| Régulation du métabolisme : introduction |
| Tweet |
|
|
1. Vue globale du métabolisme et de sa régulation 2. Les signaux extracellulaires et intracellulaires 3. La fonction des protéines et l'activité des enzymes 4. La concentration des métabolites |
5. La quantité d'enzymes synthétisées 6. Les moyens d'étude de la régulation du métabolisme 7. Liens Internet et références bibliographiques |
1. Vue globale du métabolisme et de sa régulation Toute cellule est le siège de milliers de réactions biochimiques. Cet ensemble de réactions s'appelle le métabolisme. Les réactions forment un réseau de voies très ramifiées le long desquelles les molécules, que l'on appelle des métabolites, sont transformées.
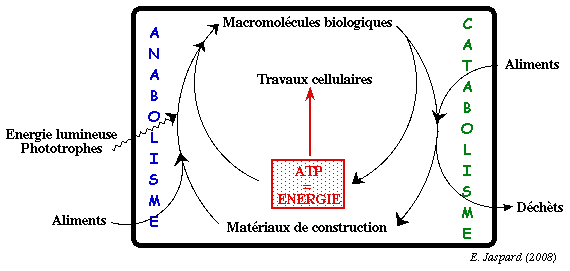
L'ensemble de ces réactions se déroulent à une très grande vitesse, bien supérieure à celles qu'elles auraient isolément dans la nature, grâce à des catalyseurs biologiques que sont les enzymes. Certaines voies métaboliques libèrent de l'énergie en décomposant des molécules de structure élaborée en composants élémentaires de structure plus simples pour finir par l'oxydation complète des biomolécules en CO2 et H2O. Cet ensemble de processus de dégradation s'appelle le catabolisme. A l'inverse, l'énergie libérée au cours des processus cataboliques est utilisée pour fabriquer un très large ensemble de molécules complexes à partir de quelques précurseurs simples. Cet ensemble de réactions de biosynthèse s'appelle l'anabolisme.
L'environnement des organismes et des cellules change en permanence. En conséquences, les réactions du métabolisme doivent donc être finement régulées pour :
|
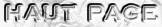
|
La relation structure - fonction des protéines Cet ensemble de réactions fait appel à des protéines (enzymes ou non enzymes). Si le métabolisme est une merveille de fonctionnement coordonné de millions de molécules c'est, entre autre, parce que les protéines ont une structure idéalement adaptée à chacun de leur(s) partenaire(s), structure dont découle leur(s) fonction(s). De manière générale, les protéines sont :
La régulation du métabolisme implique une succession de trans-conformations des protéines qui contrôlent cette régulation : pour la plupart, ces protéines passent en permanence d'une conformation non active à une conformation active et inversement. Les exemples typiques sont les cascades de phosphorylation ou la fixation de ligands (calmoduline, protéines 14.3.3, messagers secondaires ...). La relation structure - fonction des protéines, c'est-à-dire le lien entre la structure (ou l'absence de structure) d'une protéine et sa/ses fonction(s) dans la cellule est donc un aspect important de la régulation du métabolisme. Avec l'avènement des domaines en "omique", la régulation du métabolisme est intégrée dans un ensemble global qu'est la cellule avec des modèles généraux qui sont à la base de la métabolomique. Ces domaines en "omique" intègrent la bioinformatique et les bases de données. |
| Aperçu des différents niveaux (non hiérachiques) de la régulation cellulaire qui aboutissent à une régulation du métabolisme | ||
| Signaux extracellulaires | Perception d'une grande diversité de signaux | Récepteurs Communication entre cellules |
| Signaux intracellulaires | Transmission des signaux | Cascade de signalisation - Messagers secondaires |
| Gènes | Régulation transcriptionnelle | Taux réel de gènes transcrits : facteurs de transcription, ARN non codant, régulation épigénétique |
| Transcrits | Régulation post-transcriptionnelle et traductionnelle | Taux réel de transcrits traduits : polyadényltion alternative, interférence ARN |
| Protéines | Modifications co- et post-traductionnelles | Activation des précurseurs : zymogènes Phosphorylation - glycosylation, ... |
| Translocation dans les compartiments cellulaires | Peptide signal (adressage) - canaux de translocation | |
| Protéolyse | Dégradation non spécialisée (lysozome) Dégradation spécialisée (ubiquitine - protéasome / SUMOylation) |
|
| Enzymes | Modulation de l'activité des enzymes | Allostérie - pH - force ionique - inhibitions - ... |
| Métabolites | Transports | Passifs ou actifs (diffusion, pompes, canaux ioniques, ...) |
| Concentration réelle des métabolites disponibles | Compartimentation sub-cellulaire |
|
| Régulation énergétique | Variation d'énergie libre de Gibbs | |
2. Les signaux extracellulaires et intracellulaires Les signaux jouent un rôle capital dans la régulation du métabolisme. Ils émanent :
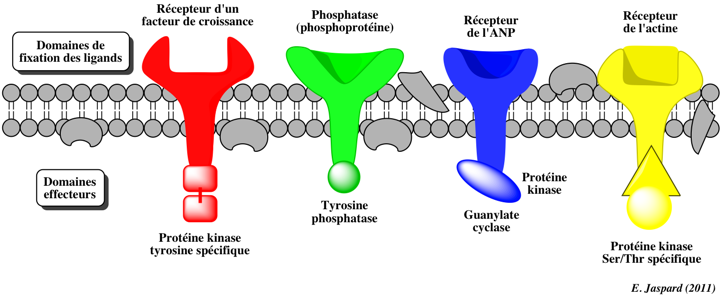
Les signaux extracellulaires et intracellulaires sont captés par des récepteurs de différents types :
Messagers secondaires Ce sont des molécules qui relayent le signal jusqu'à la cible intracellulaire via un ensemble d'évènements que l'on appelle la transduction du signal ou cascade de signalisation. Parmi les messagers secondaires, on peut citer : l'AMP cyclique, le calcium, la calmoduline, l'inositol triphosphate, le diacylglycérol et un très grand nombre d'autres molécules ... Dans la grande majorité des cas, cette cascade d'évènements de signalisation :
|
3. La fonction des protéines et l'activité des enzymes
a. Les modes de régulation de l'activité des enzymes
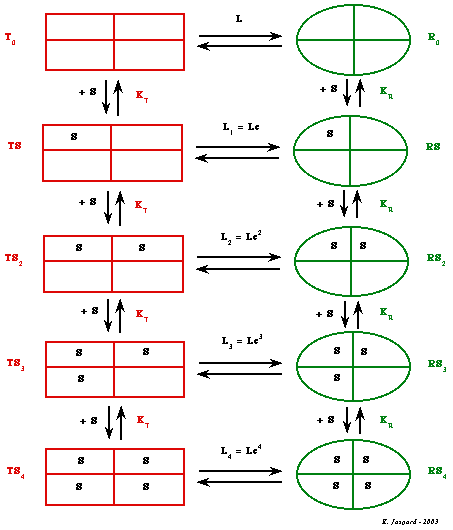
Les effecteurs allostériques modifient la structure des enzymes donc ils modulent très finement leur activité. Figure ci-dessous : modèle de Monod, Wyman & Changeux (phosphofructokinase, enzyme tétramérique constituée de 4 sous-unités identiques).
Les deux états conformationnels que peut adopter un protomère sont dénommés respectivement :
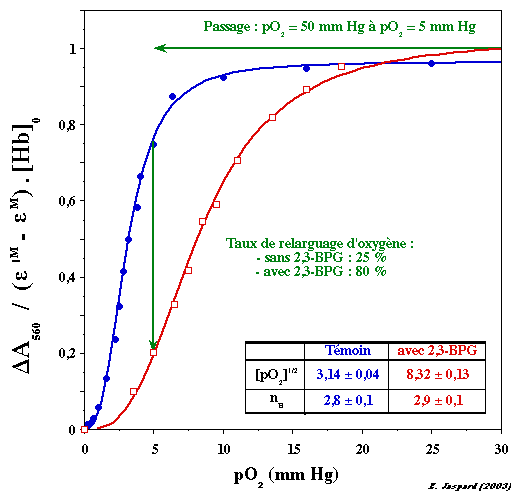
Ces deux conformations sont en équilibre (quelle que soit la concentration d'un ligand donné) et cet équilibre est régit par la constante allostérique L0. Figure ci-dessous : courbes de saturation de l'hémoglobine par l'oxygène en présence ou non du 2,3-bisphosphoglycérate (2,3-BPG).
L'hémoglobine fixe moins bien l'oxygène quand le 2,3-BPG est aussi fixé : quand la pression en oxygène diminue (passage poumons => muscles), l'oxygène est donc relargué de l'hémoglobine. |
b. La disponibilité des métabolites substrats L'activité des enzymes dépend aussi de l'accessibilité et de la disponibilité de leurs métabolites substrats. Dans le cas de substrats venant de l'extérieur de la cellule, il faut qu'ils puissent y pénétrer. Ou bien qu'ils puissent pénétrer dans un compartiment sub-cellulaire (et en sortir) comme la mitochondrie ou le chloroplaste par exemple, sièges des réactions qui synthétisent notamment l'ATP. La composition des membranes est donc capitale pour assurer cette pérméabilité aux métabolites dans les 2 sens. Il existe de nombreux mécanismes de transport des métabolites :
Un exemple important est le transport du glucose : son entrée/sortie (transporteur GLUT4) dans la cellule est contrôlé en partie par l'insuline. Selon le taux de cette hormone, des quantités variables de glucose entrent dans la cellule, modulant ainsi le niveau d'activité des enzymes impliqués dans le métabolisme énergétique (entre autre). |
|
4. La concentration des métabolites La concentration des métabolites d'une réaction donnée, disponibles à un moment donné est un mode de régulation du métabolisme. Cette concentration est régie par la variation d'énergie libre de Gibbs associée à chaque réaction du métabolisme. Pour une réaction quelconque : A + B <=> C + D La variation d'énergie libre de Gibbs de cette réaction s'écrit : [C]éq
. [D]éq
[C]φ
. [D]φ
[C]éq
. [D]éq De ces relations, il ressort un concept primordial : c'est la concentration physiologique des métabolites qui régit la variation d'énergie libre d'une réaction, donc sa spontanéité, donc le sens dans lequel elle se déroule au sein d'une voie métabolique. |
|
ΔG'réaction = 0 réaction réversible |
Les réactions biochimiques pour lesquelles le rapport des concentrations physiologiques des métabolites est proche du rapport des concentrations à l'équilibre, sont dites se dérouler au voisinage de l'équilibre. Ces réactions sont donc facilement réversibles (elles se déroulent dans le sens du flux métabolique ou dans le sens inverse : exemple de la glycolyse et de la néoglucogénèse). Pour ces réactions, la moindre tendance à s'éloigner d'un état proche de l'équilibre est immédiatement rétablie par le très haut pouvoir de catalyse des enzymes qui les contrôlent. |
ΔG'réaction < 0 réaction irréversible |
Les réactions pour lesquelles ce n'est pas le cas sont des réactions irréversibles qui se caractérisent par des valeurs de ΔG' très négatives. Les réactions irréversibles régulent souvent le flux global des voies métaboliques. Elles sont catalysées par des enzymes qui sont en quantités limitées dans la cellule. La biosynthèse de ces enzymes est soumise à des mécanismes de régulation extrêmement fins (notamment au niveau de la transcription des gènes qui codent ces enzymes). Ces enzymes ont des propriétés catalytiques particulières et sont le plus souvent des enzymes à régulation dite allostérique (par opposition aux enzymes dites Michaéliennes). Les métabolites de ces réactions régulent souvent l'activité d'enzymes qui catalysent d'autres réactions en aval ou en amont. Ces métabolites sont appelés effecteurs de l'activité enzymatique. Cette dernière caractéristique explique aussi pourquoi les réactions réversibles ne peuvent pas constituer des points de contrôle du métabolisme : l'activité des enzymes qui catalysent les réactions réversibles n'est, dans la grande majorité des cas, pas modulable par des effecteurs. |
ΔG'réaction > 0 réaction impossible isolément |
Réaction impossible isolément dans le sens du flux métabolique. Elle nécessite un apport d'énergie fournie par une réaction qui lui est couplée. Dans les réactions des voies métaboliques, le couplage dépend de la présence d'un intermédiaire commun aux divers composants de la réaction globale. Cet intermédiaire commun est une molécule dont la structure chimique lui confère une forte énergie libre de Gibbs qui peut être transférée à une autre molécule. La molécule universelle qui possède cette forte énergie libre est l'adénosine triphosphate ou ATP. |
5. La quantité d'enzymes synthétisées Puisque les enzymes catalysent les réactions biochimiques :
La quantité d'enzymes synthétisées est contrôlée de diverses manières. a. Plusieurs gènes (famille multigènique) codant des enzymes clé de la régulation du métabolisme En cas de mutation rendant non fonctionnel un gène, cette redondance de gènes permet à la cellule d'avoir d'autres gènes fonctionnels et 'ainsi le taux d'enzyme clé synthétisée est constant. Par exemple, il existe au moins 7 gènes codant la phosphofructokinase chez Arabidopsis thaliana (voir Mustropha et al. (2007) "Characterisation of the ATP-dependent phosphofructokinase gene family from Arabidopsis thaliana" Febs Letter 581; 2401-2410). Voir un exemple d'application bioinformatique à l'analyse de la famille multigènique de la globine. b. Le niveau de transcription des gènes Exemple de la répression catabolique de l'opéron lactose chez les bactéries : quand la concentration en glucose diminue, le métabolisme du lactose devient nécessaire. Le signal de carence alimentaire est une augmentation de la concentration de l'AMP cyclique (AMPc). L'AMPc se fixe à la protéine "CAP" ("Catabolite gene Activator Protein"). Ce complexe se lie à l'ADN et augmente l'affinité de l'ARN polymérase pour le promoteur de l'opéron, ce qui se traduit par une augmentation d'un facteur 50 de la transcription de l'opéron lactose. |
|
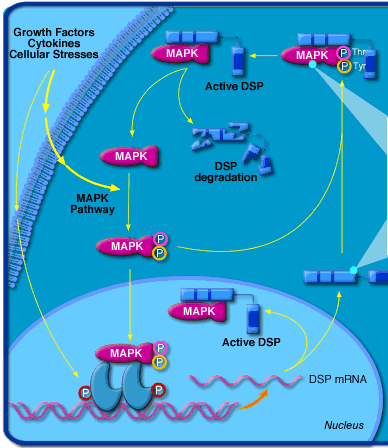
c. La modulation de l'activité et de la localisation des facteurs de transcription La phosphorylation et la déphosphorylation de certains facteurs de transcription modulent leur activité. Exemple : les MAP kinases ("Mitogen-activated protein kinases") sont impliquées dans des voies de signalisation en réponse à des signaux extracellulaires (facteurs de croissance, cytokines, ultraviolets, agents de stress, ... - figure ci-contre).
Source : Biocarta Les MAP kinases régulent un grand nombre d'activités cellulaires, en particulier la transcription des gènes. Ce sont des sérine/threonine protéines kinases qui phosphorylent certains facteurs de transcription, modifiant ainsi leur activité et parfois leur localisation sub-cellulaire (passage du cytoplasme vers le noyau). DSP : "dual-specificity phosphatase". Les phosphatases hydrolysent les groupements phosphoryle (réaction inverse des kinases). |
|
d. La régulation post-transcriptionnelle et la régulation traductionnelle
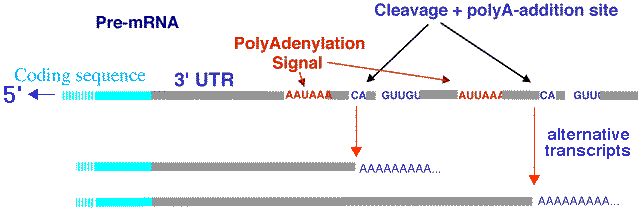
Une étude des transcrits issus de 10 chromosomes humains a montré que 19% sont poly(A)+, 44% sont poly(A)-, c'est-à-dire non polyadénylés et 37% sont poly(A)+ et poly(A)-. Voir : Cheng et al. (2005) "Transcriptional maps of 10 human chromosomes at 5-nucleotide resolution" Science 308, 1149-1154 La polyadénylation alternative génère différents transcrits à partir d'un même gène (schéma ci-dessous).
Source : D. Gautheret - INSERM ERM206 Polyadénylation chez les procaryotes : voir Sarkar (1997) "Polyadenylation of mRNA in prokaryotes" Annual Rev. Biochem. 66, 173-197 |
|
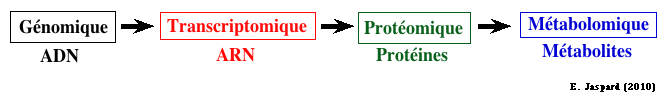
6. Les moyens d'étude de la régulation du métabolisme Les moyens sont les méthodes biochimiques, génétiques et de biologie cellulaires classiques. D'autres approches plus récentes y contribuent aussi.
a. La génomique fonctionnelle et la génomique structurale qui ont pour but entre autres :
b. La transcriptomique qui analyse l'ensemble des transcrits (produits de la transcription des gènes) dans une cellule ou un compartiment cellulaire à un moment donné. Elle utilise des techniques telles que :
c. La protéomique qui a pour but d'identifier et de quantifier l'ensemble des protéines synthétisées ou protéome, à un moment donné et dans des conditions données au sein d'un tissu, d'une cellule ou d'un compartiment cellulaire. La protéomique et la transcriptomique sont donc des approches complémentaires trés puissantes qui peuvent être utilisées pour des études fondamentales ou appliquées en biologie, en médecine, en agriculture. En effet, dans les deux cas, les infomations recueillies permettent d'aborder l'ensemble des réponses cellulaires dans leur globalité et non plus de manière partielle. La protéomique apporte des réponses auxquelles la transcriptomique ne peut répondre :
d. La métabolomique De plus en plus de génomes d'organismes sont séquencés ou en cours de séquençage. Voir les bases de données suivantes :
Les modèles du métabolisme à l'échelle d'un génome établissent le pont entre les informations issues du séquençage de ce génome (et des analyses subséquentes de ce génome), les données biochimiques et les phénotypes lié à ce métabolisme. Cette démarche de métabolomique est récente car elle est la conséquence logique de tous les domaines en "omique". e. Les outils bioinformatiques et les bases de données qui ont été développées spécifiquement pour l'étude de la régulation du métabolisme. |
| 7. Liens Internet et références bibliographiques |
| "The
UCSD-Nature Signaling Gateway" - A comprehensive resource for information
about cell signaling.
KEGG PATHWAY Database "MetaCyc" : database of nonredundant, experimentally elucidated metabolic pathways (more than 1,200 pathways from more than 1,600 different organisms). REACTOME : "Open-source, open access, manually curated and peer-reviewed pathway database" |
![]()