| Les systèmes CRISPR - Cas ("Clustered Regularly Interspaced Short Palindromic Repeats - CRISPR-associated") |
| Tweet |
|
|
1. Présentation générale 2. Classification des différents systèmes CRISPR-Cas 3. Les motifs PAM ("Protospacer Adjacent Motif") de l'ADN invasif 4. Le complexe [Cas1 - Cas2] : acquisition de la séquence espaceur 5. L'endonucléase Cas9 6. Structure et mécanisme catalytique de Cas9 7. Edition des gènes avec les systèmes CRISPR-Cas9 de type II |
8. Compléments sur la technique CRISPR-Cas
9. Quelques logiciels et sites WEB dédiés à CRISPR-Cas 10. Liens Internet et références bibliographiques |
1. Présentation générale Article de Ishino et al. en 1987 : mise en évidence d'une région de 5 répétitions partiellement palindromiques dans le génome d'Escherichia Coli à l'extrémité 3' du gène codant l'alcaline phosphatase. Extrait de la fin de l'article de Ishino et al. (1987) : "An unusual structure was found in the 3'-end [...] Five highly homologous sequences of 29 nucleotides were arranged as direct repeats with 32 nucleotides as spacing. [...] A dyad symmetry with 14 nucleotide pairs was also found in the middle of these sequences [...] the biological significance of these sequences is not known." Les terminologies ont beaucoup évolué (1987 => 2002) en fonction de la compréhension du processus.
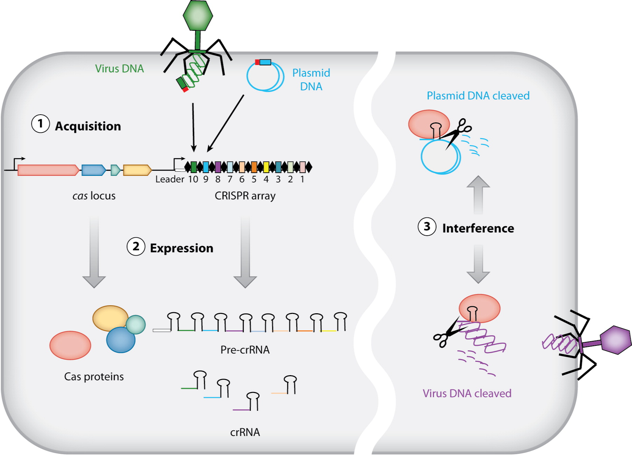
CRISPR-Cas est le premier (et jusqu'à présent le seul) système de l'immunité adaptative découvert chez les archées et les bactéries : elles acquièrent ainsi une résistance à un ADN invasif. Comme d'autres systèmes de défense, un système CRISPR-Cas est basé sur un mécanisme sophistiqué de discrimination [soi / non-soi]. Voir l'interférence ARN par comparaison. |
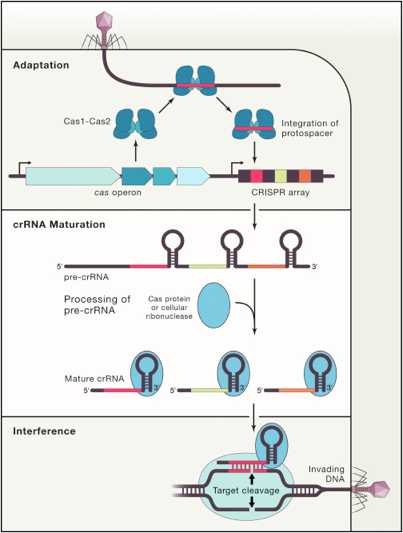
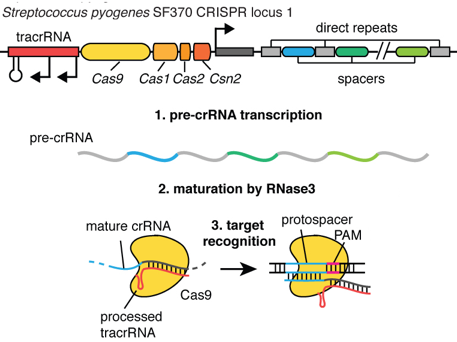
Phase d'acquisition (aussi appelée d'adaptation) : un système CRISPR-Cas conserve de courtes séquences d'ADN invasif (les séquences espaceurs - "spacers") comme éléments de mémoire des génomes des virus et des plasmides rencontrés. Phase de transcription - maturation : la matrice CRISPR (ensemble des séquences répétées - "repeats" - et des séquences espaceurs) est transcrite en un long transcrit primaire appelé pre-crRNA. Celui-ci est maturé en un ensemble de courts fragments d'ARN appelés crRNAs ("CRISPR RNAs") : chacun d'entre eux contient une séquence unique complémentaire d'un fragment d'ADN invasif. Phase d'interférence : Les crRNAs sont utilisés comme ARN guide des protéines Cas pour cibler et hydrolyser l'ADN apparenté de virus ou de plasmides invasifs lors de nouvelles infections.
Source : The Doudna Lab La séquence de tête ("leader sequence") est une longue séquence riche en AT positionnée immédiatement en amont de la matrice CRISPR. Elle contient généralement à la fois le promoteur qui commande la transcription des crRNAs et la séquence de reconnaissance pour l'insertion de la séquence espaceur. |
Une matrice CRISPR :
pre-crRNA : long transcrit primaire issu de la matrice CRISPR crRNAs : courtes séquences d'ARN issues de la maturation des séquences répétées du pre-crRNA tracrRNA ("trans-acting small RNA") : petit ARN qui s'apparie avec chaque répétition du pre-crRNA pour former un ARN double brin [tracrRNA:crRNA] séquence espaceur (porté par le crRNA mature) : séquence complémentaire de la séquence cible de l'ADN invasif (appelée proto-espaceur - "protospacer") afin qu'il y ait hybridation PAM ("Protospacer Adjacent Motif) : séquence de l'ADN invasif qui suit immédiatement la séquence cible proto-espaceur Séquence de la matrice CRISPR-Cpf1 de type V de F. novicida U112 depuis le début de la séquence de tête (minuscule - gris) jusqu'à la dernière séquence répétée.
Source : Fonfara et al. (2016) Majuscules - noir : séquences répétées; minuscules - vert : séquences espaceurs Les séquences encadrées correspondent aux crRNA matures : ils sont composés d'une partie d'une séquence répétée en 5' et d'une partie d'une séquence espaceur en 3'. |
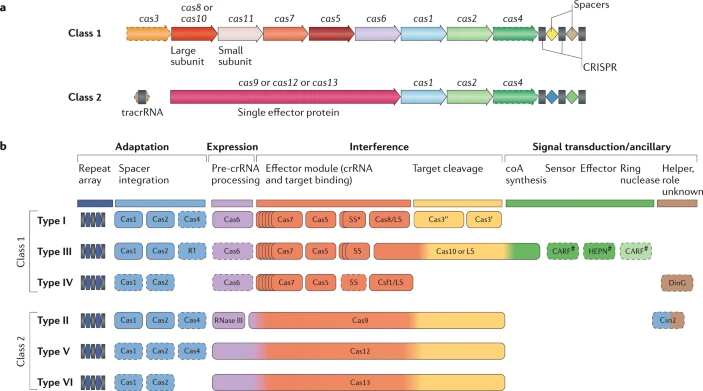
2. Classification des différents systèmes CRISPR-Cas On distingue :
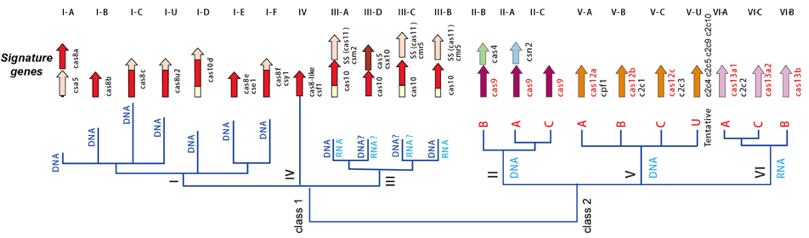
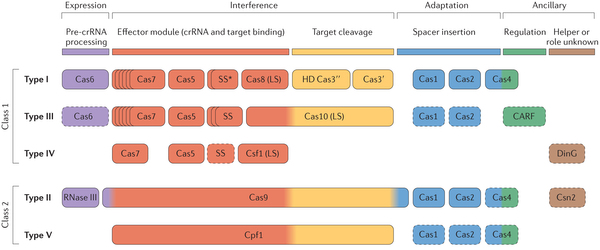
Source : Makarova et al. (2020) Il existe 6 types de systèmes CRISPR-Cas de l'immunité adaptative qui diffèrent par :
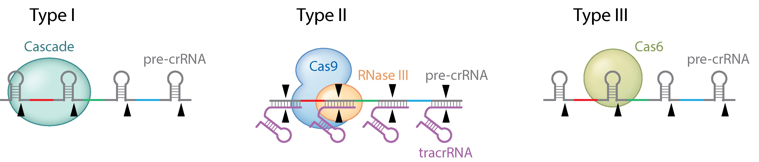
Source : Jiang & Marraffini (2015) Les systèmes CRISPR-Cas de types I utilisent des complexes multiprotéiques pour le ciblage guidé par le crRNA.
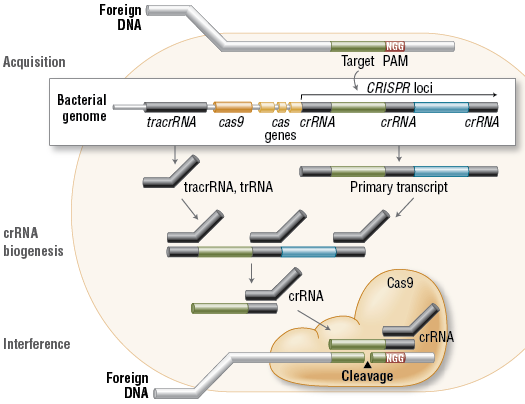
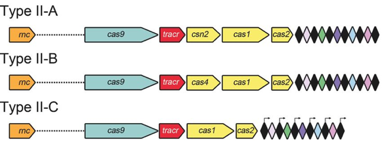
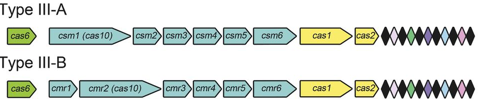
Les systèmes CRISPR-Cas de types II sont plus simples : un petit ARN agissant en trans appelé tracrRNA ("trans-acting small RNA") s'apparie avec chaque répétition du pre-crRNA pour former un ARN double brin [tracrRNA:crRNA] clivé par la RNase III en présence de l'endonucléase Cas9. Ils générent des cassures double brin dans l'ADN invasif. Les systèmes de types III sont plus complexes. Ces loci contiennent des gènes codant des protéines Cas10 et des modules Csm (pour le type III-A ) ou Cmr (pour le type III-B) qui forment ensemble les complexes de ciblage [Cas10-Csm] ou [Cas10-Cmr].
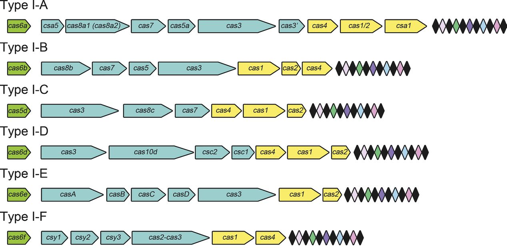
Composition des gènes cas des systèmes CRISPR-Cas de types I-A à I-F :
Source des 3 figures : Charpentier et al. (2015) Les endonucléases Cas impliquées dans la biogenèse des crRNA des systèmes de types III-A et III-B appartiennent à la famille Cas6.
|
Classe 2 : exemple du système CRISPR de type VI Ce système englobe la protéine C2c2 - Cas13a (bactérie Leptotrichia shahii) :
|
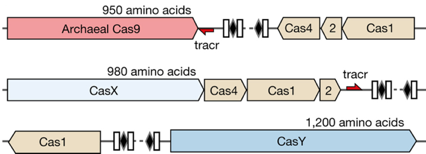
Découverte de nouveaux systèmes CRISPR-Cas La découverte de nouveaux systèmes CRISPR-Cas évolue très rapidement. Par exemple, l'étude de 155 millions de gènes codant des protéines (gènes issus d'un métagénome dérivé d'eaux souterraines, de drainages de mines, de sédiments, d'intestin de nourrisson et d'autres communautés microbiennes) a permis d'identifier :
Source : Burstein et al. (2017)
De nombreux outils bioinformatiques sont développés pour l'analyse des génomes afin de découvrir de nouveaux systèmes CRISPR-Cas et les classifier.
Source : CRISPRminer2 |
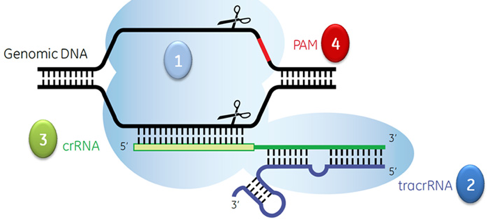
3. Les motifs PAM de l'ADN invasif Les séquences répétées ("repeats") du locus CRISPR sont séparées par de courtes séquences d'ADN non répétitif - les séquences espaceurs ("spacers") - qui proviennent de l'ADN invasif d'un plasmide ou d'un virus lors de la phase d'adaptation. Lors de l'interférence, la séquence nucléotidique espaceur (portée par le crRNA) doit être identique à une séquence du génome de ce plasmide ou de ce virus appelée proto-espaceur ("protospacer" - environ 20 nucléotides) afin que le complexe CRISPR-Cas bloque la réplication de l'ADN invasif. Dans les systèmes CRISPR-Cas de types I, de types II et de types V, il existe une séquence possèdant un motif conservé qui suit immédiatement la séquence cible proto-espaceur. Cette séquence de l'ADN invasif de 2 à 5 nucléotides est appelée motif adjacent au proto-espaceur ("Protospacer Adjacent Motif " - PAM).
Source : CRISPR Genome Engineering Resources Le motif PAM est :
|
| Espèce et type d'endonucléase Cas9 | Motif PAM - "N" désigne n'importe quel nucléotide |
| Streptococcus pyogenes (SpCas9) - Type IIA | SpCas9 reconnaît le motif PAM 5'-NGG-3' située à 3 paires de base en 3' du site de clivage sur le brin d'ADN non complémentaire. |
| SpCas9 - D1135E | NGG |
| SpCas9 - VRER | NGCG |
| SpCas9 - EQR | NGAG |
| SpCas9 - VQR | NGAN ou NGNG |
| Streptococcus mutans - Type IIA | NGG |
| Streptococcus thermophilus B - Type IIA | NNAAAAW |
| Staphylococcus aureus - SaCas9 | NNGRRT ou NNGRR(N) |
| Treponema denticola - Type IIA | NAAAAC |
| Lactobacillus buchneri - Type IIA | NAAAAN |
| Francisella novicida - Type IIB | NG |
| Neisseria meningitidis - Type IIC | NNNNGATT |
| Sources : Addgene et Anders et al. (2014) | |
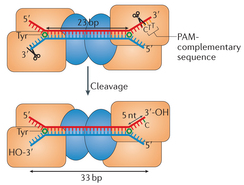
L'endonucléase Cas9 reconnaît le motif PAM au sein des séquences CRISPR répétées pour distinguer le soi du non-soi. La séparation ATP-indépendante des brins d'ADN nécessaire à la formation de l'hétéroduplexe [ARN guide - ADN cible] commence au niveau des motifs PAM. La reconnaissance des motifs PAM est donc nécessaire pour l'activité catalytique. Les deux résidus arginine du motif de reconnaissance DR1333KR1335Y de l'endonucléase Cas9 sont critiques pour la fixation du motif PAM. Le site de fixation de la séquence PAM a une structure désordonnée en absence d'ARN guide et des interactions avec le substrat. Voir la structure tridimensionnelle 4UN3. |
4. Le complexe [Cas1 - Cas2] : acquisition de la séquence espaceur Le complexe intégrase [Cas1 - Cas2] est universellement conservé : les gènes codant les enzymes Cas1 et Cas2 (qui appartiennent en général au même opéron) sont les seuls présents dans les 45 familles recencées de systèmes CRISPR-Cas. La phase d'acquisition de la séquence espaceur (courte séquence de l'ADN invasif) :
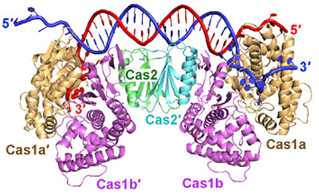
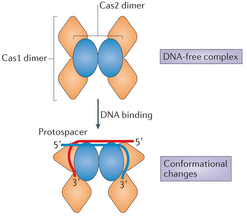
Ni Cas1, ni Cas2 ne semblent avoir un rôle dans la phase de transcription - maturation ou la phase d'interférence. L'enzyme Cas1 seule hydrolyse (activité endonucléase) : l'ARN simple brin linéaire, l'ADN simple brin et l'ADN double brin court (34 nucléotides) et des séquences d'ADN ramifiées (jonctions de Holliday, fourches de réplication, ADN double brin avec un débord 5' sur l'un des brins, ...). L'activité endonucléase de Cas2 n'intervient pas dans l'acquisition : la fonction principale de l'enzyme Cas2 est de former un échafaudage non catalytique au sein du complexe [Cas1-Cas2]. La structure du complexe [Cas1 - Cas2] de Escherichia coli lié à une séquence d'ADN proto-espaceur de 33 nucléotides a été déterminée : c'est un hexamère constitué de 4 enzymes Cas1 et 2 enzymes Cas2.
Source : Wang et al. (2015) Le complexe [Cas1 - Cas2] modifie la conformation de la séquence d'ADN proto-espaceur : cette séquence est incurvée et enjambe le dimère central Cas2. Chacune de ses extrémités nucléophiles 3'-OH entre dans un canal qui mène aux sites actifs de Cas1.
Source : Amitai & Sorek (2016) L'intégration nécessite un ADN cible super-enroulé et des extrémités 3'-OH sur l'ADN espaceur. L'intégration commence probablement par une attaque nucléophile de l'extrémité 3'-OH de la séquence proto-espaceur sur le brin négatif de la première séquence répétée. Chez Escherichia coli, la reconnaissance de PAM met en jeu une poche formée dans un dimère Cas1.
Source : Amitai & Sorek (2016) Cette poche reconnaît spécifiquement la séquence complémentaire de PAM (c'est-à-dire : 5'-CTT-3', puisque PAM de E. coli d'un système CRISPR-Cas de type I-E est 5'-AAG-3'). Cette poche positionne la séquence complémentaire de PAM dans une orientation correcte par rapport à l'histidine catalytique (His 208) du site actif de l'un des monomères Cas1. Cela conduit à l'hydrolyse de l'ADN simple brin :
|
|
Visualisation de Cas1 de Escherichia coli à une résolution de 1,95 Å Code PDB : 3NKD
|
Autres systèmes CRISPR-Cas Les systèmes CRISPR-Cas de types I et de types III adoptent une architecture en hélice qui s'enroulent autour du crRNA. Cette structure en hélice peut être le signe d'une évolution en réponse aux besoins topologiques pour fixer un hétéroduplexe [crRNA - ADN] plus long. Les systèmes CRISPR-Cas de types I (7 sous-types) contiennent tous le gène cas3. L'enzyme Cas3 s'apparente aux hélicases. L'agencement hélicoïdal ouvert du complexe Cascade (système de type I multi sous-unités) facilite probablement le recrutement de la nucléase Cas3 agissant en trans lors de l'hydrolyse de la séquence d'ADN cible. Quand la matrice CRISPR contient déjà une séquence espaceur dirigée contre un phage ou un plasmide particulier, le mécanisme d'interférence Cascade et Cas3 des systèmes CRISPR-Cas de types I peut être aussi impliqué dans l'acquisition d'une séquence espaceur. Les grandes sous-unités des systèmes de types III et des systèmes de types IV sont Cas10 et Csf1 (une enzyme de la famille Cas8), respectivement.
Source : Makarova et al. (2015) Les 2 couleurs utilisées pour Cas4 (exonucléase de type RecB) et les 3 couleurs utilisées pour Cas9 traduisent le fait que ces enzymes participent à différents stades de la réponse CRISPR-Cas.
Par exemple, Cas9 est impliquée dans les 3 phases du mécanisme CRISPR-Cas (reconnaissance PAM et acquisition d'une séquence espaceur / phase de transcription et maturation, phase d'interférence). Le domaine de fixation à l'ARN appelé RAMP ("Repeat-Associated Mysterious Protein") est présent dans les familles Cas5, Cas6, Cas7 et Cmr3. |
|
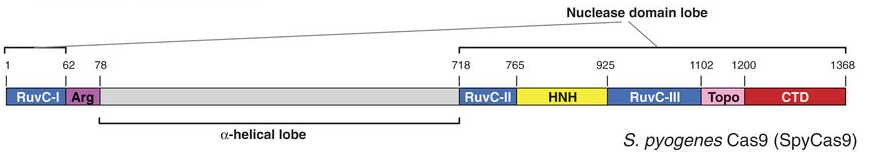
L'endonucléase Cas9 de Streptococcus pyogenes (SpyCas9 ou Csn1 - 1368 acides aminés - Q99ZW2) est spécifique des systèmes de type II-A. Elle contient deux domaine nucléase très conservés :
Autour de ces domaines, l'endonucléase Cas9 contient des régions flanquantes dépourvues de similarité de séquence apparente avec des protéines de structures connues. L'activité endonucléase sur l'ADN cible double brin nécessite le cation Mg2+.
Source : Jinek et al. (2014)
Dans les systèmes de type II, la maturation correcte du pre-crRNA nécessite le tracrRNA, la ribonucléase III et l'endonucléase Cas9.
Source : Millenium science Le tracrRNA est partiellement complémentaire du crRNA et il sert de guide pour l'action de la ribonucléase III. La longueur des séquences tracrRNAs prédites est très variable (de 72 à 171 nucléotides). Dans cette étape, l'endonucléase Cas9 stabilise l'interaction [tracrRNA:pre-crRNA]. Cas9 n'a pas de fonction catalytique dans la maturation du pre-crRNA. L'endonucléase Cas9 reconnaît le motif PAM. Cette reconnaissance est une étape critique du ciblage de l'ADN par Cas9 : c'est une condition préalable à la séparation ATP-indépendante des brins et à la formation de l'hétéroduplexe [ARN guide - ADN cible] qui commencent au niveau du motif PAM. Le complexe [Cas9:crRNA:tracrRNA] catalyse le clivage endo-nucléolytique de l'ADN double brin (linéaire ou circulaire) cible. Cas9 est inactive en absence des deux ARN guides.
Le tracrRNA est également nécessaire au mécanisme d'interférence avec l'ADN invasif. |
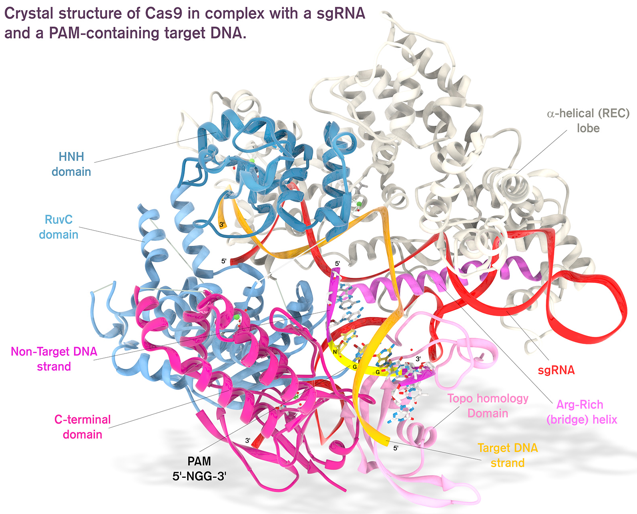
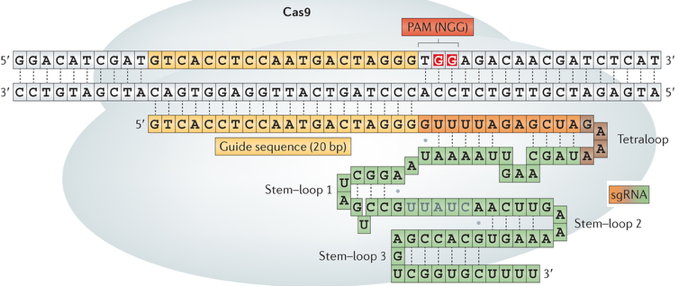
6. Structure et mécanisme catalytique de Cas9 a. Structure La structure globale de Cas9 de Streptococcus pyogenes (SpCas9) forme deux lobes : un lobe de reconnaissance (REC) et un lobe nucléase (NUC).
Structure du complexe [Cas9 / ARN guide / ADN cible] : ARN guide - rouge; ADN cible - orange; ADN non cible - fuchsia; trinucléotide PAM (5'-NGG-3') - jaune. Le lobe REC est composé des domaines REC1 et REC2 :
Aucune similitude structurale du lobe REC avec d'autres protéines n'a pour l'instant été mise en évidence : il s'agirait donc d'un domaine fonctionnel dont le repliement est spécifique de l'enzyme Cas9. Le lobe NUC est composé des domaines RuvC, HNH et CTD (domaine d'interaction avec PAM). Le lobe NUC forme une surface chargée positivement qui interagit avec l'extrémité 3' de l'ARN guide.
|
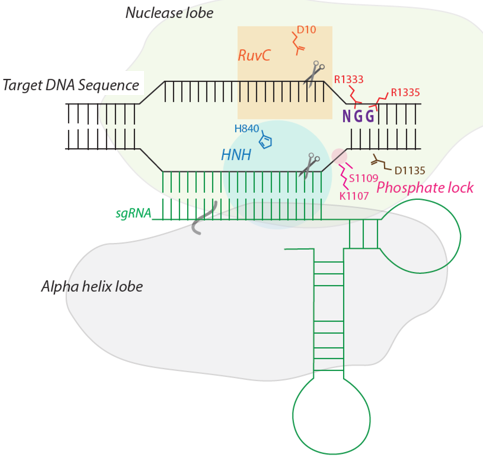
b. Mécanisme catalytique L'endonucléase Cas9 est dans une conformation auto-inhibée en absence d'acide nucléique ligand. Cas9 subit un réarrangement conformationnel très important lors de la fixation de l'ARN guide et ce réarrangement facilite la fixation de l'ADN cible. L'ARN guide déroule l'hélice d'ADN cible : l'hétéroduplexe [ARN guide - ADN cible] est ainsi formé. Cet hétéroduplexe (chargé négativement) est fixé dans une gorge (chargée positivement) à l'interface des lobes REC et NUC. Cas9 plie l'hélice d'ADN de 30° : cela crée une boucle de type R ("R-loop") qui positionne chaque brin d'ADN pour l'hydrolyse par les deux domaines nucléase. [Remarque : une boucle de type R est une structure d'acide nucléique à 3 brins, composée d'un hybride [ADN:ARN] et du brin d'ADN non matrice associé.] Les deux lobes se referment pour former un canal central (d'une largeur d'environ 25 Å) : cette réorganisation semble positionner les deux centres catalytiques de l'enzyme sur des côtés opposés de ce canal et les deux brins d'ADN séparés sont fixés dans les deux sites actifs. Les résidus Arg1333 et Arg1335 de SpCas9 reconnaissent les deux guanine du motif PAM NGG.
Source : Ishida et al. (2015) Puis Ser1109 et Lys1107 interagissent avec un groupement phosphate adjacent au motif PAM, ce qui déclenche la fusion locale de l'ADN double brin, de sorte que l'ARN guide ("sgRNA" en vert - figure ci-contre) forme un appariement de type Watson-Crick avec le brin d'ADN complémentaire.
|
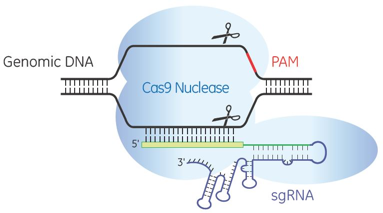
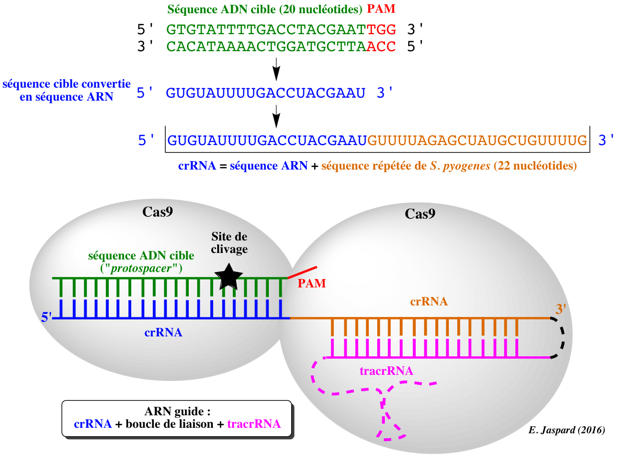
7. Edition des gènes avec les systèmes CRISPR-Cas9 de type II ("CRISPR-Cas9 genome editing") Le système CRISPR-Cas9 nécessite un ARN spécifique pour recruter et diriger l'activité nucléase de Cas9 vers la séquence d'ADN cible du gène d'intérêt. Cet ARN guide est :
Source : Dharmacon Démarche pour la conception d'un ARN guide unique Une séquence d'ADN génomique de 20 nucléotides est choisie dans le gène que l'on souhaite cibler. Par exemple, pour l'extinction d'un gène, on choisit une séquence cible située entièrement dans un exon constitutif du gène codant. Cette séquence cible doit se situer immédiatement avant le motif PAM (NGG) : la séquence cible peut donc être choisie sur l'un ou l'autre des 2 brins d'ADN à condition qu'elle soit suivie du motif PAM. L'un des avantages de la méthode dérivée de CRISPR est que le premier nucléotide de ce 20-mer peut être n'importe lequel des 4 nucléotides car le crRNA est obtenu par synthèse chimique.
Figure ci-dessous : exemple de séquence prédite d'ARN guide unique (sgRNA).
Source : Kim & Kim (2014) |
| Exemple de logiciel | But / données en entrée |
| COSMID | Searches genomes for potential off-target sites of Cas9 RNA-guided endonucleases / Designed sgRNA (10 - 55 nt) |
| Cas-OFFinder | Searches for potential off-target sites of Cas9 RNA-guided endonucleases / Designed sgRNA (10 - 25 nt) |
| sgRNA Designer | Target sequence (<10 kbp), gene ID |
|
8. Compléments sur la technique CRISPR-Cas a. Les systèmes nucléases ZNF et TALEN En raison de sa simplicité et des possibilités d'adaptation, la technique CRISPR est rapidement devenue la plus populaire pour l'ingénierie des génomes. Elle a supplanté deux techniques préalables utilisant des nucléases - enzymes de restriction artificielles :
Une nucléase ZFN est constituée de deux domaines :
Source : Sigma
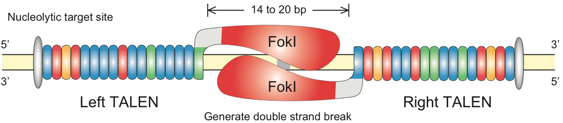
Les nucléases TALEN sont synthétisées par fusion :
Source : Genome engineering
En théorie, on peut donc créer autant de nucléases TALEN que de séquences d'ADN à hydrolyser. Cependant la synthèse du gène codant domaine de fixation à l'ADN pose des problèmes. En conséquence, la conception et la synthèse de nucléases TALEN spécifiques de chaque séquence d'ADN cible s'est avérée trop lourde et trop chère. |
b. Exemples de développements et d'applications de diverses techniques CRISPR La technique CRISPR-Cas est promise a de multiples évolutions / adaptations, ce qui laisse entrevoir une multiplicité d'applications. 1er exemple : [CRISPR-Cas9 / cytidine désaminase] Un système a été développé dans lequel la fusion [CRISPR-Cas9 / cytidine désaminase] conserve la capacité d'être dirigé par un ARN guide, n'induit pas de cassure double brin de l'ADN et convertit directement une cytidine en uridine (substitution C -> T ou G -> A) dans une fenêtre d'environ cinq nucléotides. Voir : Komor et al. (2016) 2ème exemple : dCas9 = endonucléase Cas9 désactivée dCas9 de S. pyogenes est une forme enzymatiquement inactive de Cas9 (mutation D10A dans le domaine RuvC1 et mutation H841A dans le domaine HNH). dCas9 ne peut donc pas hydrolyser l'ADN cible mais elle est toujours capable de le reconnaître et de le fixer par le biais de l'ARN guide et du motif PAM. Applications dCas9 :
Voir : Dominguez et al. (2016) - Nelles et al. (2016) 3ème exemple : le système de type V CRISPR-Cpf1 Cpf1 : CRISPR from Prevotella and Francisella 1 - nucléase d'environ 1300 acides aminés.
Voir : Sontheimer & Wolfe (2015) - Dong et al. (2016) - Fonfara et al. (2016) - Yamano et al. (2016) 4ème exemple : recherche de paires de médicament anti-cancéreux Les tumeurs deviennent souvent résistantes aux médicaments individuels. En revanche certaines combinaisons de médicaments peuvent détruire ces tumeurs résistantes. Une méthode qui inactive systématiquement 2 gènes en même temps dans les cellules permet de chercher de telles combinaisons. Cette méthode est basée sur le système d'édition de gènes CRISPR-Cas9 (« CRISPR-based double knockout »).
L'inactivation de 2 gènes (BCL2L1 et MCL1) détruit les cellules résistantes : les médicaments qui inhibent les protéines codées par ces gènes détruisent plus de cellules leucémiques que ne le font chacun des deux médicaments séparément. Voir : Han et al. (2017) |
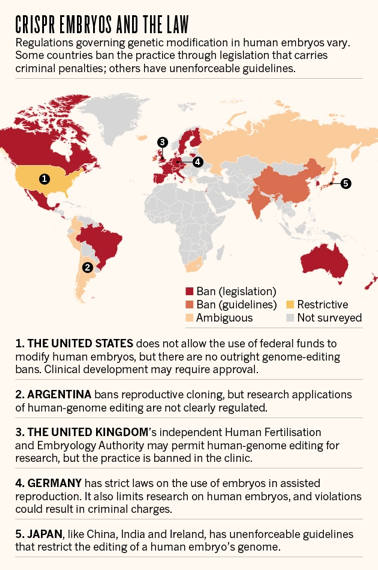
c. Les questions éthiques que soulève la technique CRISPR-Cas Certaines applications de la technique CRISPR-Cas soulève de nombreuses questions sur le plan éthique, en particulier son application sur de l'ADN humain. En 2015 et en 2017, des équipes chinoises (Liang et al., 2015 - Tang et al., 2017) ont utilisé CRISPR-Cas pour modifier le génome d'embryons humains. Cette approche a soulevé une vive controverse sur le plan éthique. Cependant, les positions évoluent très rapidement : aucun pays ne veut être à la traine en regard des possibilités d'édition des gènes des systèmes CRISPR-Cas. Exemples :
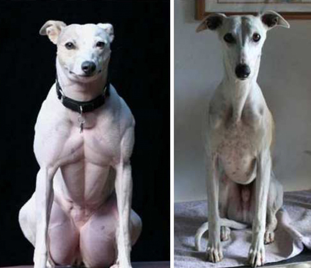
Source : Ledford H. (2015) En avril 2016, le département américain de l'agriculture a déclaré qu'il ne réglementera pas un champignon (Agaricus bisporus) génétiquement modifié avec le système CRISPR-Cas9 (Y. Yang n'a modifié que 2 nucléotides de l'ADN pour obtenir un variant plus résistant au brunissement dû à l'oxydation - enzyme polyphénol oxidase). Le champignon peut maintenant être cultivé et vendu sans passer par le processus réglementaire de l'agence. C'est le premier organisme modifié par CRISPR-Cas9 à recevoir un feu vert du gouvernement américain. Ci-dessous : organismes modifiés par le système TALEN (veau sans corne)
ou par le système CRISPR-Cas9 (chien hyper-musclé)
|
d. Bataille juridique concernant la paternité de la technique CRISPR-Cas La technique CRISPR-Cas est également source d'une terrible bataille judiciaire en ce qui concerne la paternité (donc les brevets) de certaines applications, en particulier les techniques de "base". Exemple de brevet : US8697359B1/ The Broad Institute - MIT / "CRISPR-Cas systems and methods for altering expression of gene products" F. Zhang (4/15/2014) Cette bataille est d'autant plus importante :
Février 2017 : l'Office des brevets et des marques des États-Unis (USPTO) publie un premier verdict en faveur de Broad Institute (Cambridge, Massachusetts) sur les droits à la technologie d'édition du génome. Entreprises biotechnologiques :
Ethique et controverse :
Le Prix Nobel de Chimie 2020 a été attribué à Emmanuelle Charpentier & Jennifer Doudna " for the development of a method for genome editing". |
| 9. Quelques logiciels et sites WEB dédiés à CRISPR-Cas |
| Logiciel ou site WEB | Type de prédiction |
| Cas-Designer | A bulge-allowed quick guide-RNA designer for CRISPR/Cas derived RGENs |
| CRISPResso | Analysis of CRISPR-Cas9 genome editing outcomes from deep sequencing data |
| CCTop | CRISPR/Cas9 target online predictor |
| CRISPI | CRISPI: a CRISPR interactive database |
| CRISPRmap | Provides a detailed insight into repeat conservation and diversity of bacterial and archaeal systems |
| E-CRISP | Design of CRISPR constructs |
| Protospacer | Finding, evaluating, and sharing Cas9 guide-RNA (gRNA) designs |
| WU-CRISPR | Characteristics of functional guide RNAs for the CRISPR/Cas9 system |
| FORECasT | Predictor of the mutational outcomes of a given gRNA (Favored Outcomes of Repair Events at Cas9 Targets) |
| inDELPHI | Machine learning algorithm that is aimed to assist scientists using CRISPR |
| 10. Liens Internet et références bibliographiques |
|
Quelques articles clé dans l'historique CRISPR-Cas Ishino et al. (1987) "Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product" J. Bacteriol. 169, 5429 - 5433 / Première description d'un système "CRISPR" chez une bactérie Gram-négative Hermans et al. (1991) "Insertion element IS987 from Mycobacterium bovis BCG is located in a hot-spot integration region for insertion elements in Mycobacterium tuberculosis complex strains"" Infect. Immun. 59, 2695 - 2705 / Première description d'un système "CRISPR" chez une bactérie Gram-positive Mojica et al. (1993) "Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites" Mol. Microbiol. 9, 613 - 621 / Première description d'un système "CRISPR" chez une archae Mojica et al. (2000) "Biological significance of a family of regularly spaced repeats in the genomes of Archaea, Bacteria and mitochondria" Mol. Microbiol. 36, 244 - 246 Jansen et al. (2002) "Identification of genes that are associated with DNA repeats in prokaryotes" Mol. Microbiol. 43, 1565 - 1575 Bolotin et al. (2005) "Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin" Microbiol. 151, 2551 - 2561 Barrangou et al. (2007) "CRISPR provides acquired resistance against viruses in prokaryotes" Science 315, 1709 - 1712 Horvath et al. (2008) "Diversity, Activity, and Evolution of CRISPR Loci in Streptococcus thermophilus" J. Bacteriol. 190, 1401 - 1412 Jinek et al. (2012) "A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity" Science 337, 816 - 821 Bondy-Denomy et al. (2013) "Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system" Nature 493, 429 - 432 |
|
|
Makarova et al. (2011) "Evolution and classification of the CRISPR–Cas systems" Nat. Rev. Microbiol. 9, 467 - 477 Bhaya et al. (2011) "CRISPR-Cas systems in bacteria and archaea: versatile small RNAs for adaptive defense and regulation" Annu. Rev. Genet. 45, 273 - 297 Wiedenheft et al. (2011) "Structures of the RNA-guided surveillance complex from a bacterial immune system" Nature 477, 486 - 489 |
|
|
Pennisi E. (2013) "The CRISPR Craze" Science 341, 833 - 836 Lange et al. (2013) "CRISPRmap: an automated classification of repeat conservation in prokaryotic adaptive immune systems" Nuc. Acids Res. 41, 8034 - 8044 Shah et al. (2013) "Protospacer recognition motifs: mixed identities and functional diversity" RNA Biol. 10, 891 - 899 |
|
|
Jinek et al. (2014) "Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation" Science 343 Nishimasu et al. (2014) "Crystal structure of Cas9 in complex with guide RNA and target DNA" Cell 156, 935 - 949 Sander & Joung (2014) "CRISPR-Cas systems for editing, regulating and targeting genomes" Nat. Biotech. 32, 347 - 355 Anders et al. (2014) "Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease" Nature 513, 569 - 573 Chylinski et al. (2014) "Classification and evolution of type II CRISPR-Cas systems" Nuc. Acids Res. 42, 6091 - 6105 Kim & Kim (2014) "A guide to genome engineering with programmable nucleases" Nat. Rev. Genet. 15, 321 - 334 |
|
|
Charpentier et al. (2015) "Biogenesis pathways of RNA guides in archaeal and bacterial CRISPR-Cas adaptive immunity" FEMS Microbiol. Rev. 39, 428 - 441 Jiang & Marraffini (2015) "CRISPR-Cas: New Tools for Genetic Manipulations from Bacterial Immunity Systems" Annu. Rev. Microbiol. 69, 209 - 228 Makarova et al. (2015) "An updated evolutionary classification of CRISPR-Cas systems" Nat. Rev. Microbiol. 13, 722 - 736 Nunez et al. (2015) "Foreign DNA capture during CRISPR–Cas adaptive immunity" Nature 527, 535 - 538 Wang et al. (2015) "Structural and Mechanistic Basis of PAM-Dependent Spacer Acquisition in CRISPR-Cas Systems" Cell 163, 840 - 853 Ishida et al. (2015) "Minimizing off-Target Mutagenesis Risks Caused by Programmable Nucleases" Int. J. Mol. Sci. 16, 24751 - 24771 Sontheimer & Wolfe (2015) "Cas9 gets a classmate" Nat. Biotech. 33, 1240 - 1241 |
|
|
Doench et al. (2016) "Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9" Nat. Biotechnol. 34, 184 - 191 Amitai & Sorek (2016) "CRISPR-Cas adaptation: insights into the mechanism of action" Nat. Rev. Microbiol. 14, 67 - 76 Kang et al. (2016) "Introducing precise genetic modifications into human 3PN embryos by CRISPR/Cas-mediated genome editing" J. Assist. Reprod. Genet. 33, 581 - 588 Fonfara et al. (2016) "The CRISPR-associated DNA-cleaving enzyme Cpf1 also processes precursor CRISPR RNA" Nature |
|
|
Dominguez et al. (2016) "Beyond editing: repurposing CRISPR-Cas9 for precision genome regulation and interrogation" Nat. Rev. Mol. Cell. Biol. 17, 5 - 15 Nelles et al. (2016) "Programmable RNA Tracking in Live Cells with CRISPR/Cas9" Cell 165, 488 - 496 East-Seletsky et al. (2016) "Two distinct RNase activities of CRISPR-C2c2 enable guide-RNA processing and RNA detection" Nature 538, 270 - 273 Yamano et al. (2016) "Crystal Structure of Cpf1 in Complex with Guide RNA and Target DNA" Cell S0092-8674 |
|
Dong et al. (2016) "The crystal structure of Cpf1 in complex with CRISPR RNA" Nature 532, 522 - 526 Komor et al. (2016) "Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage" Nature doi: 10.1038 Burstein et al. (2017) "New CRISPR–Cas systems from uncultivated microbes" Nature 542, 237 - 241 |
|
Tang et al. (2017) "CRISPR/Cas9-mediated gene editing in human zygotes using Cas9 protein" Mol Genet Genomics Han et al. (2017) "Synergistic drug combinations for cancer identified in a CRISPR screen for pairwise genetic interactions" Nature Biotech. Hille et al. (2018) "The biology of CRISPR-Cas: backward and forward" Cell 172, 1239 - 1259 Allen et al. (2019) "Predicting the mutations generated by repair of Cas9-induced double-strand breaks" Nat. Biotechnol. 37, 64 - 72 |
|
Makarova et al. (2020) "Evolutionary classification of CRISPR-Cas systems: a burst of class 2 and derived variants" Review Nat. Rev. Microbiol. 18, 67 - 83 |
|
Le Prix Nobel de Chimie 2020 a été attribué à Emmanuelle Charpentier & Jennifer Doudna " for the development of a method for genome editing". |
|
![]()