| Structure et fonctions de la mitochondrie |
| Tweet |
|
|
1. Introduction 2. Structure 3. Dynamique des mitochondries : fusion et fission 4. Rôle de la cardiolipine 5. Les transports au travers des membranes mitochondriales - La carnitine 6. L'ADN mitochondrial |
7. L'édition de l'ADN mitochondrial 8. Communication intracellulaire de la mitochondrie 9. Arguments pour une origine bactérienne de la mitochondrie 10. Dégradation d'agrégats cytosoliques dans la mitochondrie 11. Mitochondrie, ferroptose et dihydro-orotate déshydrogénase 12. Liens Internet et références bibliographiques |
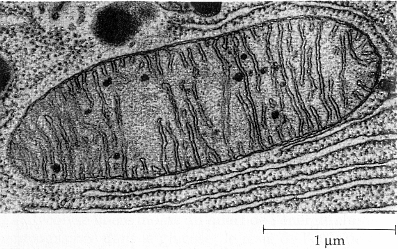
1. Introduction En condition aérobie, le pyruvate issu de la glycolyse est oxydé dans la mitochondrie. L'origine de la mitochondrie semble établie : elle dérive de l'endosymbiose d'une bactérie de la classe des α-protéobactéries dans une cellule précurseur. L'origine de la double membrane mitochondriale n'est pas élucidée. Les mitochondrie ont une structure en forme de batonnet ou de sphère de 0,5 à 1 µm de diamètre. Leur nombre est variable selon l'activité métabolique. Ce sont des organites semi-autonomes : elles possèdent leur propre génome (ADN, gènes), des ribosomes 70S, des ARN, et une trentaine de protéines y sont synthétisées directement. Le génome mitochondrial des plantes est beaucoup plus grand que chez les animaux (voir ci-après) : 195 à 2400 kilo paires de bases. La plupart de l'ADN en excès est non codant et la structure du génome semble trés fluide. |
|
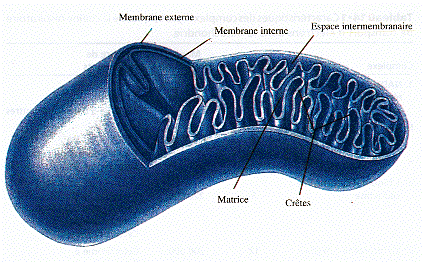
La mitochondrie est limitée par deux membranes aux propriétés très différentes :
Source : "Principes de Biochimie" Horton et al. (1994) Ed. DeBoeck Universités La membrane externe est pauvre en protéines et contient une protéine transmembranaire, la porine, qui permet le passage des ions et des métabolites hydrosolubles de masse molaire < 10.000 Da. A l'inverse, la membrane interne est très riche en protéines mais elle est quasiment impérméable aux ions et aux métabolites hydrosolubles. Ces substances ne peuvent traverser la membrane qu'à l'aide de protéines membranaires de transport (l'ATP, l'ADP et le Pi sont transportés par ce type de protéines) ou par des mécanismes plus sophistiqués qu'on appelle navette. L'espace entre ces deux membranes s'appelle l'espace intermembranaire.
Source : "Biologie" Campbell (1995) Ed. DeBoeck Universités La zone interne de la mitochondrie (bordée par la membrane interne) s'appelle la matrice. Elle contient les enzymes du cycle de Krebs et la plupart de celles qui catalysent la β-oxydation des acides gras. La chaîne respiratoire est localisée dans la membrane interne des mitochondries. Le nombre des crêtes accroit la surface de cette membrane et ainsi chaque mitochondrie contient des milliers d'exemplaires de la chaîne de transport d'électrons. Les crêtes pénètrent dans la matrice. |
|
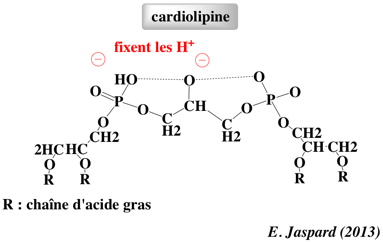
La cardiolipine (diphosphatidylglycérol) est un glycérophospholipide des membranes des bactéries et des mitochondries.
La cardiolipine représente 18 à 20% des phospholipides de la membrane interne des mitochondries. C'est un lipide caractéristique des membranes qui développent un potentiel électrochimique via le transport des électrons pour la synthèse d'ATP. La cardiolipine a de nombreux rôles :
|
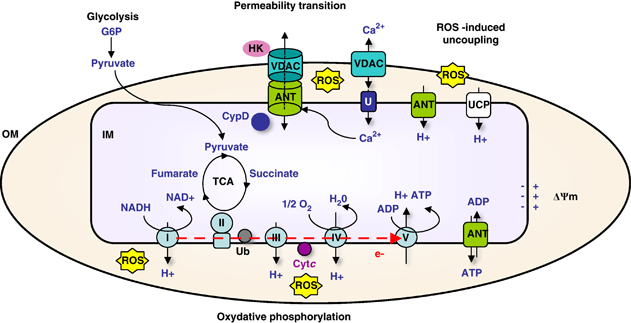
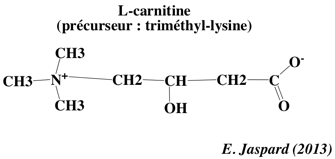
5. Les transports au travers des membranes mitochondriales - La carnitine Voir un cours sur les types de transport membranaire. Le canal ionique "Voltage-dependent anion-selective channel protein" (VDAC) est une porine qui semble la voie principale de passage des métabolites à travers la membrane externe. Il existe 3 isoformes de VDAC chez l'homme. Sa structure est en forme de tonneau β (VDAC-1 : 19 brins β, représentant une nouvelle classe de protéine membranaire). Le tonneau a un diamètre interne de 2,5 nm qui permettrait le transport des acides gras, du pyruvate, des acides aminés et des nucléotides. Le changement de conformation de ce canal anionique dépend du potentiel de membrane : ouvert pour un potentiel nul et fermé pour un potentiel de 30 - 40 mV.
Source : Brenner et al. (2011)
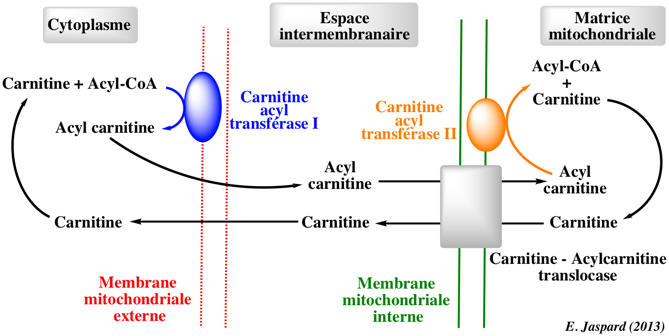
L'acide gras à transporter se fixe au coenzyme-A qui est remplacé par la carnitine pour former l'acyl-carnitine (figure ci-dessous). Cette réaction est catalysée par la carnitine O-palmitoyltransférase 1.
La carnitine est ensuite reconnue par un transporteur : la carnitine/acyl-carnitine translocase ("mitochondrial carnitine/acylcarnitine carrier protein") situé dans la membrane interne. Ce transporteur échange le complexe [acide gras-carnitine] pour une carnitine libre issue de l'intérieur de la mitochondrie. La carnitine est séparée de l'acide gras dans la matrice par la carnitine palmitoyltransférase 2 (E.C. 2.3.1.21). L'acide gras se fixe au coenzyme-A pour former un palmitoyl-CoA qui subit la β-oxydation dans la matrice. |
|
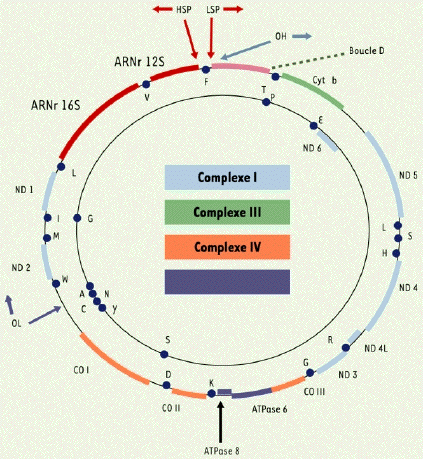
Une mitochondrie contient plusieurs copies d'ADN circulaire. L'ADN mitochondrial représente moins de 1% de l'ADN total de la cellule. L'ADN mitochondrial est compacté en agrégats appelés nucléoïdes ou mito-chromosomes. Le composant le plus abondant des nucléoïdes est le facteur de transcription A qui contribue de manière significative à la compaction de l'ADN, de manière similaire à l'action des histones. Le génome mitochondrial chez les animaux (hérité de la mère) est une molécule d'ADN double brin de de 16,569 paires de base (beaucoup plus petit que celui des plantes) avec 435 ilots CpG et 4747 cytosines dans des sites non-CpG. L'ADN mitochondrial contient 2 gènes d'ARN ribosomiques (12S et 16S) et 22 gènes d'ARN de transfert (représentés par les points dans la figure ci-dessous).
Source : May-Panloup et al. (2004) La plupart des protéines de la mitochondrie sont codées par le génome nucléaire et importées dans la mitochondrie. Cependant, certaines protéines mitochondriales sont codées par l'ADN mitochondrial :
La boucle D est une courte région de contrôle de la réplication et de la transcription qui comporte deux promoteurs de la transcription (HSP : "Heavy Strand Promoter" et LSP : "Light Strand Promoter") et une origine de réplication (OH). |
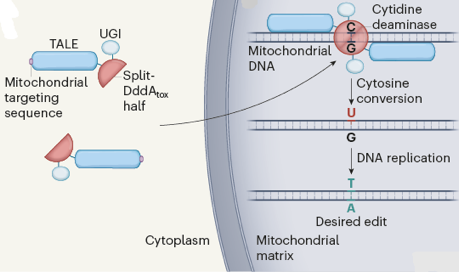
7. L'édition de l'ADN mitochondrial Un type de modification post-transcriptionnel particulier est l'édition des ARN ("RNA editing"). Cette modification affecte essentiellement les ARN messagers mais également certains ARNt mitochondriaux : une conversion de C en U corrige une paire de bases incorrecte [C:A] en une paire de bases classique [U:A]. Elle a pour conséquence un changement dans la séquence en acides aminés des protéines codées. Exemple du gène de la cytoxydase :
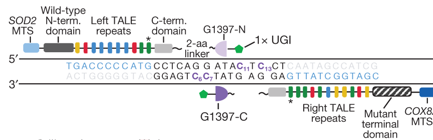
Outil moléculaire pour l'édition de l'ADN mitochondrial Une toxine sécrétée par la bactérie Burkholderia cenocepacia tue les bactéries voisines en changeant leur ADN. Cette toxine est une enzyme :
La caractéristique principale de DddA est qu'elle cible l'ADN double brin (toutes les cytidine désaminases identifiées préalablement ciblent l'ADN simple brin). De plus, les méthodes classiques d'édition des génomes utilisent des nucléases qui hydrolysent les 2 brins d'ADN : la DddA convertit C en U sans induire de cassures de l'ADN double brin. Cette enzyme est donc particulièrement bien adaptée à l'édition du génome mitochondrial, cet organite étant dépourvu de mécanismes efficaces de réparation de l'ADN double brin. Cependant, la cytidine désaminase est toxique pour les cellules de mammifères :
Le complexe [TALE - split-DddAtox] doit traverser les 2 membranes mitochondriales pour atteindre l'ADN mitochondrial dans la matrice :
La DddA convertit C en U : cependant, bien que U ait les mêmes propriétés d'appariement de bases que T, c'est un nucléotide constitutif des ARN.
La construction finale est constituée d'une séquence de signal de ciblage mitochondrial, d'une protéine TALE, d'une moitié "split-DddAtox" et d'une UGI.
Source : Mok et al. (2020)
Source : Aushev & Herbert (2020) |
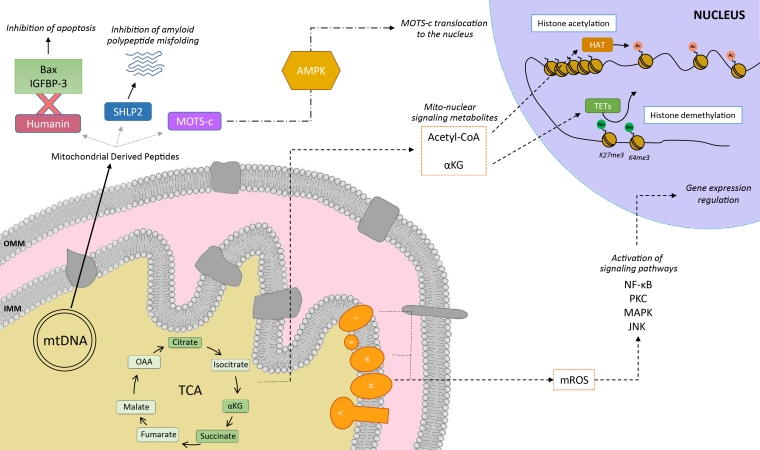
8. Communication intracellulaire de la mitochondrie Les mitochondries communiquent avec le reste des composants cellulaires via de multiples molécules de natures très diverses comme des fragments d'ADN mitochondrial, des lipides mitochondriaux (par exemple, la cardiolipine), des métabolites et des petits peptides. Ces mécanismes de communication ne sont pas forcément liés à un dysfonctionnement mitochondrial, mais utilisés comme informations sur la base de divers indices (par exemple, le flux de nutriments ou les états rédox). Les peptides dérivés des mitochondries ("Mitochondrial Derived Peptides", MDP) sont des micropeptides de signalisation codés par de courts cadres de lecture ouverts ("short Open Reading Frame", sORF) du génome mitochondrial.
Source : Valera-Alberni & Canto (2018)
Les mitochondries sont des plateformes de signalisation anti-virale et, en raison de leur origine bactérienne, l'ADN mitochondrial et d'autres composants mitochondriaux déclenchent des réponses immunitaires innées et une pathologie inflammatoire. La libération dans le cytoplasme d'ADN mitochondrial active la voie cGAS ("Cyclic GMP-AMP synthase") - STING - TBK1 qui active la transcription du gène stimulé par l'interféron qui favorise l'immunité antivirale. Ainsi, lorsqu'il est endommagé, l'ADN mitochondrial en avertit le noyau. En effet, il semble que les dommages et la libération subséquente de l'ADN mitochondrial provoquent une réponse protectrice qui augmente la réparation de l'ADN nucléaire, l'ADN mitochondrial jouant un rôle de "sentinelle de stress génotoxique". |
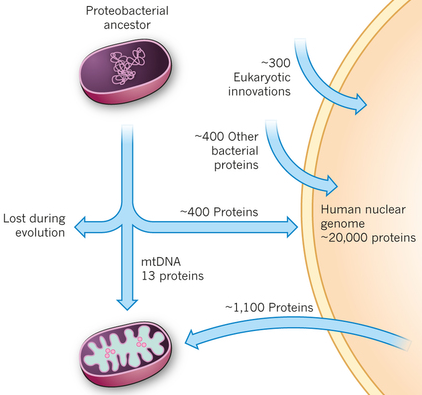
9. Arguments pour une origine bactérienne de la mitochondrie Le protéome mitochondrial "moderne" de l'homme consiste en :
Source : Vafai & Mootha (2012) La similarité des séquences des protéines de la mitochondrie avec celles de l'organisme vivant le plus proche des espèces ancestrales protéo-bactériennes, Rickettsia prowazekii, a été étudiée (bioinformatique). Cela a permis de déterminer que environ 800 des protéines mitochondriales codées par l'ADN génomique ont une origine protéo-bactérienne. Un point très important est qu'environ 300 protéines mitochondriales codées par l'ADN génomique n'ont aucun homologue chez les organismes procaryotes séquences : on peut considérer ces protéines comme des innovations des eucaryotes. Les ribosomes mitochondriaux ressemblent à ceux de certaines bactéries et l'ADN polymérase mitochondriale ressemble à celle du bactériophage. La ribonucléotide réductase (synthèse des désoxy-ribonucléotides) n'est trouvée que dans le cytosol, mais la déficience de cette enzyme est la cause du syndrome de déplétion de l'ADN mitochondrial. |
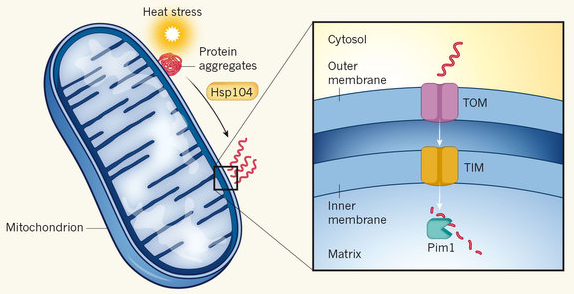
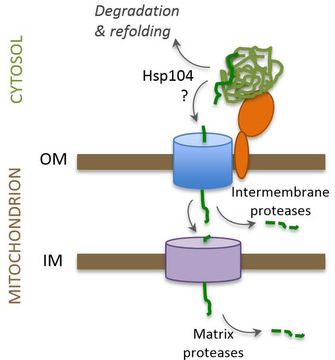
10. Dégradation d'agrégats cytosoliques dans la mitochondrie Dans la levure, les protéines cytosoliques sujettes à l'agrégation sont importées dans les mitochondries pour y être dégradées. Les agrégats protéiques formés lors d'un choc thermique entrent dans l'espace intermembranaire et dans la matrice de la mitochondrie (d'autres agrégats protéiques le font même sans stress). La dissolution rapide des agrégats cytosoliques nécessite la machinerie d'import dans la mitochondrie et des protéases :
Source : Chacinska A. (2017) Les agrégats cytosoliques sont attachés aux mitochondries par interaction avec des récepteurs de l'import tels que TOM70. Les protéines individuelles dissociées des agrégats par HSP104 sont importées par le complexe d'import de la membrane externe vers l'espace intermembranaire :
Source : Ruan et al. (2017)
|
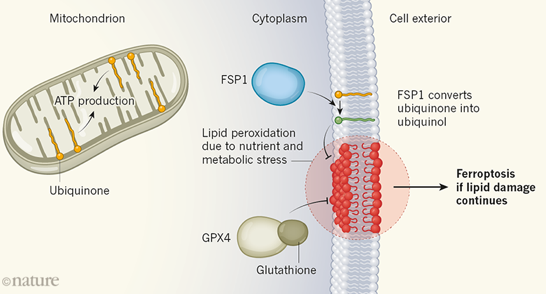
11. Mitochondrie, ferroptose et dihydro-orotate déshydrogénase La production d'énergie dépendante de l'oxygène au sein d'un organite entouré de deux membranes lipidiques a un coût : en effet, un tel processus génère des espèces réactives de l'oxygène ("Reactive Oxygen Species" - ROS) qui endommagent les structures cellulaires et compromettent leur fonction.
Les cellules de mammifères utilisent des systèmes de protection et de réparation des effets toxiques des lipides membranaires modifiés.
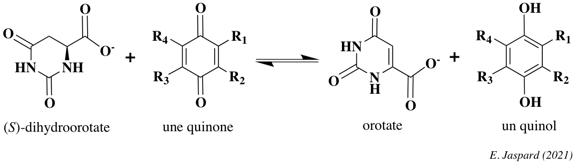
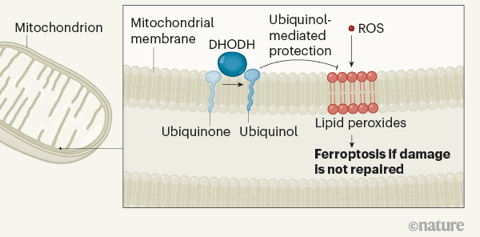
Source : B.R. Stockwell (2019) La peroxydation lipidique est associée à des changements importants de l'abondance des métabolites dans la voie de synthèse des bases pyrimidiques (composant de l'ADN et de l'ARN). La plupart des composants de cette voie de synthèse sont dans le cytoplasme mais une enzyme, la dihydro-orotate déshydrogénase (quinone) (DHODH - EC 1.3.5.2), est dans les mitochondries. Cette enzyme catalyse la conversion du dihydro-orotate en orotate par une réaction d'oxydation qui utilise l'ubiquinone et génère l'ubiquinol, permettant la réparation des dommages oxydatifs des lipides mitochondriaux.
La supplémentation des cellules avec les produits finaux de la voie de synthèse des pyrimidines n'affecte pas la peroxydation lipidique : le rôle anti-ferroptotique de la DHODH est indépendant de sa fonction dans la synthèse des pyrimidines.
Source : Garcia-Bermudez & Birsoy (2021) La DHODH synthétise donc l'ubiquinol - pour atténuer la peroxydation lipidique - exclusivement dans les mitochondries, selon un mécanisme qui est le pendant du système FSP1. |
| 12. Liens Internet et références bibliographiques |
|
"Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X |
|
|
Otera et al. (2010) "Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells" J. Cell Biol. 191, 1141 - 1158 Brenner et al. (2011) "Adenine nucleotide translocase family: four isoforms for apoptosis modulation in cancer" Oncogene 30, 883 - 895 Scott et al. (2012) "Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death" Cell 149, 1060 - 1072 Vafai & Mootha (2012) "Mitochondrial disorders as windows into an ancient organelle" Nature 491, 374 - 383 van der Bliek et al. (2013) "Mechanisms of mitochondrial fission and fusion" Cold Spring Harb Perspect Biol. 5, a011072 |
|
|
Friedman & Nunnari (2014) "Mitochondrial form and function" Nature 505, 335 - 343 Labbé et al. (2014) "Determinants and functions of mitochondrial behavior" Annu Rev Cell Dev Biol. 30, 357 - 391 Burté et al. (2015) "Disturbed mitochondrial dynamics and neurodegenerative disorders" Nat. Rev. Neurol. 11, 11 - 24 Wang & Youle (2016) "Cell biology: Form follows function for mitochondria" Nature 530, 288 - 289 |
|
|
Ruan et al. (2017) "Cytosolic proteostasis through importing of misfolded proteins into mitochondria" Nature doi:10.1038/nature21695 Chacinska A. (2017) "Cell biology: Sort and destroy" Nature 543, 324 - 325 Valera-Alberni & Canto (2018) "Mitochondrial stress management: a dynamic journey" Cell Stress 2, 253 - 274 |
|
|
Kim et al. (2019) "The mitochondrial-derived peptide MOTS-c is a regulator of plasma metabolites and enhances insulin sensitivity" Physiol. Rep. 7, e14171 Wu et al. (2019) "Mitochondrial DNA stress signalling protects the nuclear genome" Nat. Metabolism 1, 1209 - 1218 B.R. Stockwell (2019) "A Powerful Cell-Protection System Prevents Cell Death by Ferroptosis" Nature 575, 597 - 598 |
|
|
Mok et al. (2020) "A bacterial cytidine deaminase toxin enables CRISPR-free mitochondrial base editing" Nature 583, 631 - 637 Aushev & Herbert (2020) "Mitochondrial genome editing gets precise" Nature 583, 521 - 522 Garcia-Bermudez & Birsoy (2021) "A mitochondrial gatekeeper that helps cells escape death by ferroptosis" Nature 593, 514 - 515 |
|
![]()