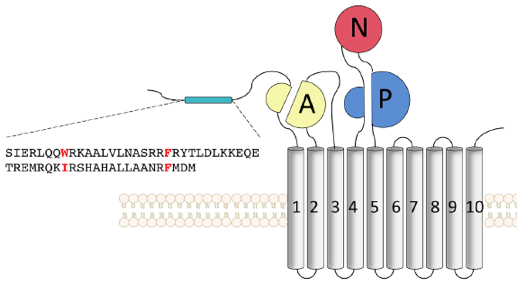
Structure de GLUT4 La structure de GLUT4 de l'homme est prédite par le logiciel d'apprentissage profond (intelligence artificielle) AlphaFold - EBI. La chaîne polypeptidique de GLUT4 contient 509 acides aminés qui forment 12 hélices α transmembranaires :
Ces 2 domaines forment un pseudo-axe de symmétrie autour d'un tunnel central polaire qui laisse entrer/sortir le glucose. Ce tunnel de nature amphipathique semble formé par les hélices 1, 2, 4, 5, 7, 8, 10 et 11. Les séquences cytoplasmiques des extrémités N-terminale et C-terminale sont particulières et expliquent en grande partie l'aptitude au traffic transmembranaire de ce transporteur.
Source : Huang & Czech (2007)
Ces motifs dirigent probablement les aspects cinétiques de l'endocytose et de l'exocytose dans un systéme continu de traffic de recyclage. Les deux leucines et les motifs acides de l'extrémité C-terminale sont trouvés également dans une aminopeptidase ("Insulin-Regulated AminoPeptidase" - IRAP) qui, de manière semblable dans les adipocytes, est séquestrée dans des membranes intracellulaires enrichies en GLUT4 et sujette à l'action de l'insuline. |
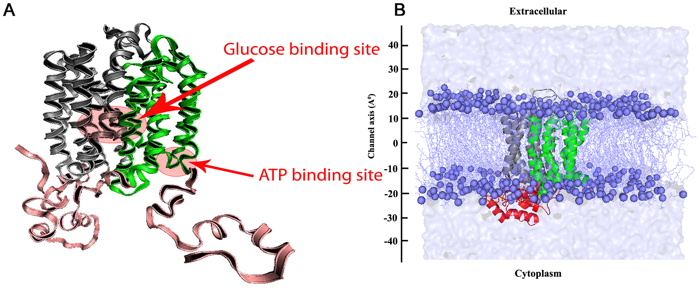
Régulation de l'activation de GLUT4 et rôle inhibiteur de l'ATP Pour être fonctionnel, GLUT4 doit être activé au niveau de la membrane plasmique. Cette activation est régulée :
La fixation de l'ATP sur GLUT1 (transporteur du glucose des érythrocytes) supprime son activité de transport. Diverses expériences et un modèle structural obtenu par simulation (figure ci-dessous) tendent à montrer un rôle semblable de l'ATP pour GLUT4.
Source : Mohan et al. (2010) Quand l'ATP est fixé, l'interface entre les 2 domaines de GLUT4 adopte une conformation plus compacte en raison de réarrangements de nombreuses liaisons hydrogène et de ponts salins inter-domaines. La poche de fixation de l'ATP est formée par la boucle qui relie les hélices transmembranaires VIII et IX. Cette poche confine le motif de fixation de l'ATP (GRRTLHL) : elle est entourée par l'extrémité cytoplasmique des hélices transmembranaires V, X et XII.
|
|
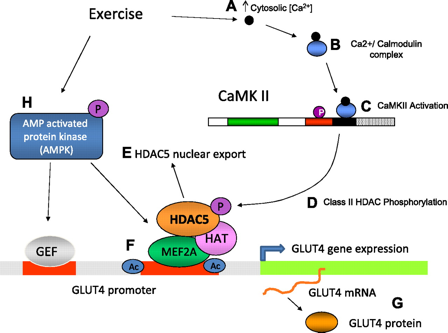
Mécanismes de la régulation de la transcription de GLUT4 par la CaMKII
Source : Ojuka et al. (2012) Etape A : la contraction des muscles pendant un exercice physique augmente la concentration cytosolique de Ca2+. Etape B : Ca2+ se fixe à la calmoduline et l'active. Etape C : le complexe [Ca2+/calmoduline] se fixe sur le domaine "calmoduline-like" de la CaMKII et l'active. Etape D : la CaMKII phosphoryle les HDAC de la classe II au sein des complexes [MEF2/HDAC] fixés sur le promoteur du gène Glut4, ce qui induit la dissociation des complexes et l'export du noyau des HDAC (étape E). Etape F : le facteur de transcription MEF2 forme alors des complexes avec des "HAT-like" comme p300. Les HAT acétylent les histones aux voisinage du domaine de MEF2 qui se fixe sur le gène Glut4. Cela augmente l'accessibilité des facteurs de transcription et des ARN polymérases, ce qui augmente le taux de synthèse de GLUT4 (étape G). Etape H : L'activation de l'AMPK (protéine kinase activée par l'AMP) est un autre signal important qui médie l'augmentation de la transcription du gène GLUT4 induite par l'exercice physique. |
f. Les protéines de transport facilité de la superfamille MFS Chez les procaryotes, 25% des protéines de transport qui assurent la diffusion facilitée appartiennent à une superfamille : "Major Facilitator Superfamily" ou MFS.
XylE est un symport de la superfamille MFS ([D-xylose/H+]) de Escherichia coli, homologue de GLUT1, GLUT2, GLUT3 et GLUT4. Figure ci-dessous : structure de XylE fixé au D-xylose (au centre de la figure). Les hélices extracellulaires et intracellulaires sont colorées en bleu foncé (en haut) et en orange (en bas), respectivement.
Source : Sun et al. (2012) - PDB : 4GBY Le repliement de XylE est typique de la superfamille MFS :
Le D-xylose est piègé au centre du domaine transmembranaire : ce domaine est complètement occlu du côté intracellulaire mais il est accessible au solvant du côté extracellulaire via un canal trop étroit pour permettre la fuite du D-xylose.
|
g. Mécanisme de couplage allostérique entre la liaison du soluté et le déclenchement fermeture - ouverture des accès au soluté Le protozoaire Plasmodium falciparum est le parasite responsable du paludisme qui est la cause de plus de 400.000 décès chaque année. L'évolution constante de Plasmodium falciparum lui permet de résister aux thérapies. De nouvelles voies doivent donc être explorées en permanence afin de développer des médicaments capables de lutter contre cet organisme. Le transporteur 1 d'hexose de Plasmodium falciparum (PfHT1) fixe les oses des globules rouges de l'hôte infecté pour survivre dans ces cellules : cette protéine est donc une cible pharmacologique. PfHT1 (504 acides aminés, 12 hélices α transmembranaires prédites) : classification TCDB : 2.A.1.1.24 - Pfam PF00083. Contrairement à de nombreuses protéines de transport des oses qui sont spécifiques d'un ose, PfHT1 transporte plusieurs types d'oses de manière tout aussi efficace (KMglucose = 1 mM). La structure de PfHT1 lui permet de coupler la fixation des oses dans leur site de liaison au processus par lequel ils sont véhiculés au travers de la protéine. Ce couplage accroît sa capacité à transporter efficacement une large gamme d'oses.
Les mêmes résidus d'acides aminés de PfHT1 sont nécessaires pour fixer le D-glucose et le D-fructose. Par ailleurs, le remplacement des résidus dans et autour du site de liaison des oses de PfHT1 par des résidus d'acides aminés que l'on trouve dans les transporteurs GLUT n'a conféré aucune sélectivité de type GLUT aux protéines PfHT1 ainsi modifiées. La faible sélectivité d'oses de PfHT1 ne résulte donc pas seulement de la nature des résidus d'acides aminés de son site de liaison.
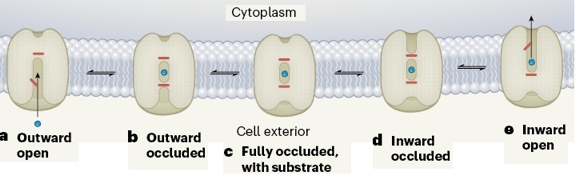
Source : Althoff & Abramson (2020) Depuis que les premières structures de transporteurs de la superfamille MFS ont été décrites, on sait que des faisceaux d'hélices α se "balancent" autour du site central de liaison du soluté transporté. Il en résulte 3 conformations alternées qui génèrent une voie de passage au travers de la protéine :
Dans le cas de PfHT1, tous les résidus de liaison des oses conservent leur orientation tout au long du cycle de translocation : les changements entre les 3 conformations ne résultent pas de réarrangements structuraux du site de liaison des oses. Mécanisme de PfHT1
En conclusion, contrairement à ses homologues chez l'homme, l'aptitude de PfHT1 à transporter de nombreux oses résulte de sa dynamique de déclenchement contrôlée par les solutés transportés, ce qui lui permet d'adopter la conformation occluse plus facilement et plus rapidement. Voir un cours sur l'allostérie. PfHT1 est inhibée par des petites molécules anti-paludiques (C3361 et MMV009085) qui se fixent dans une poche hydrophobe dans PfHT1. Cette propriété pourrait faciliter la conception de nouveaux médicaments antipaludiques. |
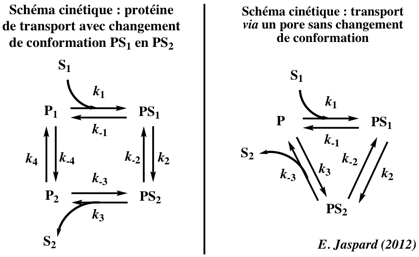
5. Le transport actif primaire ou direct Symport : transport de deux (ou plus) solutés dans le même sens via un gradient électrochimique. Sextracellulaire + H+extracellulaire ou Na+extracellulaire <=> Sintracellulaire + H+intracellulaire ou Na+intracellulaire La vitesse de transport par les uniports (transport d'un seul soluté dans un seul sens) ou les symports est d'environ 102 - 104 molécules.sec-1. Antiport : un soluté est transporté dans le sens de son gradient de concentration, l'autre soluté est transporté dans le sens opposé à son gradient de concentration. La protéine qui assure ce type de transport existe dans deux conformations (P et P*) en interconversion : S1extracellulaire + P <=> S1intracellulaire + P* P* + S2intracellulaire <=> S2extracellulaire + P Exemples de protéines membranaires effectuant un transport actif primaire :
Exemples de transport actif par les protéines de transport de la superfamille MFS :
Une nouvelle classe d'enzyme : les translocases - E.C. 7 Aucune des 6 classes d'enzymes ne décrivait correctement le groupe d'enzymes qui ont pour fonction le mouvement des ions ou des molécules au travers des membranes. Certaines de ces enzymes hydrolysant l'ATP étaient jusqu'alors classées en tant que ATPases (E.C. 3.6.3.-).
Les sous-classes d'enzymes suivantes désignent les types d'ions ou de molécules transportés : Voir la nomenclature de l'IUMB ("International Union of Biochemistry and Molecular Biology"). |
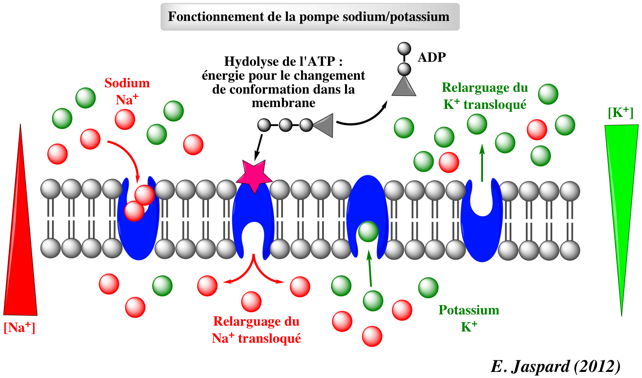
b. Principe du transport actif primaire Le transport actif primaire utilise de l'énergie (souvent issue de l'hydrolyse de l'ATP) pour transporter un soluté contre son gradient de concentration.
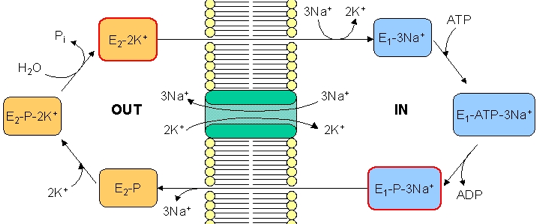
c. Description du cycle de la pompe à sodium - potassium ou ATPase - [Na+/K+] 1. Cet antiport est initialement dans un état E1 à haute affinité pour l'ion Na+. Il fixe 3 ions Na+ dans le cytosol : formation du complexe E1-3Na+. Les résidus aspartate sont phosphorylés. On aboutit au complexe E1-ATP-3Na+. 2. L'hydrolyse de l'ATP génère un complexe à haute énergie E1-P-3Na+.
Source : "Membrane III" 3. La pompe fixe alors 2 ions K+ du milieu extra-cellulaire et forme le complexe E2-P-2K+. 4. L'hydrolyse du groupement phosphate génère un complexe à haute énergie E2-2K+. 5. L'énergie de ce complexe lui permet de changer de conformation : le site de fixation de K+ est alors orienté vers le cytosol. Il s'ensuit le relarguage de 2 ions K+. La forme E1 de la pompe est régénérée et le processus peut recommencer. |
|
6. Les pompes - transport actif primaire La vitesse de transport des ions par les pompes est d'environ 102 - 103 ions.sec-1. a. Les différents types de pompes α. Les pompes ioniques ou ATPases ionophores : l'énergie qu'elles libèrent en hydrolysant l'ATP est utilisée pour le transport d'ions. Il en existe différents types. ATPases de type "P"
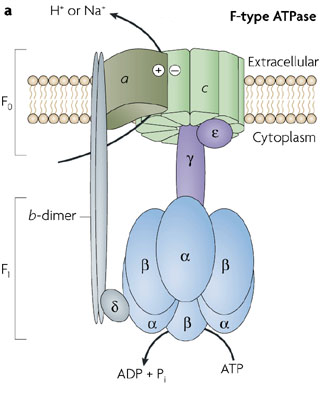
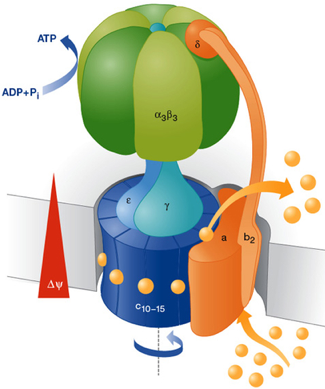
ATPases de type "F" (ou FoF1-ATPase) : on les trouve dans la membrane plasmique des bactéries, la membrane interne de la mitochondrie, la membrane des thylacoïdes des chloroplastes.
Source : Armen et al. (2007) ATPases de type "V" (ou vacuolaires)
Autres types d'ATPases
La superfamille des transporteurs ABC ("ATP Binding Cassette transporters")
β. Les pompes phosphotransférases (Escherichia coli) : c'est un mécanisme par translocation d'un groupe phosphoryle dont la source d'énergie est le phosphoénolpyruvate. γ. Les pompes photosensibles
|
|
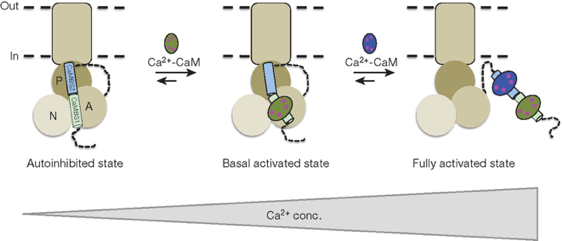
b. Mécanisme d'activation des pompes ATPases - Ca2+ par la calmoduline Les ions calcium sont des messagers secondaires de nombreuses voies de transduction du signal. Les cellules investissent beaucoup d'énergie dans le contrôle et le maintien d'un gradient entre la concentration intracellulaire (≈ 0.1 μM) et extracellulaire (≈ 2 mM) de calcium. La famille des pompes à calcium activées par la calmoduline, à laquelle appartiennent les Ca2+-ATPases de la membrane plasmique ("Plasma-Membrane Ca2+-ATPase"), sont des régulateurs clé de la concentration intracellulaire de calcium chez les Eucaryotes. Contrairement aux autres ATPases de type P, les Ca2+-ATPases contiennent un domaine N-terminal (chez les plantes) ou C-terminal (chez les mammifères) responsable de l'auto-inhibition. Le domaine N-terminal auto-inhibiteur contient plusieurs acides aminés basiques conservés capitaux pour l'auto-inhibition : Arg58, Arg61, Lys67 et Lys68.
Source : Tidow et al. (2012) - Structure de la Ca2+-ATPase de Arabidopsis thaliana A : domaine activateur; N : domaine de fixation du nucléotide; P : domaine de phosphorylation; En vert : domaine auto-inhibiteur (acides aminés 40 à 95) qui forme une longue hélice α Figure ci-dessous : Mécanisme proposé pour l'activation de la Ca2+-ATPase de Arabidopsis thaliana par la CaM.
Source : Tidow et al. (2012) La fixation de deux molécules de CaM complexée au calcium déplace l'hélice auto-inhibitrice du coeur catalytique, ce qui active la Ca2+-ATPase. 1er temps
2ème temps
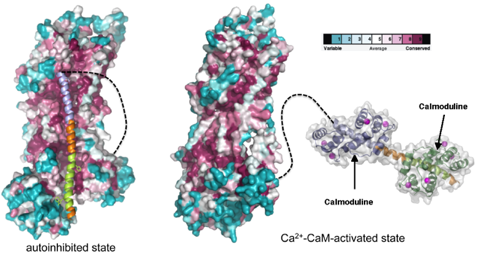
Figure ci-dessous : Structure du complexe [Ca2+-ATPase/CaM2] à une résolution de 1.95 Å. Les 2 sites de fixation de la CaM sont séparés par 8 acides aminés.
Source : Tidow et al. (2012)
|
|
Attention : il ne faut pas la confondre avec les FoF1-ATPases car elle fonctionne à l'envers des pompes ioniques.
Les ATP synthases sont des protéines oligomériques de masse molaire supérieure à 450 kDa et de structure très complexe : les types différents de chaînes polypeptidiques qui les composent sont au nombre de 8 chez E. coli, 8 ou 9 dans le chloroplaste, 10 ou plus dans la mitochondrie et au moins 13 pour celle du coeur de boeuf.
Source : Dimroth et al. (2006)
|
7. Transport actif secondaire La superfamille des facilitateurs majeurs ("Major Facilitator Superfamily" - MFS) est la plus grande famille de protéines de transport secondaire transmembranaire.
Ce transport actif est donc appelé secondaire parce que sa source d'énergie (le gradient électrochimique) doit être préalablement générée par un transport primaire. Voir la description thermodynamique des processus de transport, des aspects cinétiques et de la distribution des différentes conformation des transporteurs : Zhang et al. (2015) => voir les appendices. Exemple du transport du glucose
|
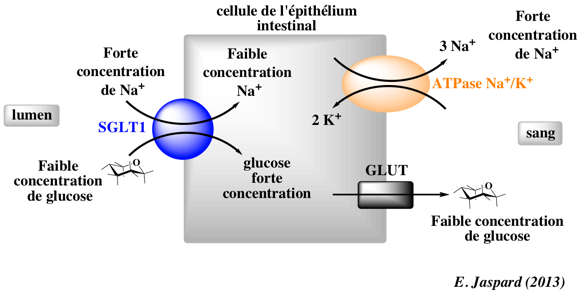
| Les symports [Na+/ glucose] |
Transporteur SGLT1 (gène SCL5A1)
Source : Wright et al. (2017) |
Transporteur SGLT2 (gène SCL5A2)
|
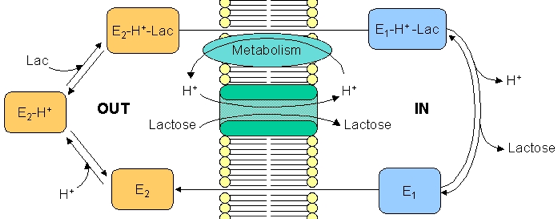
b. Description du cycle de la lactose perméase de Escherichia coli
Ce transporteur existe sous 2 formes :
Source : "Membrane III" La forme E1 n'est convertie en forme E2 que quand les sites de fixation du H+ et du lactose sont soit libres, soit occupés. 1. La forme E2 fixe le lactose. Quand la concentration extracellulaire du H+ augmente, la conformation du transporteur change : le site de fixation du lactose est alors orienté vers le milieu intracellulaire et devient un site à faible affinité (forme E1). Le lactose est alors relargué. 2. Les processus métaboliques diminuent la concentration du H+ : la forme E1, qui jusque là était ligandée au H+, relargue ce H+. Les 2 sites de fixation étant libres, l'enzyme re-bascule dans la forme E2. Le mécanisme peut de nouveau avoir lieu ce qui accroît la concentration intracellulaire du lactose. |
Visualisation de la lactose perméase de Escherichia coli complexée à un thiodigalactoside, à une résolution de 3,6 Å Code PDB : 1PV7 Seule la chaîne A est représentée. La structure peut prendre quelques secondes avant d'apparaître. |
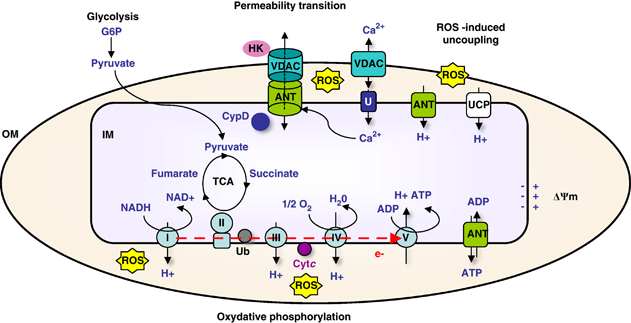
8. Les transports au travers des membranes mitochondriales Les canaux ioniques "Voltage-dependent anion-selective channel proteins" (VDAC) sont des porines de la membrane externe de la mitochondrie qui constituent la voie principale pour l'échange d'un grand nombre de métabolites. Il existe 3 isoformes de VDAC chez l'homme. Leur structure est en forme de tonneau β :
L'hexokinase, la glucokinase et la glycérol kinase (enzymes cytosoliques ATP-dépendantes) et la créatine kinase (enzyme mitochondriale) sont fixées à VDAC. De plus VDAC est un important régulateur du transport du Ca2+. En effet, Ca2+ est un co-facteur de la pyruvate déshydrogénase et de l'isocitrate déshydrogénase : la production énergétique et l'homéostasie énergétique sont donc toutes deux tributaires de la perméabilité de VDAC au Ca2+.
Source : Brenner et al. (2011) VDAC est associée à :
L'ensemble VDAC, ANT et CypD forme ce que l'on appelle le "Permeability Transition Pore complex". |
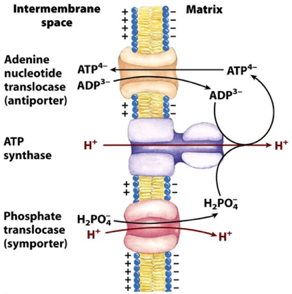
Les autres principaux transports sont :
Source : Lehninger Principles of Biochemistry - 5th edition (2008) - WH Freeman
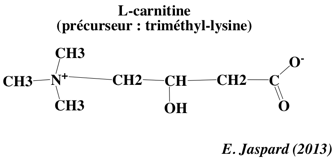
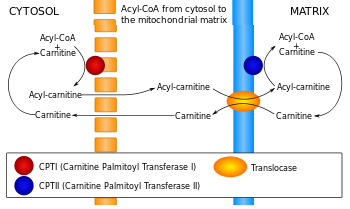
Le transport des acides gras au travers de la membrane interne de la mitochondrie est facilité par leur liaison à la carnitine. L'acide gras à transporter se fixe au coenzyme-A qui est remplacé par la carnitine pour former l'acyl-carnitine (figure ci-dessous). Cette réaction est catalysée par la carnitine O-palmitoyltransférase 1.
La carnitine est ensuite reconnue par un transporteur : la carnitine/acyl-carnitine translocase situé dans la membrane interne.
Source : Wikipédia Ce transporteur ("mitochondrial carnitine/acylcarnitine carrier protein") échange le complexe [acide gras-carnitine] pour une carnitine libre issue de l'intérieur de la mitochondrie. La carnitine est séparée de l'acide gras dans la matrice par la carnitine palmitoyltransférase 2 (E.C. 2.3.1.21). L'acide gras se fixe au coenzyme-A pour former un palmitoyl-CoA qui subit la β-oxydation dans la matrice. |
|
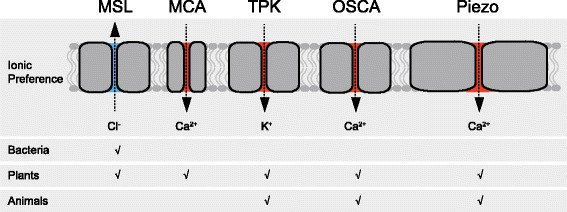
9. Détection du toucher et de la chaleur A. Patapoutian et D. Julius ont reçu le prix Nobel de médecine en 2021 "for their discoveries of receptors for temperature and touch". La capacité à détecter les forces physiques est le propre de tous les organismes : les cellules convertissent les stimuli mécaniques en signaux électriques ou chimiques via des canaux ioniques activés mécaniquement. L'identification de familles de canaux ioniques mécanosensibles a permis de grandes avancées dans le domaine de la mécanotransduction : plusieurs rôles mécanosensoriels tels que le toucher, la détection de la pression artérielle et l'ouïe sont maintenant liés aux mécanotransducteurs primaires. La figure ci-dessous présentent des homologues de canaux ioniques mécanosensibles chez les bactéries, les plantes et les animaux. La perméabilité principale aux ions et le flux prédominant d'ions sont indiqués.
Source : Hamant & Haswell (2017) MSL : "MscS-like"; MCA : "Mid1-Complementing Activity"; TPK : "Two Pore Potassium"; OSCA : "Reduced hyperosmolality-induced [Ca2+] increase"; canaux Piezo. |
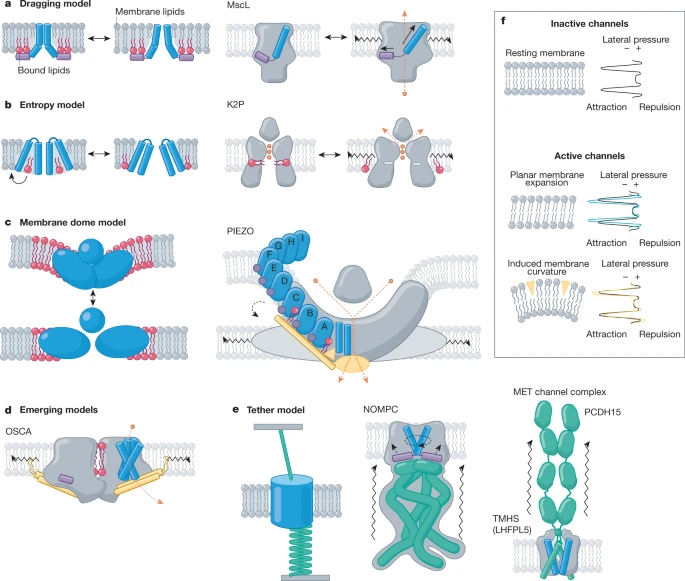
a. Modèles de mécanismes pour différentes familles de canaux ioniques mécanosensibles
Source : Kefauver et al. (2020) Code couleurs : hélices amphipathiques (violet); hélices transmembranaires (TM) (bleu); lipides liés (rouge); structures en forme de faisceau (or); attaches (émeraude); ions (orange); lipides membranaires (gris). Modèle a
Modèle b
Modèle c
Modèle d
Modèle e
|
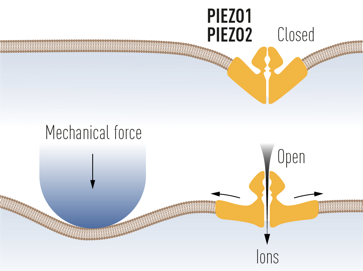
b. Détection du toucher A. Patapoutian et ses collaborateurs ont démontré que le canal ionique Piezo2 ("Piezo-type mechanosensitive ion channel component 2") est essentiel pour le sens du toucher et dans la détection de la position et du mouvement du corps - propriété appelée proprioception.
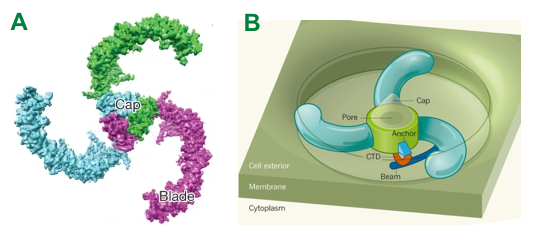
Source : The Nobel Prize in Physiology or Medicine 2021 La structure de Piezo2 de la souris a été déterminée par cryomicroscopie électronique :
Source : A : Wang et al. (2019) - B : Jan & Jan (2018) |
|
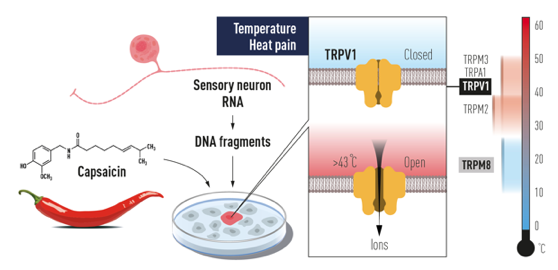
c. Détection de la chaleur D. Julius et ses collaborateurs ont utilisé la capsaïcine (8-méthyl-N-vanillyl-6-nonénamide) de piment. C'est un capsaïcinoïde analgésique non narcotique (métabolite secondaire).
Source : The Nobel Prize in Physiology or Medicine 2021 Par des approches indépendantes, D. Julius et A. Patapoutian ont utilisé le menthol pour identifier le récepteur TRPM8 ("Transient Receptor Potential cation channel subfamily M member 8") qui est activé par le froid, le menthol, l'eucalyptol et l'iciline. |
d. La superfamille des canaux cationiques TRP La superfamille des canaux cationiques TRP ("Transient Receptor Potential") représente une diversité de mécanismes d'activation et de sélectivités plus importante que tout autre famille de canaux ioniques. Cette superfamille est divisée en 7 sous-familles :
Les canaux TRP sont des tétramères. Les sous-unités possèdent 6 domaines transmembranaires qui forment des pores sélectifs pour les cations, qui, dans plusieurs cas, présentent une perméabilité élevée au calcium. Le transport catalysé est : Ca2+extracellulaire <=>Ca2+intracellulaire |
| 10. Liens Internet et références bibliographiques |
|
"Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X "Carrier mediated transport through biomembranes " - Ranjan et al. (2011) Voir des animations des différents types de transport Animations et quizz : "Biology, Eighth Edition (Raven)" |
|
|
Bases de données "Transporter Classification database" "TransportDB : Relational database describing the predicted cytoplasmic membrane transport protein complement for organisms whose complete genome sequence are available" "OPM : Orientations of Proteins in Membranes database" "Ligand-Gated Ion Channel database" |
|
Articles fondateurs de la thermodynamique des mécanismes de transports biologiques Goldman D.E. (1943) "Potential, impedance, and rectification in membranes" J. Gen. Physiol. 27, 37 - 60 Teorell, T. (1948) "Membrane electrophoresis in relation to bio-electrical polarization effects" Nature 162, 961 Ussing H. H. (1949) "Transport of ions across cellular membrane" Physiol. Rev. 29, 127 - 155 Hodgkin & Katz (1949) "The effect of sodium ions on the electrical activity of giant axon of the squid" J. Physiol. 108, 37 - 77 Hodgkin, Huxley & Katz (1952) "Measurement of current-voltage relations in the membrane of the giant axon of Loligo" J. Physiol. 116, 424 - 448 Singer & Nicolson (1972) "The fluid mosaic model of the structure of cell membranes" Science 175, 720 - 731 |
|
|
Busch & Saier (2002) "The Transporter Classification (TC) System, 2002" Critic. Rev. Biochem. Mol. Biol. 37, 287 - 337 Forrest et al. (2011) "The structural basis of secondary active transport mechanisms" Biochim. Biophys. Acta 1807, 167 - 188 Sun et al. (2012) "Crystal structure of a bacterial homologue of glucose transporters GLUT1–4" Nature 490, 361 - 366 |
|
|
"Transport Membranaire" - Alex J. Baertschi Huang & Czech (2007) "The GLUT4 glucose transporter" Cell Metab. 5, 237 - 252 Goodenough & Paul (2009) "Gap Junctions" Cold Spring Harb. Perspect. Biol. 1, a002576. |
|
|
Dimroth et al. (2006) "Catalytic and mechanical cycles in F-ATP synthases" EMBO reports 7, 276 - 282 Armen et al. (2007) "Inventing the dynamo machine: the evolution of the F-type and V-type ATPases" Nat. Rev. Microbiol. 5, 892 - 899 Brenner et al. (2011) "Adenine nucleotide translocase family: four isoforms for apoptosis modulation in cancer" Oncogene 30, 883 - 895 Tidow et al. (2012) "A bimodular mechanism of calcium control in eukaryotes" Nature 491, 468 - 472 |
|
|
Mohan et al. (2010) "Molecular Dynamics Simulation Studies of GLUT4: Substrate-Free and Substrate-Induced Dynamics and ATP-Mediated Glucose Transport Inhibition" PLoS One 5, e14217 Ojuka et al. (2012) "The role of CaMKII in regulating GLUT4 expression in skeletal muscle" Am. J. Physiol. Endocrinol. Metab. 303, E322 - E331 Arai et al. (2014) "A molecular fluorescent probe for targeted visualization of temperature at the endoplasmic reticulum" Sci. Rep. 4, 6701 |
|
|
Arai et al. (2015) "Mitochondria-targeted fluorescent thermometer monitors intracellular temperature gradient" Chem. Commun. 51, 8044 - 8047 Nomura et al. (2015) "Structure and mechanism of the mammalian fructose transporter GLUT5" Nature 526, 397 - 401 Zhang et al. (2015) "Energy coupling mechanisms of MFS transporters" Protein Sci. 24, 1560 - 1579 |
|
|
Alvarez & Latorre (2017) "The enduring legacy of the "constant-field equation" in membrane ion transport" J. Gen. Physiol. 149, 911 - 920 Wright et al. (2017) "Novel and Unexpected Functions of SGLTs" Physiology (Bethesda) 32, 435 - 443 Myers et al. (2018) "Structure of native lens connexin 46/50 intercellular channels by cryo-EM" Nature 564, 372 - 377 |
|
|
Steinhorst & Kudla (2019) "How plants perceive salt" Nature 572, 318 - 320 Jiang et al. (2019) "Plant cell-surface GIPC sphingolipids sense salt to trigger Ca2+ influx" Nature 572, 341 - 346 |
|
|
Althoff & Abramson (2020) "Protein structure reveals how a malaria parasite imports a wide range of sugars" Nature, News and views Qureshi et al. (2020) "The molecular basis for sugar import in malaria parasites" Nature 578, 321 - 325 Shimomura et al. (2020) "A native prokaryotic voltage-dependent calcium channel with a novel selectivity filter sequence" Elife 9, e52828 Drew et al. (2021) "Structures and General Transport Mechanisms by the Major Facilitator Superfamily (MFS)" Chem. Rev. 121, 5289 - 5335 |
|
|
Détection du toucher et de la chaleur Prix Nobel de médecine 2021 "for the discoveries of receptors for temperature and touch" Caterina et al. (1997) "The capsaicin receptor: a heat-activated ion channel in the pain pathway" Nature 389, 816 - 824 Peier et al. (2002) "A TRP channel that senses cold stimuli and menthol" Cell 108, 705 - 715 Bertrand Coste et al. (2010) "Piezo1 and Piezo2 are essential components of distinct mechanically-activated cation channels" Science 330, 55 - 60 Hamant & Haswell (2017) "Life behind the wall: sensing mechanical cues in plants" BMC Biol. 15, 59 Jan & Jan (2018) "Force-activated ion channels in close-up" Nature 573, 225 - 229 Wang et al. (2019) "Structure and mechanogating of the mammalian tactile channel PIEZO2" Nature 573, 225 - 229 Douguet & Honoré (2019) "Mammalian Mechanoelectrical Transduction: Structure and Function of Force-Gated Ion Channels" Cell 179, 340 - 354 Kefauver et al. (2020) "Discoveries in structure and physiology of mechanically activated ion channels" Nature 587, 567 - 576 |
|
![]()