| Rôle de GLUT4, de l'insuline et du glucagon dans la régulation de la glycolyse |
| Tweet |
|
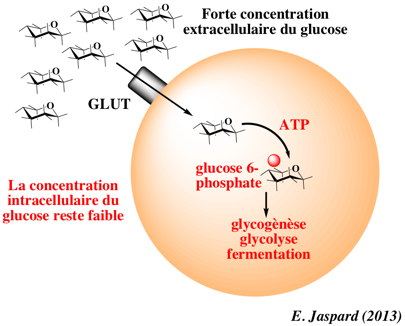
La contraction et l'insuline sont les stimuli majeurs qui activent le transport du glucose dans les muscles squelettiques. La mobilisation du glucose par les muscles squelettiques représente environ 70% du glucose prélevé au sérum. Ce processus est donc extrêmement important dans l'homéostasie du glucose. L'aptitude à transporter le glucose au travers de la membrane plasmique est une caractéristique commune à quasiment tous les types de cellules, de la simple bactérie à la cellule neuronale hautement spécialisée. Il existe 2 types de transport du glucose. a. Transport facilité du glucose Il est assuré par des membres de la famille des transporteurs GLUT ("GLUcose Transporter") qui forment un pore au travers de la membrane (uniport).
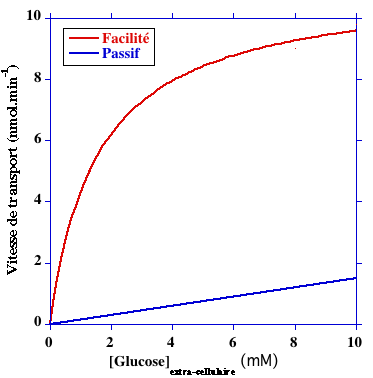
Certains oses entrent via ce type de pore par diffusion passive, c'est-à-dire sans apport d'énergie (ce type de transport s'effectue dans le sens du gradient de concentration ou gradient électrochimique). Le transport d'un soluté par diffusion facilitée augmente considérablement la vitesse de transport de ce soluté. Par ailleurs , KMglucose GLUT1 ≈ 1,5 mM (figure ci-dessous).
|
| Les transporteurs d'oses - GLUT : 12 hélices α transmembranaires. |
| GLUT1 (gène SLC2A1) : exprimé dans presque tous les tissus (mais à un faible niveau dans le foie et les muscles squelettiques). C'est le principal transporteur du glucose dans les érythrocytes. KMglucose = 3 - 7 mM. |
| GLUT2 (gène SCL2A2) : exprimé essentiellement dans le foie, les cellules pancréatiques β, les reins et les intestins. KMglucose = 15 - 20 mM. Il transporte aussi le fructose et le galactose. GLUT2 assure le transport bi-directionnel du glucose au travers de la membrane. En d'autres termes, c'est le transporteur qui permet au glucose de sortir de la cellule après qu'il ait été déphosphorylé par la glucose-6 phosphatase (enzyme de la néoglucogénèse). GLUT2 de l'homme est une protéine de 524 acides aminés avec environ 56% d'identité avec GLUT1 de l'homme. |
| GLUT3 (gène SCL2A3) : exprimé essentiellement dans les neurones. De tous les transporteurs du glucose, c'est celui qui a le KM le plus faible. |
| GLUT4 (gène SCL2A4) : exprimé essentiellement dans les tissus insulino-dépendants comme les tissus adipeux et les muscles squelettiques. KMglucose = 5 mM. Voir un développement ci-dessous. |
| GLUT5 (gène SCL2A5) : exprimé essentiellement dans les entérocytes et les intestins. Il transporte spécifiquement le fructose. |
| GLUT6 (gène SCL2A6) : exprimé essentiellement dans le cerveau et les leukocytes. Il transporte le glucose. Attention : il a aussi comme appellation GLUT9 mais il est différent du suivant. |
| GLUT9 (gène SCL2A9) : exprimé essentiellement dans les reins, le foie et le placenta. Il transporte le glucose à une vitesse faible. Il transporte aussi l'urée et le fructose. |
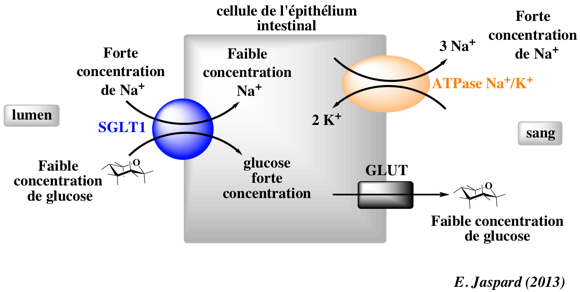
b. Transport actif secondaire Il s'effectue via les symports [Na+ / glucose] ("Sodium/Glucose Co-Transporter" - SGLT1 et SGLT2).
Ces symports utilisent le gradient transmembranaire de Na+généré par la pompe à sodium ou [Na+/K+]-ATPase constitué de 12 hélices α transmembranaires. |
| Les symports [Na+/ glucose] |
| SGLT1 (gène SCL5A1) : symport [Na+ / glucose] ("Sodium/Glucose Co-Transporter 1"). Le rapport du transport est 1 molécule de glucose pour 2 ions Na+. Il contient 11 hélices α transmembranaires. |
| SGLT2 (gène SCL5A2) : symport [Na+ / glucose] ("Sodium/Glucose Co-Transporter 2"). Le rapport du transport est 1 molécule de glucose pour 1 ion Na+. Il contient 14 hélices α transmembranaires. |
|
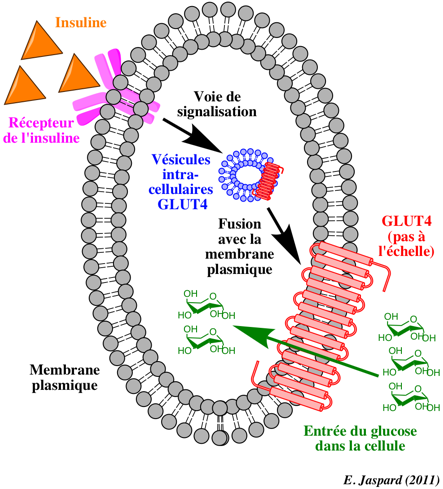
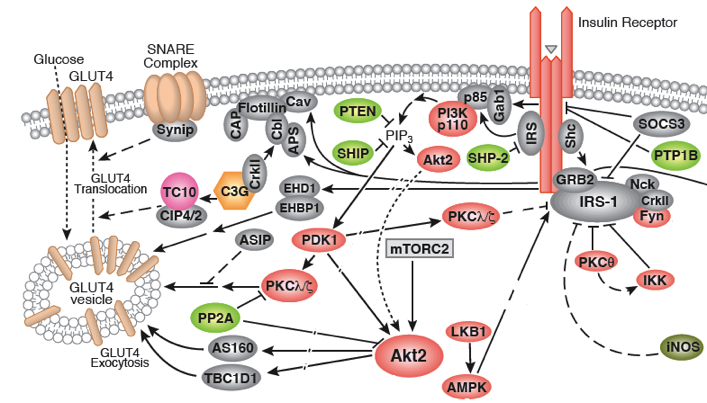
Rôle de l'insuline et transporteur GLUT4 Dans une cellule non stimulée ou quand la concentration en insuline est faible, le transporteur insulino-dépendant du glucose GLUT4 est localisé dans des vésicules de stockage des cellules hépatiques et musculaires. Ces vésicules doivent fusionner avec la membrane plasmique avant que le glucose ne pénètre dans la cellule.
Quand le niveau de glucose circulant est élevé, l'insuline est libérée par les ilots de Langerhans et elle facilite la mobilisation du glucose via une augmentation de la synthèse et de la translocation de GLUT4 des compartiments endosomiques vers la membrane plasmique. |
|
En conséquence, l'absorption du glucose augmente.
Source : Cell Signaling |
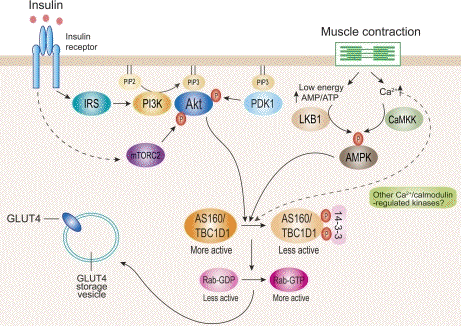
Source : Sakamoto & Holman (2008)
A la surface de la cellule, GLUT4 facilite la diffusion du glucose dans le sens de son gradient de concentration dans les cellules du foie et du muscle. Une fois dans les cellules, le glucose est rapidement phosphorylé par la glucokinase dans le foie et par l'hexokinase dans les autres tissus pour former le glucose-6-phosphate qui alimente la glycolyse ou qui est polymerisé en glycogène. Le glucose-6-phosphate ne peut pas rétro-diffuser à l'extérieur des cellules, ce qui maintient aussi le gradient de concentration du glucose libre pour qu'il puisse diffuser passivement dans les cellules. |
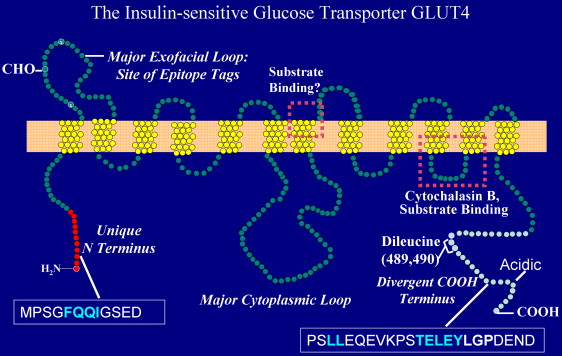
Structure de GLUT4 La protéine GLUT4 (Uniprot : P14672) est l'une des nombreuses isoformes qui constituent une famille de 14 transporteurs d'hexoses chez l'homme (GLUT1 à GLUT12, GLUT14, et HMIT) subdivisés en 3 classes. La chaîne polypeptidique de GLUT4 contient 509 acides aminés qui forment 12 hélices transmembranaires. Le domaine N-terminal contient les hélices I à VI et le domaine C-terminal contient les hélices VII à XII. Ces 2 régions forment un pseudo-axe de symmétrie autour d'un tunnel central polaire qui laisse entrer/sortir le glucose. Ce tunnel de nature amphipathique semble formé par les hélices 1, 2, 4, 5, 7, 8, 10 et 11. Les séquences des domaines cytoplasmiques N-terminal et C-terminal sont particulières et expliquent en grande partie l'aptitude au traffic transmembranaire de ce transporteur.
Source : Huang & Czech (2007)
Ces motifs dirigent probablement les aspects cinétiques de l'endocytose et de l'exocytose dans un systéme continu de traffic de recyclage. Les deux leucines et les motifs acides du domaine C-terminal sont trouvés également dans une aminopeptidase ("insulin-regulated aminopeptidase" - IRAP) qui, de manière semblable dans les adipocytes, est séquestrée dans des membranes intracellulaires enrichies en GLUT4 et sujette à l'action de l'insuline. |
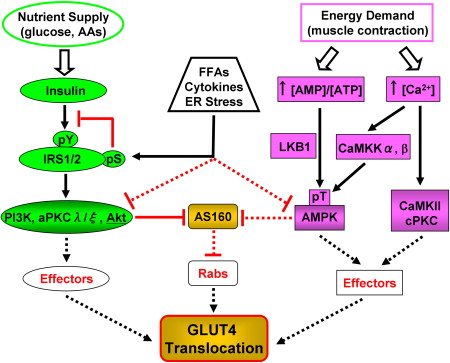
Convergence des voies de signalisation initiées par l'insuline et par l'exercice conduisant à la translocation de GLUT4 La signalisation de l'insuline via la voie PI3-kinase (PI3K) et la contraction du muscle via un rapport élevé AMP/ATP et une concentration intracellulaire élevée de Ca2+ conduit à l'activation de protèines kinases qui agissent en aval : la CaM kinase II, Akt, des protéines kinases C isoformes λ et ζ, la protéine kinase activée par l'AMP. Ces protèines kinases phosphorylent des effecteurs qui modulent les étapes de la voie du traffic de GLUT4. AS160 est l'un de ces effecteurs qui régule négativement l'une des étapes précoces de l'exocytose de GLUT4. La régulation négative de ces voies par les acides gras, les cytokines et la réponse au stress du réticulum endoplasmique sont observées dans l'obésité et les diabètes, ce qui contribue à la résistance à l'insuline.
Source : Huang & Czech (2007) Les lignes pointillées impliquent des voies hypothètiques non encore mises en évidence expérimentalement. |
Effets de l'insuline Puisque le glucose pénètre dans le muscle et le tissu adipeux par l'intermédiaire du transporteur GLUT4 qui est contrôlé par l'insuline, une carence en insuline (diabète insulino dépendant) a pour conséquence une élévation de la concentration du glucose sanguin.
|
Régulation de la phosphofructokinase-2/fructose 2,6-bisphosphatase-2 (PFK-2/FBPase-2) par le glucagon Dans le foie, la PFK-2 est sous le contrôle du glucagon, une hormone produite par le pancréas quand le taux de glucose sanguin baisse. Quand la concentration du glucagon augmente, la protéine kinase AMP cyclique dépendante (PKA) phosphoryle une sérine ou une thréonine de la [PFK-2 / FBPase-2]. Exemples : foie de rat : Ser 32 - coeur de boeuf : Ser 84 - levure : Thr 157. La déphosphorylation est catalysée par la protéine phosphatase 2A xylulose-5-phosphate-dépendante qui est activée par le glucose.
La phosphorylation :
Il en résulte une baisse de concentration du fructose 2,6-bisphosphate et la phosphofructokinase-1 de la glycolyse est moins activée. En conséquence, la glycolyse est ralentie quand la concentration du glucagon augmente après une baisse du taux de glucose sanguin. |
La diminution d'insuline augmente les effets du glucagon
Les changements du métabolisme induits par le glucagon favorisent la néoglucogénèse en défaveur de la glycolyse et entraînent une hyperglycémie. |
| Liens Internet et références bibliographiques |
| Huang & Czech (2007) "The GLUT4 Glucose Transporter" Cell Metab. 5, 237 - 252 | Article |
| Mohan et al. (2010) "Molecular Dynamics Simulation Studies of GLUT4: Substrate-Free and Substrate-Induced Dynamics and ATP-Mediated Glucose Transport Inhibition" PLoS ONE 5, e14217 | Article |
|
Sakamoto & Holman (2008) "Emerging role for AS160/TBC1D4 and TBC1D1 in the regulation of GLUT4 traffic" Am. J. Physiol. Endocrinol. Metab. 295, E29 - 37 Stöckli et al. (2008) "Regulation of Glucose Transporter 4 Translocation by the Rab Guanosine Triphosphatase-Activating Protein AS160/TBC1D4: Role of Phosphorylation and Membrane Association" Mol. Endocrinol. 22, 2703 - 2715 Sheena et al. (2011) "Elucidation of the Glucose Transport Pathway in Glucose Transporter 4 via Steered Molecular Dynamics Simulations" PLoS ONE 6, e25747 |
|
![]()