| HMG-CoA réductase, le cholestérol et les statines |
| Tweet |
|
|
1. Présentation générale 2. La HMG-CoA réductase ou HMGCR 3. Régulation de l'HMGCR |
4. Le cholestérol et les lipoprotéines 5. Les statines : inhibiteurs de l'HMGCR 6. Liens Internet et références bibliographiques |
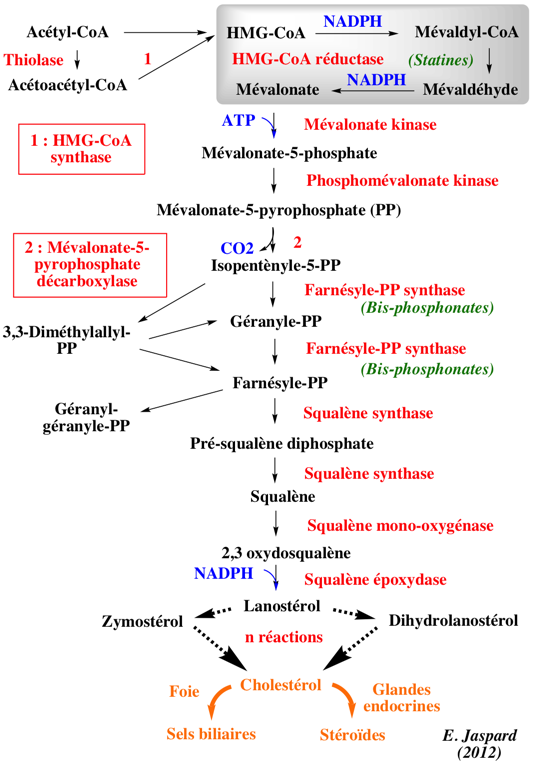
1. Présentation générale La biosynthèse de l'isopentènyle diphosphate (isopentènyle pyrophosphate), le précurseur des isoprénoïdes chez toutes les formes de vie s'effectue via 2 voies métaboliques : la voie du mévalonate et la voie du glycéraldéhyde 3-phosphate/pyruvate (appelée aussi voie non-mévalonate).
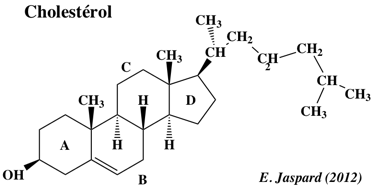
La HMG-CoA réductase (ou 3-Hydroxy-3-Méthyl-Glutaryl-CoA réductase ou HMGCR) est l'enzyme qui contrôle la voie du mévalonate qui, entre autres devenirs, mène à la biosynthèse du choléstérol et d'autres isoprénoïdes.
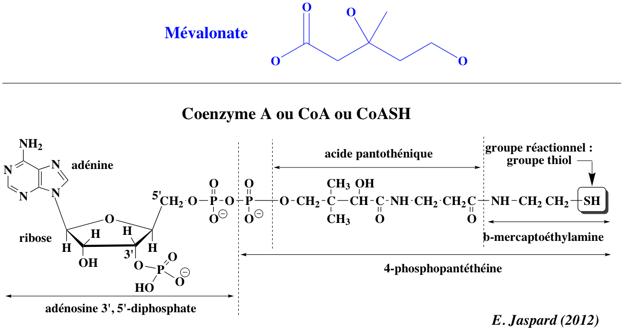
CoA : coenzyme A ou CoASH (voir ci-dessous).
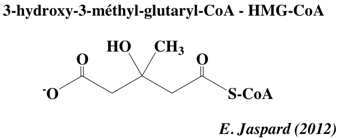
L'essentiel du cholestérol dans la circulation n'entre pas directement dans le système sanguin à partir de l'alimentation. Le cholestérol est surtout synthétisé dans le réticulum endoplasmique par un ensemble de réactions dont l'étape limitante est la réaction catalysée par l'HMGCR. La réaction catalysée par l'HMGCR est : (R)-mévalonate + CoA + 2 NAD(P)+ ==> 3-hydroxy-3-méthyl-glutaryl-CoA + 2 NAD(P)H
L'un des modes de bloquage de la biosynthèse du cholestérol est donc l'inhibition de l'HMGCR. Le métabolisme du cholestérol est également hautement régulé au niveau post- transcriptionnel par plusieurs miRNA. Il existe d'autres processus d'élimination du cholestérol :
|
2. La HMG-CoA réductase ou HMGCR L'HMGCR est l'enzyme qui contrôle la voie du mévalonate donc la synthèse du cholestérol. Il existe 2 classes d'HMGCR (PFAM PF00368) :
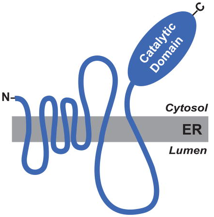
Chez l'homme, l'HMGCR est un tétramère (en fait un dimère de dimère) de 888 acides aminés (97,5 kDa). C'est une glycoprotéine transmembranaire (réticulum endoplasmique et peroxysome). Elle contient :
Source : Jo & DeBose-Boyd (2010)
La structure de l'interface hydrophobe entre les 2 dimères est stabilisée par un pont salin entre Arg641 et Glu782 et des liaisons hydrogène entre les Glu700 des 2 monomères. L'HMGCR de classe II ne possède pas le domaine N-terminal riche en hélices. Le domaine catalytique est organisé en une grande région L et une petite région S. |
|
Visualisation de la HMG-CoA réductase de Homo sapiens à une résolution de 2,1 Å. PDB : 1DQ8 Domaine catalytique complexé au coenzyme A et au HMG (3-Hydroxy-3-Méthyl-Glutarate). Seule la chaîne A est représentée. 3 acides aminés impliqués dans la catalyse sont indiqués. His 866 n'apparaît pas.
|
|
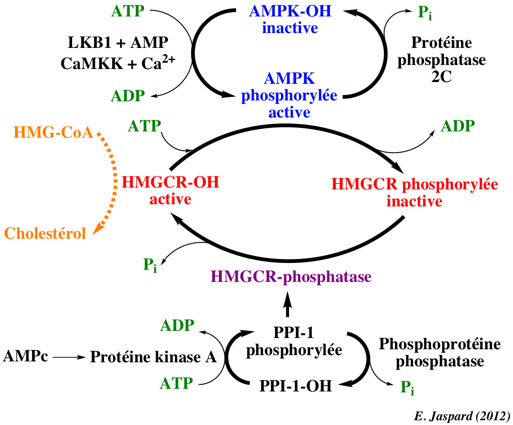
L'HMGCR est une enzyme particulièrement régulée : a. La régulation de la transcription : le gène de l'HMGCR est activé par la "sterol regulatory element binding protein", protéine qui se fixe sur le promoteur du gène de l'HMGCR quand le taux du cholestérol chute. b. La traduction de l'ARM messager codant l'HMGCR est inhibée par le farnesol, un dérivé de la voie du mévalonate. c. La dégradation de l'HMGCR : quand le taux des stérols augmente, l'HMGCR est davantage sujette à la dégradation médiée par le réticulum endoplasmique ("ER-associated degradation").
d. Enfin l'activité de l'HMGCR est régulée par phosphorylation de Ser872 par la protéine kinase activée par l'AMP (AMPK), quand la charge énergétique de la cellule est faible et la concentration en AMP est élevée. Ser872 est située à proximité du groupement phosphate du NADP : sa phosphorylation diminue donc l'affinité pour le NADPH, ce qui diminue la synthèse du mévaldyl-CoA à partir du HMG-CoA (voir la figure de la synthèse du cholestérol en haut de cette page).
Des hormones comme le glucagon et l'adrénaline (ou épinephrine) inhibe la biosynthèse du cholestérol en activant l'inhibiteur de PPI-1. Inversement, l'insuline stimule la déphosphorylation et active l'HMGCR. |
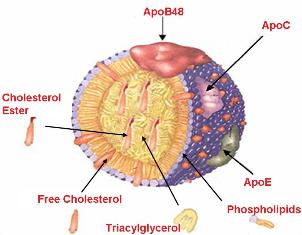
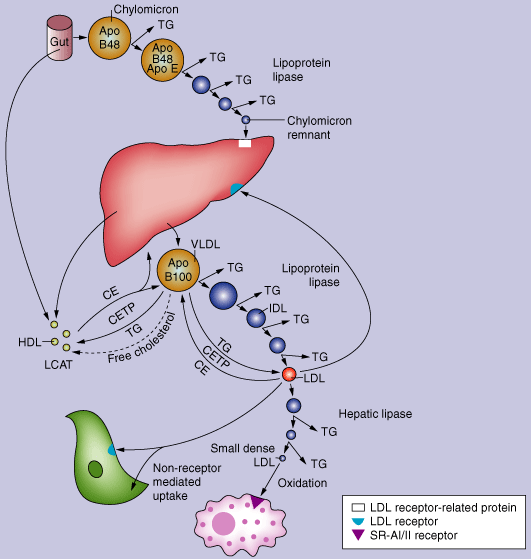
4. Le cholestérol et les lipoprotéines Le cholestérol est transporté dans le plasma essentiellement sous la forme d'esters du cholestérol associés aux lipoprotéines.
Les lipoprotéines sont des vésicules qui transportent les graisses dans le plasma sanguin. Elles ont une structure globulaire et sont constituées de :
Source : "The LaDu Research Lab"
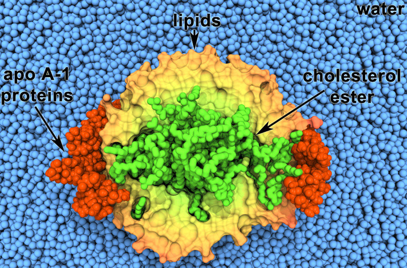
Les apoliproptéines peuvent traverser les lipoprotéines. Certaines ont un rôle structural d'autres sont des régulateurs du métabolisme. Il existe 6 classes d'apoliproptéines (A à E, H), certaines divisées en sous-classes. Les charges portées par le cholestérol libre et les phospholipides en surface sont orientées vers les molécules d'eau externes : la surface d'une lipoprotéine ressemble donc à la partie extra-cellulaire d'une membrane.
Source : "HDL and Nanodiscs" |
| Composition des principaux complexes liporotéiques | ||||||||||
| Complexe | Source | Diamètre (nm) | Densité (g/ml) | Protéines (%) | Apolipoprotéines | % TG | % PL | % CE | % CL | % AGL |
| Chylomicron | Intestin | > 75 | < 0,95 | 1 - 2 | acquièrent l'apoE et l'apoC des HDL en leur cédant leur apoA1 / apoB-48 / apoC / apoE | 85 - 88 | 8 | 3 | 1 | 0 |
| VLDL | Foie | 30 - 70 | 0,95 à 1,006 | 7 - 10 | apoB-100 / apoC / apoE | 50 - 55 | 18 - 20 | 12 - 15 | 8 - 10 | 1 |
| IDL | VLDL | 25 - 50 | 1,006 à 1,019 | 10 - 12 | apoB-100 / apoE | 25 - 30 | 25 - 27 | 32 - 35 | 8 - 10 | 1 |
| LDL | VLDL | 18 - 25 | 1,019 à 1,063 | 20 - 22 | apoB-100 / vitamine E et caroténoïdes | 10 - 15 | 20 - 28 | 37 - 48 | 8 - 10 | 1 |
| HDL2 | Intestin (entérocytes), foie (chylomicron & VLDL) | 6 - 10 | 1,063 à 1,125 | 33 - 35 | apoA / apoC / apoE | 5 - 15 | 32 - 43 | 20 - 30 | 5 - 10 | 0 |
| HDL3 | Intestin (entérocytes), foie (chylomicron & VLDL) | 1,125 à 1,21 | 55 - 57 | 3 - 13 | 26 - 46 | 15 - 30 | 2 - 6 | 6 | ||
| Albumine - AGL | Tissus adipeux | --- | > 1,281 | 99 | ------- | 0 | 0 | 0 | 0 | 100 |
TG : triacylglycérols - PL : phospholipides - CE : cholesteryl esters - CL : cholestérol libre - AGL : acides gras libres - IDL : "Intermediate Density Lipoprotein". Sources : "Lipoproteins" - Wikipedia &"Lipoproteins" - M.W. King |
||||||||||
Le foie synthétise les lipoprotéines VLDL ("Very Low Density Lipoprotein", lipoprotéine de très basse densité) qui sont converties en LDL ("Low Density Lipoprotein") via la lipoprotéine lipase associée aux cellules endothéliales ("endothelial cell-associated lipoprotein lipase"). Le cholestérol synthétisé par le foie, de même que tout régime riche en cholestérol excèdant les besoins hépatiques, est transporté dans le sérum au sein des vésicules de lipoprotéines LDL. Les lipoprotéines LDL se fixent sur des récepteurs spécifiques. Puis la vésicule s'invagine en "réceptosomes" dirigés vers les lysosomes avec lesquels ils fusionnent. Les enzymes du lysosome libèrent les acides gras, le cholestérol (incorporé dans le réticulum endoplasmique) et les récepteurs (hydrolysés en acides aminés). Le cholestérol de la membrane plasmique peut-être extrait par les lipoprotéines HDL et esterifié par LCAT, l'enzyme associée aux lipoprotéines HDL. les lipoprotéines HDL sont prélevés par les hépatocytes et se retrouvent dans les sels biliaires. Le cholestérol issu des tissus périphériques via les lipoprotéines HDL peut être transferés aux lipoprotéines VLDL et LDL via la protéine de transfert des esters de cholestérol ("cholesteryl ester transfer protein apo-D" - CETP) associée aux lipoprotéines HDL.
Source : "Chylomicron Metabolism" Le transport réverse du cholestérol permet un retour du cholestérol périphérique vers le via les lipoprotéines LDL. Finalement, le cholestérol est excrété dans la bile sous forme de cholestérol libre ou de sels biliaires après conversion dans le foie. |
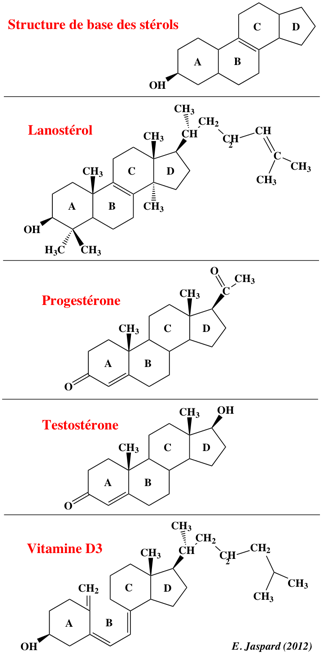
Les stérols sont des lipides dérivant du noyau cyclopentanophénanthrénique. Les différentes types de stérols se distinguent selon le nombre et la position d'insaturation et/ou de chaînes latérales.
Vitamine D3 : cholécalciférol. |
| Sous-classes des stérols | Sous-classes des stéroïdes | Sous-classes des secostéroïdes | Sous-classes des acides biliaires | Sous-classes des stéroïdes conjugués |
| cholestérol et ses dérivés | stéroïdes à 18C : oestrogènes et leurs dérivés | vitamines D2 à D7 et leurs dérivés (vitamine D2 : ergocalciférol) |
acides biliaires de 22 à 29 carbones, alcools et leurs dérivés | stéroïdes glucuronidés |
| cholestéryl esters | stéroïdes à 19C : androgènes et leurs dérivés | stéroïdes sulfatés | ||
| phytostérols et leurs dérivés | stéroïdes à 21C : gluco-corticoïdes, minéralo-corticoïdes, progestogines et leurs dérivés | stéroïdes conjugués à la glycine | ||
| stérols marins et leurs dérivés | stéroïdes conjugués à la taurine | |||
| stérols fongiques et leurs dérivés | ||||
| Source : "Classification des lipides" - Wikipédia | ||||
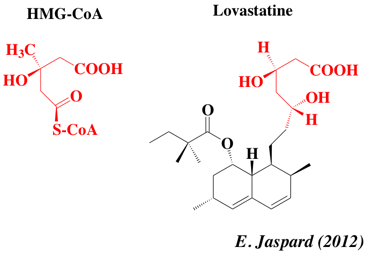
5. Les statines : inhibiteurs de l'HMGCR Ce sont des agents hypo-lipidèmiques, inhibiteurs de l'HMGCR. Ils ont été découverts par Akira Endo en 1971. L'inhibition de l'HMGCR dans le foie stimule les récepteurs du LDL, ce qui augmente l'élimination du LDL du système sanguin et une diminution du taux de cholestérol. Ces drogues incluent : la rosuvastatine (Crestor), la lovastatine (monacoline K - Mevacor), l'atorvastatine (Lipitor), la pravastatine (Pravachol), la fluvastatine (Lescol), la pitavastatine (Livalo), la simvastatine (Zocor), la cerivastatine, la vytorine qui combine la simvastatine et l'ezetimibe. Ces molécules sont des inhibiteurs compétitifs du HMG-CoA. Il y a une diminution de la synthèse du mévalonate. Par exemple, le noyau lactone de la simvastatine est hydrolysé et génère l'acide β, δ- dihydroxy, métabolite analogue structural du HMG-CoA. Figure ci-dessous : comparaison des structures du HMG-CoA et de la lovastatine.
Les statines et l'HMGCR ont une importance économique considérable dans l'industrie pharmaceutique. Celà explique les efforts concernant la résolution de structures tridimensionnelles de nombreux complexes entre cette enzyme et ces inhibiteurs : on dénombre une vingtaine de structures de complexes entre l'HMGCR et des statines. Voir le chapitre "Medical Implications for HMGR" ("HMG-CoA Reductase" - Proteopedia). |
| 6. Liens Internet et références bibliographiques | |
|
"Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X |
|
|
"HMG-CoA Reductase" - Proteopedia
"Proteopedia : the free, collaborative 3D-encyclopedia of proteins & other molecules" |
|
|
Voie des statines (interactive) Métabolisme des lipoprotéines Classification des lipides |
|
|
Friesen & Rodwell (2004) "The 3-hydroxy-3-methylglutaryl coenzyme-A (HMG-CoA) reductases" Genome Biology, 5, 248 Jo & DeBose-Boyd (2010) "Control of Cholesterol Synthesis through Regulated ER-Associated Degradation of HMG CoA Reductase" Crit Rev Biochem Mol Biol 45, 185 - 198 |
|
|
"DrugBank" "MetaCyc Pathway : mévalonate pathway I" "HDL and Nanodiscs" |
|
![]()