
| L'endosymbiose |
| Tweet |
|
|
1. Evolution biologique des procaryotes aux eucaryotes : l'endosymbiose 2. Endosymbiose primaire et endosymbiose secondaire 3. Arguments supplémentaires pour une origine bactérienne du chloroplaste |
4. Arguments supplémentaires pour une origine bactérienne de la mitochondrie 5. Evolution des organismes via la phylogènie du cytochrome C 6. Liens Internet et références bibliographiques |
| 1. Evolution biologique des procaryotes aux eucaryotes : l'endosymbiose |
| a. Deux grands types d'organismes | |
|
Les procaryotes (du grec, pro = avant et karyon = noyau) sont des êtres unicellulaires, dépourvus de noyau et bordés d'une membrane. |
Les cellules des eucaryotes (eu, propre) sont généralement de plus grande taille, avec un noyau bordé d'une double membrane (ou enveloppe nucléaire). Le plus souvent, elles contiennent aussi des membranes internes qui cloisonnent la cellule en y délimitant des organites qui ont des fonctions biologiques spécialisées. |
|
|
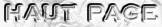
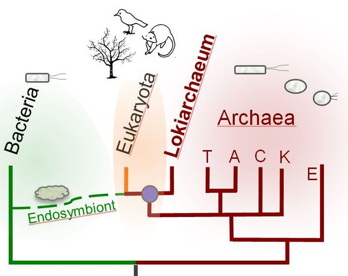
b. Proximité archées - procaryotes - eucaryotes En 2015, le génome d'une lignée d'archée, appelée Lokiarchaea (phylum Lokiarchaeota), obtenue à partir d'un métagénome prélevé dans des sédiments marins profonds de l'Atlantique a été séquencé puis assemblé par bioinformatique. Le génome contient un total de 5381 gènes codant des protéines dont un grand nombre code des protéines caractéristiques des eucaryotes (appelées protéines de signature eucaryotes - "Eukaryotic Signature Proteins"). De plus, la reconstruction de l'arbre phylogénétique du vivant incluant cet organisme a démontré que le nouveau phylum archéen (appelé Lokiarchaeota) forme un groupe monophylétique avec les eucaryotes : il serait le parent le plus proche des eucaryotes et un intermédiaire entre archées et eucaryotes (Spang et al., 2015).
Source : Astrobiology - NASA
Source : Embley & Williams (2015) Le nom Lokiarchaea est lié au site de prélèvement du métagénome : la dorsale médio-océanique de l'Arctique à une profondeur de 3200 m, située à 15 km du site d'évent actif appelé "Loki's Castle" (le "chateau de Loki", divinité de la mythologie nordique). |
|
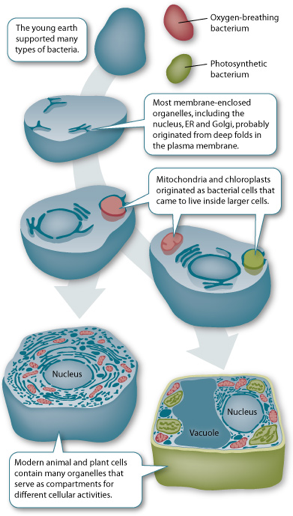
c. Notions élémentaires de la théorie de l'endosymbiose Selon les éléments à la base de la théorie endosymbiotique [développée principalement par Max FJR Taylor (1979) et Lynn Margulis (1993)], les cellules eucaryotes proviennent de l'association de plusieurs procaryotes. L'endosymbiose reprend des idées énoncées par le botaniste russe Konstantin Mereschkowski en 1905 (symbiogénèse). "Serial Endosymbiosis Theory" - SET :
L'endosymbiose de l'origine de la cellule eucaryote postule que :
Source : Futura-Sciences L'endosymbiose est étayée par les faits suivants (liste non exhaustive) :
Source : "The evolution of cell" 1. La taille des mitochondries et des chloroplastes est semblable à celle des bactéries (≈ 1 μm). 2. Chacun de ces organites possède une double membrane. La membrane externe dériverait de la cellule originale hôte. 3. La membrane externe des mitochondries et des chloroplastes et la membrane des bactéries contiennent des porines (protéines de transport). La cardiolipine est un lipide membranaire trouvé exclusivement dans la membrane interne des mitochondries et la membrane des bactéries. 4. Chacun de ces organites possède un ADN qui lui est propre (organites semi-autonomes). La comparaison des génomes indique que les cyanobactéries ont contribué à l'origine génétique des plastes. 5. Cet ADN est circulaire. 6. Chacun de ces organites possède le matériel nécessaire pour la synthèse protéique (ARNt, ribosomes, polymérases). La N-formylméthionine est utilisée comme acide aminé d'initiation de la traduction pour les protéines de ces organites comme pour les bactéries. 7. Chacun de ces organites peut se diviser par étranglement médian (après avoir dupliqué le matériel génétique). Ils sont en mouvement constant, ils fusionnent et forment un réseau dynamique. |
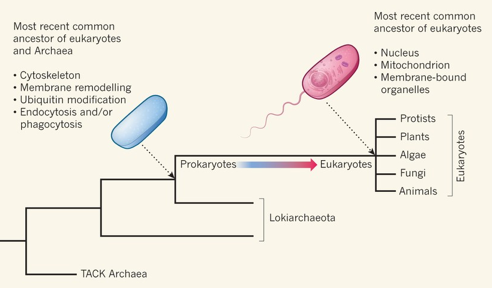
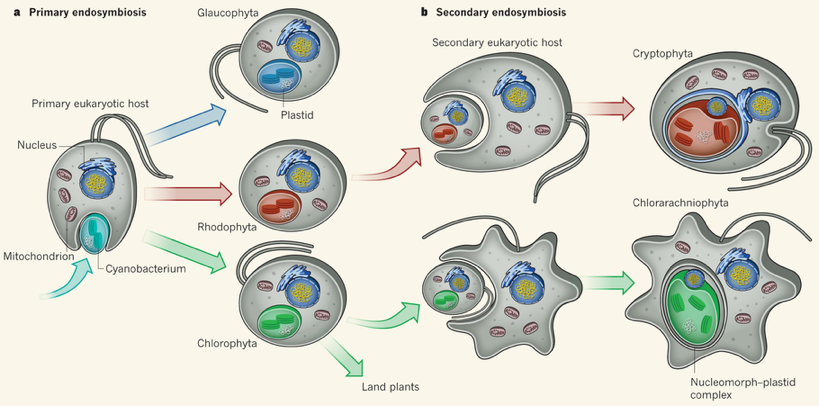
2. Endosymbiose primaire et endosymbiose secondaire a. L'endosymbiose primaire 1. Il y a environ 1,5 - 2 milliards d'années, une cellule eucaryote primitive absorbe une α-protéobactérie* : la protéobactérie devient une mitochondrie qui assure une respiration "primitive" au sulfate. Il en résulte une cellule eucaryote hétérotrophe. *α-protéobactérie : une bactérie Gram négatif (contenant une membrane externe), comme les espèces Rickettsia, qui est probablement la bactérie vivante la plus proche des mitochondries. Il existe de nombreux modèles, parmis lesquels :
2. Cette cellule eucaryote hétérotrophe absorbe à son tour une cyanobactérie, organisme photosynthétique contenant un plaste : la cyanobactérie devient un chloroplaste. Il en résulte une cellule eucaryote autotrophe. La membrane interne du chloroplaste est d'origine cyanobactérienne et la membrane externe a pour origine la membrane plasmique de la cellule qui l'a absorbé.
|
b. L'endosymbiose secondaire Une cellule eucaryote hétérotrophe absorbe une cellule eucaryote autotrophe contenant des chloroplastes : ceux-ci sont alors entourés de quatre membranes (2 membranes du chloroplaste + 1 membrane plasmique + 1 membrane de phagocytose). c. L'endosymbiose secondaire : cas de certaines algues Une cellule eucaryote hétérotrophe absorbe une cellule de Rhodophyta (algues rouges) ou une cellule de Chlorophyta (algues vertes). Cette endosymbiose secondaire débouche sur 2 nouveaux groupes d'algues : les Cryptophytes et les Chlorarachniophytes. Ces organismes contiennent des mitochondries et des plastes mais aussi un organite appelé le nucléomorphe, réminiscence du noyau de l'algue. Exemple : Guillardia theta.
Source : Gould (2012) La comparaison des génomes, notamment des algues, est un point capital pour conforter la théorie endosymbiotique. |
3. Arguments supplémentaires pour une origine bactérienne du chloroplaste Une bactérie photosynthétique telle qu'une Cyanobactérie et un chloroplaste d'une cellule eucaryote ont des points communs :
Caractéristiques des génomes chloroplastiques Les chloroplastes possèdent plusieurs copies d'un génome circulaire mesurant de 120 à 217 kb (150 kb en moyenne) chez les plantes supérieures. Le génome des chloroplastes contient en moyenne une centaine de gènes. Voir la carte interactive du génome de chloroplaste. Le séquençage complet de l'ADN chloroplastique de plusieurs plantes très éloignées au sens évolutif (algues, bryophytes, gymnospermes, angiospermes) a révélé que chez les plantes terrestres, l'organisation de ce génome circulaire est remarquablement conservée. Hormis pour certaines légumineuses et des conifères, le génome chloroplastique est caractérisé par la duplication d'une région contenant l'ADN ribosomique chloroplastique. Les gènes de l'ADN chloroplastique se répartissent en deux catégories :
L'ADN chloroplastique a un taux d'évolution assez lent. Ainsi, le séquençage de parties de l'ADN chloroplastique (comme le gène rbcL) permet de retracer l'évolution des grandes familles de plantes via des reconstructions phylogénétiques. |
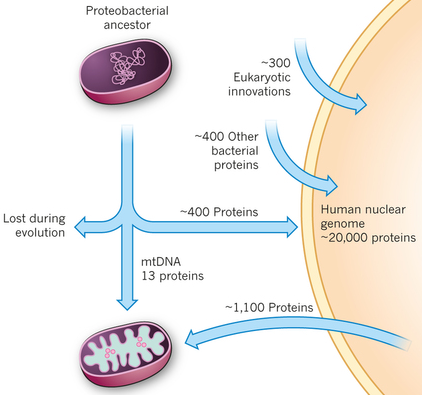
4. Arguments supplémentaires pour une origine bactérienne de la mitochondrie Le protéome mitochondrial "moderne" de l'homme semble un "vestige" du génome protéo-bactérien. Il contient 37 gènes qui codent pour 13 protéines, 22 ARN de transfert et 2 ARN ribosomiques. Remarque : 1100 autres protéines mitochondriales sont codées par l'ADN génomique.
Source : Vafai & Mootha (2012) La similarité des séquences des protéines de la mitochondrie avec celles de l'organisme vivant le plus proche des espèces ancestrales protéo-bactériennes, Rickettsia prowazekii, a été étudiée (bioinformatique). Cela a permis de déterminer que environ 800 des protéines mitochondriales codées par l'ADN génomique ont une origine protéo-bactérienne. Un point très important est qu'environ 300 protéines mitochondriales codées par l'ADN génomique n'ont aucun homologue chez les organismes procaryotes : on peut considérer ces protéines comme des innovations des eucaryotes. Les ribosomes mitochondriaux ressemblent à ceux de certaines bactéries et l'ADN polymérase mitochondriale ressemble à celle du bactériophage. La ribonucléotide réductase (synthèse des désoxy-ribonucléotides) n'est trouvée que dans le cytosol, mais la déficience de cette enzyme est la cause du syndrome de déplétion de l'ADN mitochondrial. Caractéristiques des génomes mitochondriaux des plantes
Il existe un processus propre aux mitochondries végétales : l'édition des messagers. Le transcrit primaire peut subir des modifications post-transcriptionnelles qui changent spécifiquement certaines cytidines en uraciles. Les codons de l'ARN messager sont donc modifiés. Voir le cours sur le séquençage des génomes. |
|
5. Evolution des organismes via la phylogènie du cytochrome C Visualisation du cytochrome C de Thunnus alalunga à une résolution de 1,5 Å. PDB : 5CYT Motif de fixation de l'hème (protoporphirine IX à Fe) : CXXCH
|
Le cytochrome c est une protéine de la chaîne respiratoire. Le cytochrome c existe depuis pratiquement le début de la vie sur terre et on le trouve dans la plupart des êtres vivants, (plantes, animaux, levures, bactéries). On connait un trés grand nombre de séquences du cytochrome C et d'autres cytochromes (a, a3, b, b5, b6, c1, d, f, P450, ...).
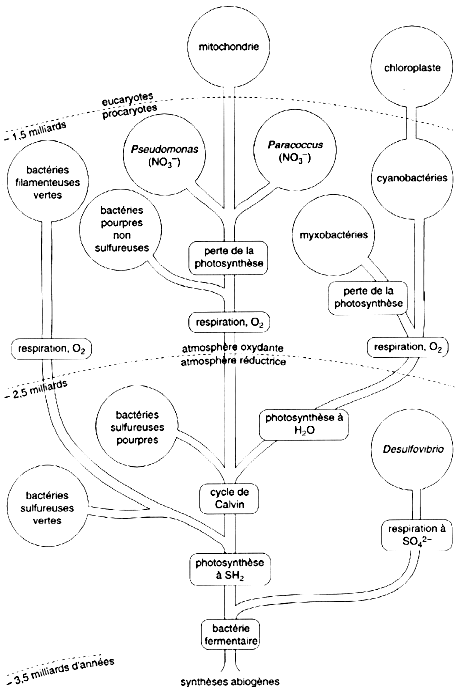
C’est donc une protéine qui se prête à des études de phylogènie. Les reconstructions d'arbres phylogénétiques par des moyens bioinformatiques, permettent de retracer l'historique de l’évolution des espèces et d'étayer la théorie endosymbiotique. La comparaison des génomes, notamment des algues, est un point capital pour conforter la théorie endosymbiotique. Figure ci-dessous : arbre, construit sur la base des séquences du cytochrome c, retraçant l'évolution des métabolismes bactériens et notamment l'apparition de la respiration bien après la photosynthèse.
Source : "Physiologie végétale" (1995) - D. Laval-Martin et P. Mazliak
|
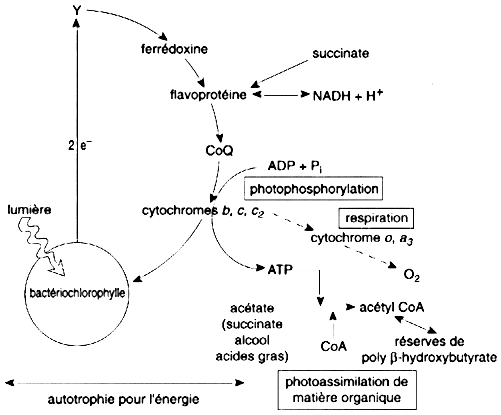
Figure ci-dessous : photo - organotrophie de la bactérie pourpre non sulfureuse Rhodospirillum rubrum.
Source : "Physiologie végétale" (1995) - D. Laval-Martin et P. Mazliak C'est un photosystème à bactériochlorophylle qui opère une circulation cyclique des électrons délocalisés par les photons captés.
Voir le transport non cyclique et cyclique des électrons des végétaux modernes. |
| 6. Liens Internet et références bibliographiques |
|
"Biologie : Licence - tout le cours en fiche" Giraud et al. (2010) - DUNOD - ISBN : 978-2-10-053449-4 J.-C. Callen - "Biologie Cellulaire" (1999), Ed. DUNOD - ISBN : 2 10 003197 X "Physiologie végétale" (1995) volume I - "Nutrition et métabolisme", Danielle Laval-Martin et Paul Mazliak, Collection "Méthodes", Hermann / ISBN : 2 7056 6253 7 |
|
|
"La théorie endosymbiotique" - (Equipe multimédia - Université Jussieu) "LEA Database" |
|
| Taylor F.J.R. (1976) "Autogenous theories for the origin of eukaryotes" Taxon 25, 377 - 390 Wegener Parfrey et al. (2011) "Estimating the timing of early eukaryotic diversification with multigene molecular clocks" Proc. Natl. Acad. Sci. U S A. 16, 13624 - 13629 |
|
Vafai & Mootha (2012) "Mitochondrial disorders as windows into an ancient organelle" Nature 491, 374 - 383 Curtis et al. (2012) "Algal genomes reveal evolutionary mosaicism and the fate of nucleomorphs" Nature Gould (2012) "Evolutionary genomics: Algae's complex origins" Nature 492, 46 - 48 |
|
Spang et al. (2015) "Complex archaea that bridge the gap between prokaryotes and eukaryotes" Nature 521, 173 - 179 Embley & Williams (2015) "Steps on the road to eukaryotes" Nature 521, 169 - 170 Zhang et al. (2025) "Deep origin of eukaryotes outside Heimdallarchaeia within Asgardarchaeota" Nature 642, 990 - 998 |
|
![]()
{kind=link}