| Porines et aquaporines |
| Tweet |
|
1. Les porines 2. Nouveau mécanisme de signalisation transmembranaire via les porines Ompf et OmpC 3. Les aquaporines |
4. Structure des aquaporines 5. Imperméabilité des aquaporines aux protons 6. Liens Internet et références bibliographiques |
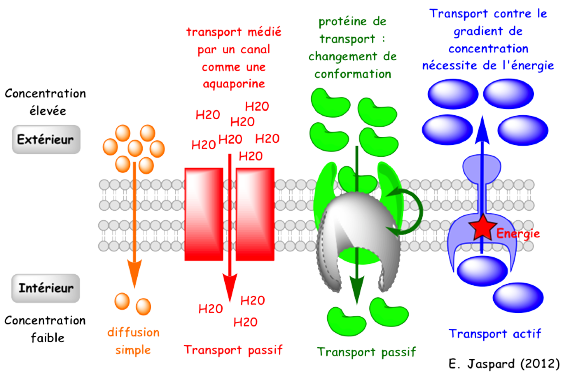
1. Les porines Les porines appartiennent à une famille de protéines membranaires intrinsèques. On les trouve dans la membrane externe des bactéries (Gram - et certaines Gram +), de la mitochondrie et du chloroplaste. Leur structure est un tonneau β ("β-barrel") constitué de feuilets β anti-parallèles. Les acides aminés polaires et non polaires alternent au sein des feuillets : les acides aminés polaires sont orientés vers l'intérieur hydrophile du pore et les acides aminés non polaires interagissent avec les lipides membranaires. Ainsi, les porines forment un pore (ou canal) dans la membrane. Elles participent donc aux transports transmembranaires. Du fait de la taille du pore, divers types de molécules (sucres, ions et acides aminés - solutés de masse molaire inférieure à 5 à 7 KDa) diffusent de manière passive. Chaque type de porine transporte sélectivement un groupe de molécules ou une molécule unique. Figure ci-contre : illustration schématique des types de transports transmembranaires.
Le canal porine est partiellement obstrué par une boucle qui détermine la taille des solutés qui peuvent traverser la porine. Cette boucle contient de nombreuses charges négative (4 Glu et 7 Asp) partiellement neutralisées par 2 atomes de calcium. |
2. Nouveau mécanisme de signalisation transmembranaire via les porines Ompf et OmpC Les porines OmpF et OmpC jouent un rôle capital dans la régulation de l’osmolarité chez certaines bactéries comme Escherichia coli. L’expression des gènes codant ces 2 porines est régulée par :
Les porines OmpF et OmpC sont des trimères en forme de tonneau β avec des canaux étroits au sein de chaque monomère qui excluent les molécules > 600 Da tout en médiant la diffusion passive de petites molécules nutriments et métabolites au travers de la membrane externe ("Outer Membrane" - OM) des bactéries Gram -. Récemment, (Housden et al., 2010) un nouveau mécanisme pour la signalisation transmembranaire a été mis en évidence. Ce mécanisme "détourne" le rôle principal des porines en translocant dans le lumen un fragment peptique intrinsèquement non structuré ("Intrinsically Unstructured Protein" - IUP) d'une colicine via les porines Ompf et OmpC. Le signal lui-même, sous la forme d'un polypeptide épitope intrinsèquement non structuré, est ainsi transféré via OmpF et OmpC et peut atteindre sa cible intracellulaire. Les colicines sont des bactériocines, toxines protéiques produites par les bactéries pour inhiber la croissance de souches bactériennes semblables ou proches. Il existe des dizaines de bactériocines (voir la base de données BACTIBASE). Les colicines ColE9 sont des nucléases (60 kDa) non spécifiques qui hydrolysent le génome bactérien. Le mécanisme d’importation de ces toxines est dicté par des domaines conservés, impliqués dans la fixation à un récepteur et dans la translocation au travers de la membrane.
Source : Housden et al., 2010
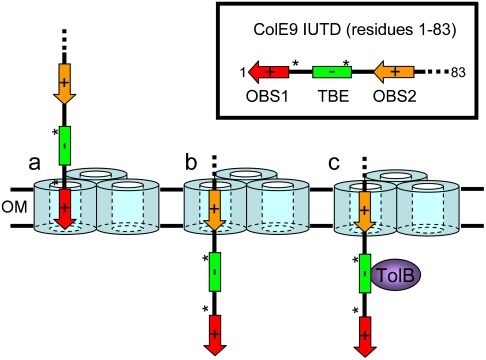
Figure ci-dessous : mécanisme de fixation séquentielle qui permet l’entrée dirigée de l’épitope au travers de la membrane externe par l’IUTD de la colicine ColE9. ColE9 possède 2 sites de fixation à OmpF : les sites OBS1 (acides aminés 2 à 18) et OBS2 (acides aminés 54 à 63) séparés par 35 acides aminés. Ces 2 sites entourent TBE.
Source : Housden et al., 2010
(a) OBS1 est guidé vers le lumen de OmpF via une association électrostatique après empilement du domaine R de la colicine sur son récepteur primaire BtuB. (b) Après dissociation de OBS1 de la porine, la diffusion de l’épitope au travers du pore est dirigée par le domaine T N-terminal extracellulaire (non montré dans la figure). Le domaine T est lui-même maintenu en place par rapport à OmpF via la formation d'un complexe entre la colicine et BtuB (non montré dans la figure). Les régions qui contiennent les prolines et la séquence TBE chargée négativement ne peuvent pas se fixer dans le pore : elles sont donc transloquées directement dans le périplasme. (c) OBS2 passe dans le pore et s’y fixe, ancrant ainsi l’IUTD dans la membrane externe : la séquence TBE se retrouve dans le périplasme. La fixation de TolB à la séquence TBE de ColE9 induit l’entrée de la colicine dans la cellule. Finalement, OBS1 adopte une conformation étendue qui traverse le canal aqueux de OmpF, de la surface extracellulaire, via la zone centrale de rétrécissement jusqu’au périplasme. |
|
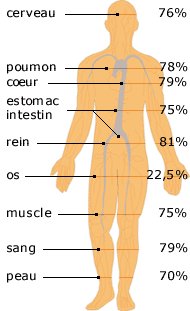
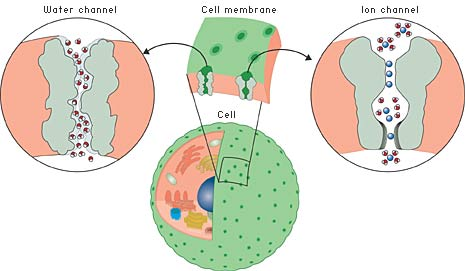
Tout être vivant est principalement constitué d'eau. En moyenne, les être humains en contiennent 65%. L'eau est indispensable à toutes les fonctions de notre organisme (maintien de la température corporelle, digestion, transit des déchets, ...). De grands volumes d'eau circulent dans notre corps de cellules en cellules. La diffusion des molécules d'eau au travers des membranes cellulaires n'explique pas la circulation de telles quantités d'eau.
Source : S. Déthiollaz (Expasy) Les aquaporines transportent sélectivement les molécules d'eau. De façon simultanée, des protons et des cations sont exclus pour maintenir le gradient ionique transmembranaire.
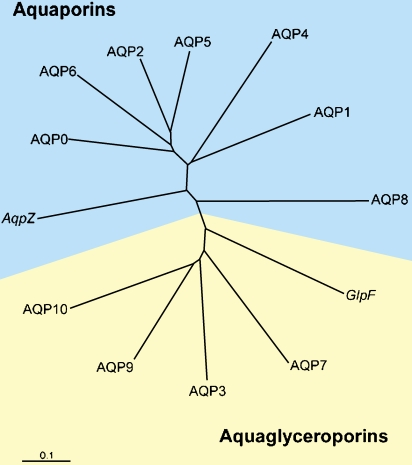
Source : "Aquaporins" Les aquaporines ont été découvertes par Peter Agre (Prix Nobel 2003). On dénombre plus de 200 aquaporines différentes, tous organismes confondus. On dénombre 13 aquaporines chez les mammifères (figure ci-dessous).
Source : P. Agre (2006) Chez les plantes, les aquaporines sont regroupées en 5 sous-familles (voir Park et al., 2010) :
Les aquaglycéroporines transportent le glycérol, le CO2, l'ammoniaque et l'urée. L'aquaporine 1 humaine est exprimée dans de très nombreux tissus : érythrocytes, tubules rénaux, coeur, rein, muscle squelettique, foie, pancréas, ... Par ailleurs on trouve majoritairement AQP0 dans la lentille de l'oeil et AQP4 dans le cerveau. Il existe 2 isoformes de AQP4 car il existe 2 sites d'initiation dela traduction (Met1 et Met 23). |
| PDB | Sous-famille | Organisme | Sélectivité |
| 1FX8 | GlpF | Escherichia coli | Glycérol, eau (faible perméabilité) |
| 1J4N | AQP1 | Bos taurus | Eau |
| 1RC2 | AqpZ | Escherichia coli | Eau |
| 1YMG | AQP0 | Bos taurus | Eau (faible perméabilité) |
| 1Z98 | PIP2 | Spinacia oleracea | Eau |
| 2F2B | AqpM | Methanothermobacter marburgensis | Glycérol, eau (faible perméabilité) |
| 2W2E | Aqy1 | Pichia pastoris | Eau |
| 3C02 | PfAQP | Plasmodium falciparum | Eau, glycérol |
| 3D9S | AQP5 | Homo sapiens | Eau |
| 3GD8 | AQP4 | Homo sapiens | Eau |
Voir Oliva et al. (2010). |
|||
|
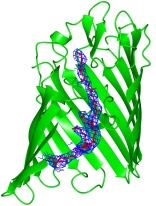
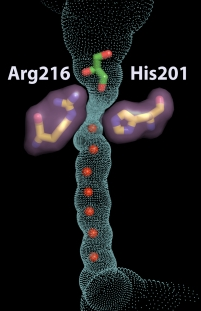
Les aquaporines sont des tétramères. Chaque monomère constitue un canal pour les molécules d'eau ou d'autres petites molécules de manière extrêmement sélective (voir tableau ci-dessus). Figure ci-dessous : exemple de l'aquaporine AQP4 humaine. Les molécules d'eau (sphères rouges) passent au travers du canal. En revanche, la molécule de glycérol (en vert et rouge en haut) ne peut pas passer la zone de constriction formée (entre autres) par His 201 et Arg 216 (équivalents tde His180 et Arg 195, citées après).
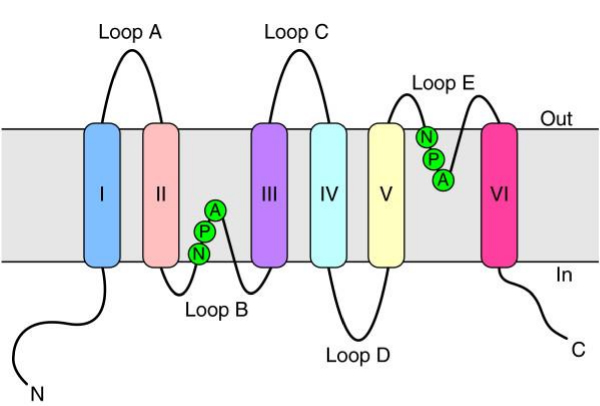
Source : Ho et al. (2009) Un monomère est constitué de 6 hélices transmembranaires et 5 boucles inter-hélices (A à E) avec les extrémités N- et C-terminales du côté cytoplasmique de la membrane (figure ci-dessous).
Source : Isokpehi et al. (2009) Les boucles A et E sont hydrophobes et contiennent un motif Asn-Pro-Ala (motif "NPA") qui recouvre le milieu de la bicouche lipidique. Ce motif joue un rôle important dans la sélectivité de l'eau. Ce motif a une forme de sablier au travers duquel passe les molécules d'eau : le diamètre du goulot est 2,8 Å, soit la place pour une seule molécule d'eau (remarque : urée et glycérol > 3.4 Å). La sélectivité s'appuie aussi sur un "filtre ar/R" (aromatique/arginine), groupement d'acides aminés qui se fixent aux molécules d'eau, excluant tout autre type de molécule (voir une animation).
Source : P. Agre (2006) Dans les aquaporines spécifiques de l'eau, les acides aminés du "filtre ar/R" sont Phe (hélice 2), His (hélice 5) et Arg (boucle E). Un 4è acide aminé (boucle E) est impliqué (Cys, Thr ou Ala). Dans les aquaporines spécifiques du glycérol (GlpF), Phe, His et le 4è acide aminé sont remplacés par Trp, Gly et Phe respectivement (Bansal et al., 2007). |
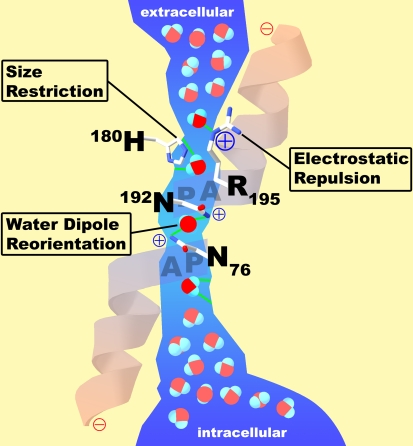
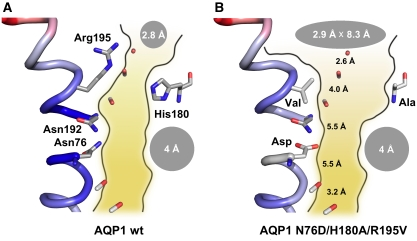
5. Imperméabilité des aquaporines aux protons Les aquaporines sont complètement imperméables aux protons et aux ions hydroxyde. Cette imperméabilité est essentielle pour le maintien du gradient électrochimique (force proton motrice) formé au cours de la chaîne respiratoire. La nature physico-chimique de cette barrière aux protons n'est pas encore clairement résolue et ce d'autant que dans bien des cas le transport des protons est médié par l'eau. Il semble qu'un des éléments moteur de cette barrière soit la perte de l'énergie de solvatation généralisée lors du transfert du proton du milieu aqueux au centre du canal. Par ailleurs, les charges positives de His 180 et de Arg 195 repoussent les protons. En effet, la mutation double H180A/R195V diminue drastiquement l'énergie libre de cette barrière de 20 kcal/mol (Chen et al., 2006) :
Source : Wu et al. (2009)
His 180, bien que trés conservée chez les aquaporines spécifiques de l'eau, est en général remplacée par une glycine dans les aqua-glycéroporines. |
|
Visualisation de l'aquaporine de Pichia pastoris à une résolution de 1,15 Å Code PDB : 2W2E Remarques :
|
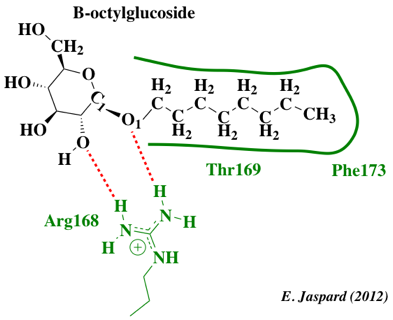
Figure ci-dessous : structure du B-octylglucoside et schéma des interactions avec certains acides aminés de l'aquaporine de Pichia pastoris.
|
| 6. Liens Internet et références bibliographiques |
|
Animation du passage d'une molécule d'eau "The porin database" : PRNDS "Structure, Dynamics, and Function of Aquaporins" "BACTIBASE : a data repository of bacteriocin natural antimicrobial peptides" Site : "Function and Structure of OmpF Porin" |
|
|
"Protéines à la Une" (Expasy") Housden et al. (2010) "Directed epitope delivery across the Escherichia coli outer membrane through the porin OmpF" Proc Natl Acad Sci U S A 107, 21412 - 21417 |
|
|
P. Agre (2006) "The aquaporin water channels" Proc. Am. Thorac. Soc. 3, 5 - 13 Isokpehi et al. (2009) "Integrative sequence and tissue expression profiling of chicken and mammalian aquaporins" BMC Genomics 10, S7 Ho et al. (2009) "Crystal structure of human aquaporin 4 at 1.8 Å and its mechanism of conductance" Proc Natl Acad Sci U S A. 106, 7437 - 7442 Park et al. (2010) "Identification of the family of aquaporin genes and their expression in upland cotton (Gossypium hirsutum L.)" BMC Plant Biol. 10, 142 Oliva et al. (2010) "Electrostatics of aquaporin and aquaglyceroporin channels correlates with their transport selectivity" Proc Natl Acad Sci U S A. 107, 4135 - 4140 |
|
|
Chen et al. (2006) "Origins of proton transport behavior from selectivity domain mutations of the aquaporin-1 channel" Biophys. J. 90, 73 - 75 Bansal et al. (2007) "Homology modeling of major intrinsic proteins in rice, maize and Arabidopsis: comparative analysis of transmembrane helix association and aromatic/arginine selectivity filters" BMC Struct Biol. 7, 27 Wu et al. (2009) "Concerted action of two cation filters in the aquaporin water channel" EMBO J. 28, 2188 - 2194 |
![]()