| Analyse des sites de coupure des peptides signaux (bactéries et réticulum endoplasmique) |
| Tweet |
|
|
Préambule et différents types de séquences de signaux de ciblage 1. Descriptions des données 2. Analyse de séquences de peptides signaux et de séquences complètes 3. Prédiction de peptides signaux 4. Calculs de paramètres physico-chimiques |
5. Les outils de conversion de formats 6. Compléments sur le traitement protéolytique des peptides signaux 7. Les systèmes protéolytiques peptidases des mitochondries et des chloroplastes 8. Peptide signal et particule de reconnaissance du peptide signal ("Signal Recognition Particle") 9. Liens Internet et références bibliographiques |
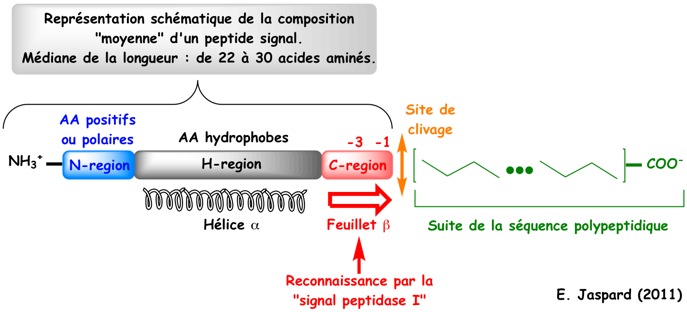
a. Préambule Certaines chaînes polypeptidiques commencent par une séquence appelée peptide signal (ou séquence d'adressage) qui indique à la cellule le compartiment vers lequel adresser ces chaînes polypeptidiques ou bien si elles doivent être sécrétées de la cellule. Günter Blobel a reçu le Prix Nobel en 1999 pour sa découverte que les chaînes polypeptidiques possèdent des signaux intrinsèques qui régissent leur transport et leur localisation dans la cellule. Un peptide signal ou séquence signal (ou signal de localisation ou peptide transit - voir le glossaire) est une séquence située à l'extrémité N-terminale (voire C-terminale / exemple : le tripeptide PTS1 - "Peroxisome targeting signal 1") de la majorité des protéines destinées :
La voie de sécrétion générale Sec ("general secretory pathway") dirige la translocation des protéines :
Les bactéries, les archées, les chloroplastes et certaines mitochondries possèdent également une voie appelée Tat ("Twin-arginine translocation") qui reconnaît généralement des peptides signaux plus longs et moins hydrophobes avec 2 résidus arginines consécutifs dans la région N-terminale. Contrairement à la voie Sec qui transporte les protéines non repliées, la voie Tat peut transloquer les protéines repliées à travers la bicouche lipidique membranaire. Pendant ou après la translocation membranaire, une signal peptidase (SPase) spécifique hydrolyse le peptide signal.
|
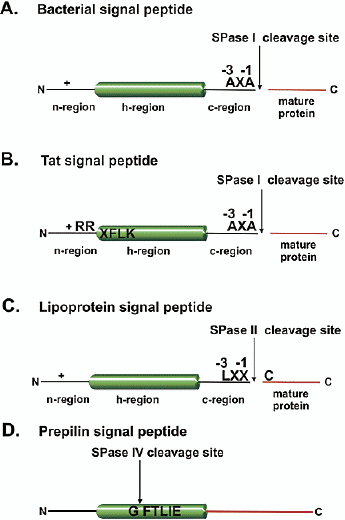
b. Différents types de séquences de signaux de ciblage (A) Le peptide signal bactérien (figure ci-dessous) adresse la protéine à la voie Sec ("general protein secretion"). (B) Le peptide signal Tat (avec le motif consensus Arg-Arg) adresse la protéine à la voie Tat ("twin-arginine translocation" - plantes, bactéries et archées). Chez les plantes, cette voie se trouve dans la membrane thylakoïde du chloroplaste. Voir les serveurs Web pour la prédisction : Tatfind, PRED-TAT. (C) Le peptide signal lipoprotéine contient la séquence "lipobox" adresse la protéine à la voie Sec. (D) Le peptide signal "prepilin-like" existe chez les piliprotéines de type IV de certaines bactéries Gram - et chez certaines protéines "prépiline" impliquées dans la sécrétion de type II.
Source : Paetzel et al. (2002) Voir un développement sur les systèmes des mitochondries et des chloroplastes. Les peptidases qui hydrolysent les peptides signaux des protéines sécrétées par les bactéries ou qui sont adressées au réticulum endoplasmique sont des protéases à sérine. Les systèmes peptidases des mitochondries et des chloroplastes sont d'origine bactérienne et les gènes codant ces peptidases ont été conservés au cours de l'évolution. Dans les mitochondries et les chloroplastes de Arabidopsis thaliana, 26 peptidases impliqués dans la dégradation des protéines ont été identifiées. Elles appartiennent à 4 familles :
|
|
1. Descriptions des données à analyser
a. Analyse des longueurs des peptides signaux
Valeurs médianes des longueurs des 3 régions des peptides signaux (Tuteja, 2005) :
|
| Organisme | Séquences "curées" SPdb | Longueur SPs | Médiane longueur SPs |
| SPsEuk | 1877 |
Plus longue chez les bactéries Gram+ car "H-region" plus longue et distribution bi-modale avec 2 pics (29 AA et 41 AA). |
22 |
| SPsGram+ | 168 | 30 | |
| SPsGram- | 307 | 23 | |
| SPs : peptide signal ("signal peptide") - SPsEuk : peptide signal des eucaryotes | |||
|
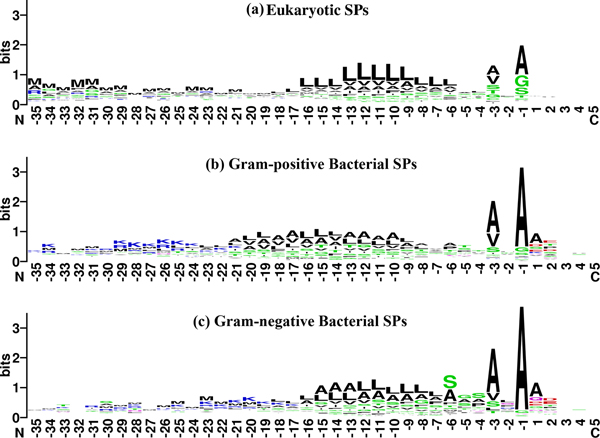
b. Analyse des fréquences des acides aminés autour du site de clivage des peptides signaux Voir un cours sur les sous-sites de fixation des hydrolases / protéases - peptidases. La position -1 (ou P1) est l'acide aminé en position C-terminale du peptide signal et la position +1 (ou P1') est l'acide aminé en position N-terminale de la chaîne polypeptidique mature.
Source : Choo & Ranganathan (2008)
Voir l'aspect mathématique de la notion de logo. |
Fréquence des acides aminés aux positions P3 - P2 - P1 / P'1 - P'2 |
||||
| P3 | P2 | P1 | P'1 | P'2 |
| Ala (Gly > Ser > Thr) | Leu > Ser | Ala (Gly > Ser > Thr) | Pas d'acide aminé préférentiel | |
| Ala > Val | Ser > Gln > Phe > Ala | Ala | Ala (Asp > Ser > Glu) | Thr > Glu > Pro > Ser |
| Ala > Val | Leu > Gln > Phe > His | Ala | Ala (Gln > Asp > Glu) | Asp > Glu > Pro > Thr |
Pro absent de P3 à P'1. |
||||
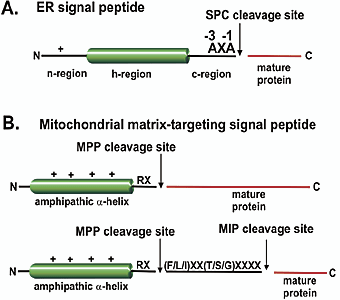
Règle dite (-3, -1) du site de clivage par les "Type I Signal Peptidases" (SPases) : les acides aminés en position -3 et -1 par rapport au site de clivage sont petits et neutres (Tuteja, 2005). La spécificité du site de clivage des SPases I ("Type I Signal Peptidases") est souvent désignée "Ala-X-Ala" car Ala est l'acide aminé le plus fréquent en position -3 et -1. Chez les Eucaryotes, cette règle est plus "souple" :
|
| 2. Analyse de séquences de peptides signaux et de séquences complètes |
|
Récupérer les séquences des peptides signaux et des séquences complètes ("Full-length sequence") de Arabidopsis thaliana à partir de SPdb - option "Download".
|
|
Analyser les données contenues dans le fichier "P30224" de Uniprot. |
AFP1_ARATH |
| Aller à l'item "PTM / Processing". Cliquer sur les chiffres ("1-29") ou sur la barre orange ("Graphical view"). Effectuer un BLAST contre la base de données "Plants" (sélectionnée dans le menu "Target database"). |
|
| Revenir à la page "P30224" - Uniprot. Quel est le fichier correspondant au NCBI ("GenBank / GenPept") ? Récupérer la séquence FASTA de ce fichier au NCBI. |
AAD02502 |
| Comparer les 10 meilleurs "hits" renvoyés avec ceux obtenus en effectuant un BLAST avec la même séquence au NCBI. Attention aux choix de la matrice et des valeurs de "Threshold" et de "gaps" dans l'un et l'autre des deux environnements. |
|
|
3. Prédiction de peptides signaux Comparer la séquence du peptide signal de AFP1_ARATH obtenue à partir de SPdb avec la séquence du peptide signal prédite par :
Trouver quelques protéines de Medicago truncatula qui possèdent un peptide signal. Piste 1 : quel résultat donne une recherche "Entrez" avec les mots clés "medicago truncatula peptide signal" ?
|
|
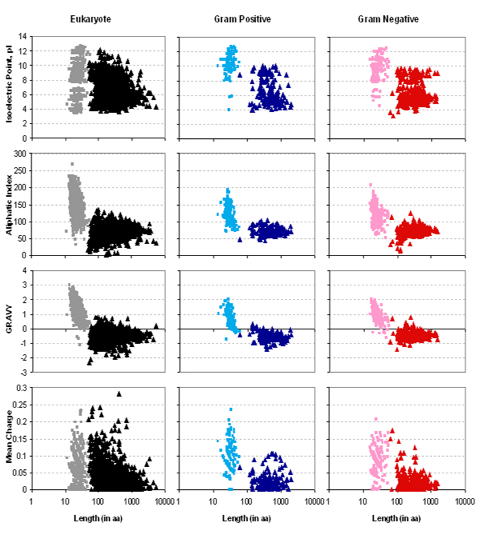
4. Calculs de paramètres physico-chimiques Aller à Expasy. Dans le menu de gauche, ouvrir le sous-menu "proteomics". Dans la liste "Tools" (à droite), choisir "ProtParam". Calculer les valeurs de pI, "Aliphatic index" et GRAVY ("GRand AVerage of hydropathY") à partir des séquences des peptides signaux : 2SS1_ARATH, 2SS2_ARATH, 2SS3_ARATH, SCP8_ARATH. A partir de "Protein calculator", calculer les valeurs de la charge nette à pH 7. A quelles propriétés physico-chimiques des acides aminés correspondent les paramètres "Aliphatic index" et GRAVY ? Interpréter les valeurs obtenues. |
| Séquence peptide signal | pI | Aliphatic | GRAVY | Total AA chargés / total AA | Charge nette à pH 7 |
| 2SS1_ARATH | 7,82 | 149.05 | 1,757 | 1 / 21 | 0,8 |
| 2SS2_ARATH | 7,82 | 125,71 | 1,590 | 1 / 21 | 0,8 |
| 2SS3_ARATH | 7,82 | 144,29 | 1,638 | 1 / 21 | 0,8 |
| SCP8_ARATH | 8,29 | 174,21 | 1,042 | 3 / 19 | 1,4 |
Récupérer les séquences matures à partir des séquences complètes correspondantes.
Source : Choo & Ranganathan (2008) |
|
Les séquences nécessaires à ces excercices sont regroupées dans le fichier : TouteSequences.doc.
|
6. Compléments sur le traitement protéolytique des peptides signaux a. Définitions La pré-séquence ("presequence") : c'est un peptide N-terminal d'une protéine précurseur qui dirige la protéine vers les mitochondries et est protéolysé dans la matrice mitochondriale. La pré-séquence peut être constituée d'une partie N-terminale qui dirige la protéine vers les mitochondries, suivie d'un peptide supplémentaire clivable qui dirige la protéine vers un compartiment intra-mitochondrial spécifique (par exemple, l'espace intermembranaire ou la membrane interne). Le peptide de ciblage ("targeting peptide") : c'est un peptide N-terminal clivable d'une protéine précurseur qui dirige la protéine vers les mitochondries ou les chloroplastes. Le peptide de ciblage est un terme plus générique pour décrire des peptides de reconnaissance d'organelles et ce terme inclue la notion de pré-sequence et de peptide transit (voir ci-dessous). Le signal de ciblage ("targeting signal") : c'est un terme générique qui décrit une séquence qui dirige une protéine vers un organite (exemples : mitochondries ou chloroplastes) ou vers un compartiment intra-organelle. Les signaux de ciblage sont ou ne sont pas protéolysés. Contrairement aux protéines mitochondriales ou chloroplastiques qui possèdent un peptide de ciblage clivable, certaines protéines d'organites contiennent des signaux de ciblage non clivables. Exemples :
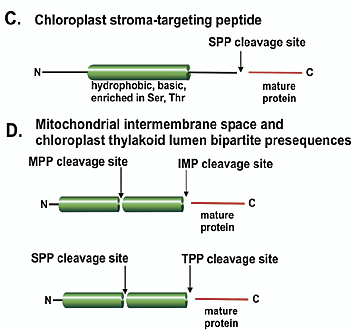
Le peptide transit ("transit peptide") : c'est un peptide N-terminal d'une protéine précurseur qui dirige la protéine vers les chloroplastes et est protéolysé dans le stroma des chloroplastes. Le peptide transit peut être constitué d'une partie N-terminale qui dirige la protéine vers les chloroplastes suivie d'un peptide supplémentaire clivable qui dirige la protéine vers un compartiment intra-chloroplastique spécifique (par exemple, la membrane thylakoïde ou le lumen). Voir un cours sur la biogénèse des protéines membranaires intégrales. b. Caractéristiques des peptides signaux Les pré-séquences mitochondriales se distinguent des peptides transit chloroplastiques par leur longueur moyenne, leur composition en acides aminés et leur propension à former des structures secondaires. Basée sur l'analyse des extrémités N-terminales de protéines matures, on estime que la longueur des pré-séquences mitochondriales varie :
La longueur des peptides transit chloroplastiques varie de 13 - 26 à 146 acides aminés. Ils sont généralement plus longs que les pré-séquence mitochondriales avec une moyenne de 58 acides aminés. Bien qu'il y ait des similitudes de composition en acides aminés des peptides de ciblage (les résidus les plus abondants dans les deux types de peptides de ciblage sont Ser, Leu, Ala et Arg), il y a des différences subtiles dans leur partie N-terminale :
Les pré-séquences ont une propension à former des hélices alpha amphiphiles et les peptides transit sont en général non structurés. Ces derniers peuvent cependant adopter une conformation alpha-hélicoïdale dans des environnements qui miment la membrane. c. Importation dans les mitochondries et les chloroplastes Plus de 95 % des protéines mitochondriales et chloroplastiques sont synthétisées dans le cytosol puis importées dans ces organites : soit l'importation d'environ 1400 protéines dans les mitochondries et de 1500 à 2300 protéines dans les chloroplastes. Pour garantir un ciblage et un import corrects, 70% et 85%, respectivement, des protéines mitochondriales et des protéines chloroplastiques sont synthétisées dans le cytosol sous forme de pré-protéines contenant des signaux de ciblage clivables. Ces signaux sont le peptide N-terminal des protéines importées qui est reconnu par des récepteurs localisés au niveau des membranes externes des organites (voir ci-dessous "Particule de reconnaissance du peptide signal"). En raison de leur toxicité et de leur influence potentielle sur le métabolisme de la cellule, le recyclage des peptides générés dans les mitochondries et les chloroplastes doit être régulé. Dans les mitochondries et dans les chloroplastes, les peptides de ciblage qui ont été clivés sont eux-mêmes protéolysés par la protéase "Presequence Protease" (PreP). |
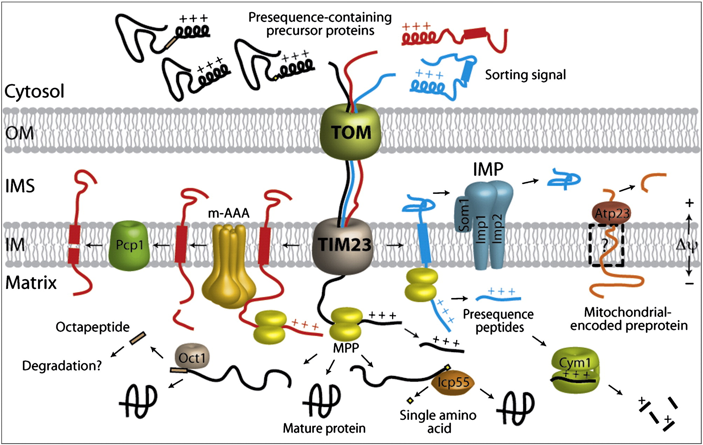
7. Les systèmes protéolytiques peptidases des mitochondries et des chloroplastes a. Les systèmes des mitochondries La plupart des protéines précurseurs dirigées vers la matrice mitochondriale sont protéolysées dans la matrice de cet organite par différentes peptidases :
Source : Mossmann et al. (2012) |
Des peptidases supplémentaires de la membrane interne ("Inner Membrane Proteases", Atp23, rhomboïdes et protéases m-AAA) catalysent des traitements supplémentaires qui débouchent sur :
Source : Paetzel et al. (2002) b. Les systèmes des chloroplastes Les peptides de ciblage chloroplastiques sont protéolysés dans le stroma par la peptidase "Stromal Processing Peptidase" (SPP). Chez Arabidopsis thaliana, le système SPP clive plus de 2000 protéines substrats qui ont en commun un motif faiblement conservé avec un résidu Ala en position 1 et un résidu Val en position 3. Si la protéine est transloquée dans la lumière des thylakoïdes, une séquence supplémentaire de transfert vers les thylakoïdes est clivée par la peptidase "Thylakoidal Processing Peptidase" (TPP). Le fonctionnement correct de la protéine D1 du centre réactionnel du photosystème II nécessite la maturation de son extrémité C-terminale par la protéase CtpA ("Carboxy-terminal processing protease A"). |
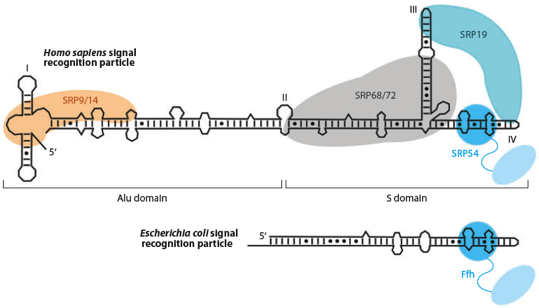
8. Peptide signal et particule de reconnaissance du peptide signal ("Signal Recognition Particle") Chez les Procaryotes et les Eucaryotes, un complexe appelé "particule de reconnaissance du peptide signal" ("signal recognition particle" - SRP) se fixe au peptide signal qui émerge du ribosome. Le complexe SRP, le ribosome et la RNAse P sont les 3 particules ribonucléoprotéiques conservées dans tous les règnes du vivant. Chez les Eucaryotes, le complexe SRP peut être structuralement et fonctionnellement divisé en deux domaines :
En bas, modèle chez les Procaryotes : interaction avec la protéine Ffh.
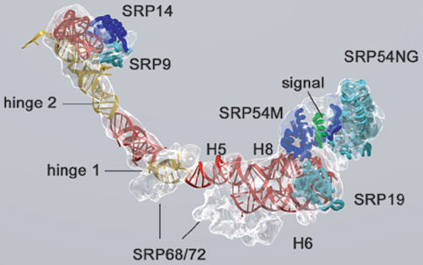
Source : Akopian et al. (2013) Figure ci-dessous : structure de l'ARN 7S, des protéines SRP et du peptide signal.
Source : Halic & Beckmann (2005) Dans les chloroplastes des plantes supérieures SRP a connu de profonds changements évolutionaires. A la place d'un ARN, il semble utiliser une protéine spécifique au chloroplaste : cpSRP43. Cependant, des homologues de SRP54 et du récepteur SRP sont présents dans les chloroplastes. |
|
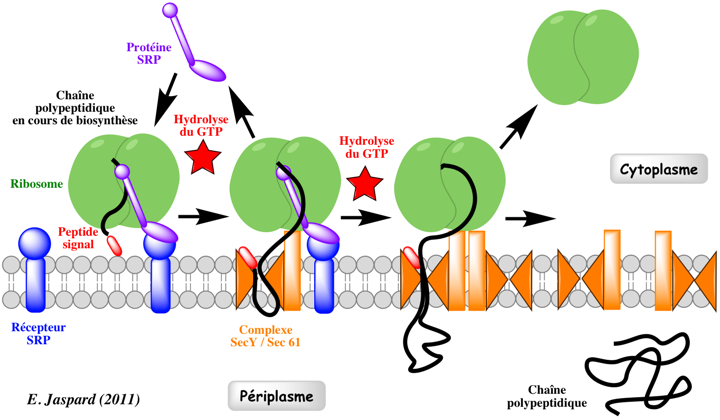
Mécanisme Le complexe SRP ralentit l'élongation de la chaîne polypeptidique en cours de biosynthèse ("elongation arrest").
Le complexe SRP se fixe au récepteur du SRP : il adresse ainsi les protéines à la membrane plasmique chez les Procaryotes ou à la membrane du RE chez les Eucaryotes. Le complexe [ribosome - chaîne polypeptidique en cours de biosynthèse] est délivré à la membrane cible via l'interaction du complexe SRP avec le récepteur de SRP. Ce complexe est transféré au translocon Sec61p chez les Eucaryotes ou au translocon secYEG chez les bactéries et les archées (voir ci-dessous). La chaîne polypeptidique en cours d'élongation emprunte le canal pour passer entièrement dans la lumière du RE (dans le cas d'une protéine sécrétée, dirigée vers l'appareil de Golgi) ou pour être intégrée à la membrane (dans le cas d'une protéine membranaire). Le complexe SRP et le récepteur de SRP se dissocient pour participer à la translocation d'une autre chaîne polypeptidique en cours de biosynthèse. Il s'agit donc d'un transport co-traductionnel puisque l'élongation de la chaîne polypeptidique continue pendant le passage dans le RE. Le cycle SRP est un processus dont l'énergie est fournie par l'hydrolyse du GTP : le complexe SRP et le récepteur du SRP possèdent une acticité GTPase. Voir une belle vidéo du mécanisme : "Signal-Recognition-Particle". |
| 9. Liens Internet et références bibliographiques |
![]()