| Métabolisme des bases puriques, des bases pyrimidiques et des nucléotides |
| Tweet |
|
|
1. Présentation générale 2. Rappels sur les bases azotées, les nucléosides et les nucléotides (et leurs pendants désoxy-) 3. Le 5-phospho-α-D-ribosyl 1-pyrophosphate : point de départ des synthèses des purines et pryrimidines 4. Biosynthèse des nucléotides de purine
|
5. Régulation de la synthèse des purines 6. Dégradation des nucléotides de purine
7. Biosynthèse des nucléotides de pyrimidine
8. Régulation de la synthèse des nucléotides de pyrimidines 9. Catabolisme des nucléotides de pyrimidines 10. Liens Internet et références bibliographiques |
1. Présentation générale Dans la cellule, les bases puriques et pyrimidiques sont presque toujours des éléments constitutifs des nucléotides. Les nucléotides entrent bien évidemment dans la composition des acides nucléiques. Mais leurs rôles sont extrêmement divers car certains nucléotides :
La plupart des organismes et des cellules sont capables de métaboliser les nucléotides. Schématiquement : a. La biosynthèse des nucléotides de purine s'effectue :
Exemple de réaction de sauvetage catalysée par l'Adénine PhosphoRibosyl Transférase (APRT) : AMP + diphosphate ===> adénine + 5-phospho-α-D-ribose 1-diphosphate
b. La biosynthèse de novo des nucléotides de pyrimidine est une voie métabolique dont les caractéristiques chimiques diffèrent de celle de la biosynthèse des nucléotides de purine. c. Le catabolisme des purines diffère de celui des pyrimidines :
|
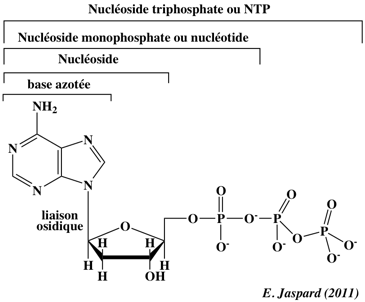
| 2. Rappels sur les bases azotées, les nucléosides et les nucléotides (et leurs pendants désoxy-) |
| Les bases azotées constitutives des acides nucléiques (voir ci-dessous la structure de ces bases azotées) | |
| purine | pyrimidine |
| adénine (A) - ADN & ARN | cytosine (C) - ADN & ARN |
| guanine (G) - ADN & ARN | thymine (T) - ADN |
| hypoxanthine - ARN | uracile (U) - ARN |
| Application : script (Python) qui dénombre les nucléotides d'une séquence et calcule la température de fusion (Tm). | |
Exemples d'autres purines dont la plupart n'interviennent pas dans la composition des acides nucléiques :
|
|
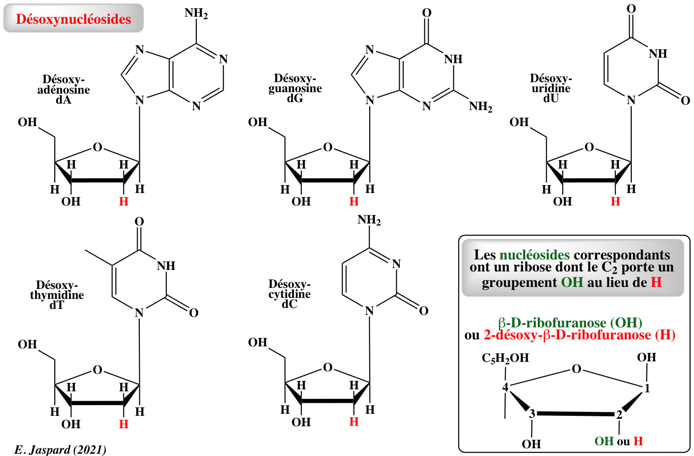
Un nucléoside est une glycosylamine : c'est-à-dire une base azotée dont un groupement amine est lié à un carbohydrate (ribofurannose) par une liaison β-N-osidique. Un désoxynucléoside utilise le 2-désoxyribofurannose. Un nucléotide résulte de la phosphorylation d'un groupement -OH du ribose du nucléoside. C'est un ester-phosphate de nucléoside. Les nucléosides diphosphates (NDP) ou les nucléosides triphosphate (NTP) sont des polyacides forts qui peuvent libérer 3 ou 4 protons (acides polyprotiques).
L'ATP (adénosine 5'-triphosphate) est un NTP particulier puisque c'est la molécule réservoir énergétique de la cellule (le GTP l'est aussi mais il est moins fréquemment utilisé).
|
| Homéostasie de la concentration optimale des nucléotides |
| Concentrations des désoxyribo- et des ribo-nucléotides mono- ou tri-phosphates dans la cellule (mM) | |||||
| Bactéries | Homme | Bactéries | Homme | ||
| dNMP - total | 43 | ----- | rNMP - total | 270 | ----- |
| dATP | 0,18 | 0,013 | ATP | 3,00 | 2,80 |
| dGTP | 0,12 | 0,005 | GTP | 0,92 | 0,48 |
| dCTP | 0,07 | 0,022 | CTP | 0,52 | 0,21 |
| dTTP | 0,08 | 0,023 | UTP | 0,89r | 0,48 |
Il existe un grand nombre de processus qui y contribuent à cette homéostasie. On peut citer 3 réactions capitales de phosphorylation :
La charge énergétique adénylique (CEA) de la cellule a été définie (Atkinson, DE, 1968) par la relation : [ATP]
+ 1/2 [ADP] Cette relation exprime la fraction molaire en ATP (qui contient 2 liaisons phosphoanhydride à haut potentiel énergétique) plus la moitié de la fraction molaire en ADP (qui n'en contient qu'une). La CEA est donc une mesure de l'énergie disponible à un instant donné dans une cellule, un tissu ou un organisme. Il s'agit d'un paramètre de valeur universelle atteignant des valeurs comparables chez tous les organismes vivants. La CEA est très sensible aux variations de l'environnement interne des organismes (stress) ou de l'environnement extérieur : plus un organisme subit l'effet d'un stress, plus il consomme d'énergie (donc d'ATP) pour contre-balancer ces effets, plus la CEA de cet organisme baisse. En théorie, la valeur de la CEA peut varier de 0 (il n'y a que de l'AMP) à 1 (il n'y a que de l'ATP). Cependant, dans la plupart des cellules, la CEA est trés finement régulée et on observe des variations de 0,7 (forte hydrolyse de l'ATP et de l'ADP en AMP) à 0,95 (typiquement une cellule au repos). |
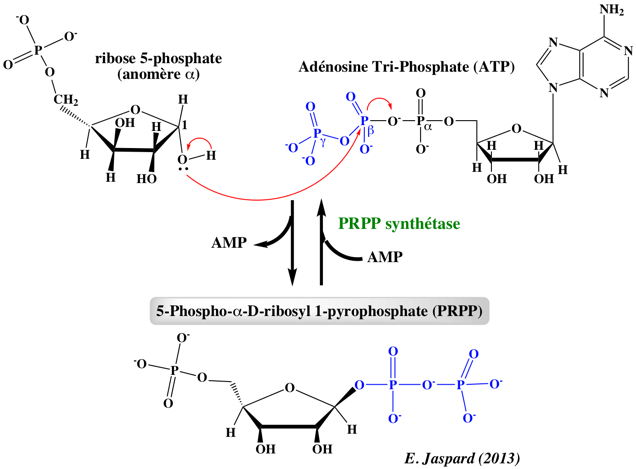
3. Le 5-phospho-α-D-ribosyl 1-pyrophosphate : point de départ des synthèses des purines et pryrimidines Tant la biosynthèse de novo des purines et des pyrimidines que la voie de sauvetage des purines commencent par l'utilisation d'un sucre activé : le 5-phospho-α-D-ribosyl 1-pyrophosphate ou 5-PRPP. La synthèse du 5-PRPP s'effectue à partir du ribose 5-phosphate et de l'ATP (donneur de groupement pyrophosphate et d'énergie).
Elle est catalysée par la ribose-phosphate pyrophosphokinase ou phosphoribosyl pyrophosphate synthétase (E.C. 2.7.6.1). Cette enzyme est inhibée par les nucléotides de purine ADP et GDP. Le 5-PRPP est un carrefour métabolique :
|
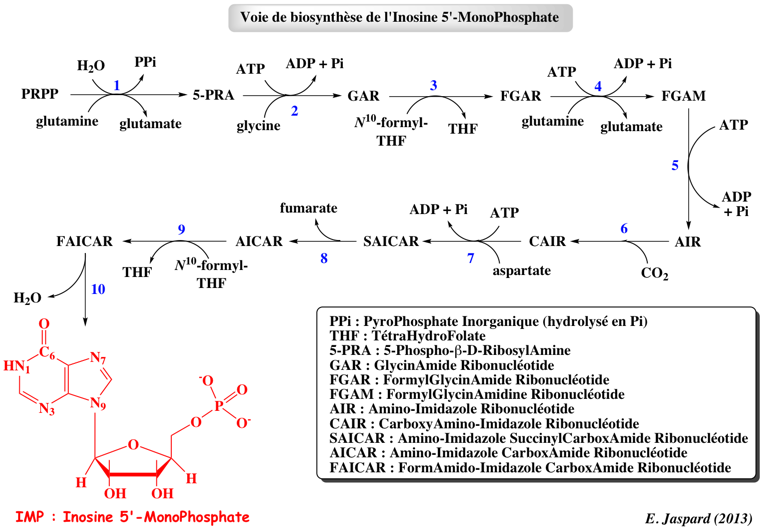
4. Biosynthèse des nucléotides de purine a. Les réactions du 5-PRPP à l'inosine monophosphate Le principal site de la biosynthèse des nucléotides de purine est le foie.
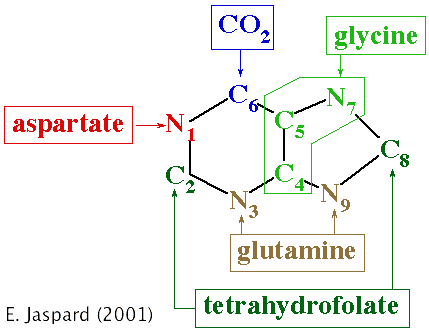
Réaction 1 : L'Amido PhosphoRibosyl Transférase catalyse la 1ère étape qui implique le 5-PRPP :
Réaction 2 : Le groupe aminé du produit, la 5-phospho-β-D-ribosylamine est acylé par la glycine pour former la glycinamide ribonucléotide. Cette réaction, catalysée par la phosphoribosylamine-glycine ligase est proche de celle catalysée par la glutamine synthétase (intermédiaire γ-glutamyl phosphate). Réaction 3 : Le 10-formyltétrahydrofolate cède son groupe formyle au groupe aminé destiné à former l'azote N-7 de l'IMP (produit final de cette voie). Cette réaction est catalysée par la phosphoribosylglycinamide formyltransférase. Réaction 4 : L'amide est transformée en amidine (R)HN-C=NH avec la glutamine comme donneur d'azote. Cette réaction est catalysée par la phosphoribosylformylglycinamidine synthase. Réaction 5 : Le cycle est fermé aux dépens de l'hydrolyse de l'ATP. Cette réaction est catalysée par la phosphoribosylformylglycinamidine cyclo-ligase ou phosphoribosyl-amino-imidazole synthétase. Les étapes 2, 3 et 5 sont catalysées par une enzyme multi-fonctionnelle qui catalyse les réactions correspondant aux activités enzymatiques EC 6.3.4.13, EC 2.1.2.2 et EC 6.3.3.1. Réaction 6 : Le carbone du CO2 (sous forme de bicarbonate) est attaché au carbone qui va devenir le C-5 du noyau purine : cette carboxylation ne dépend ni de la biotine ni de l'ATP. L'incorporation de gaz carbonique est très rare dans le métabolisme des animaux. L'atome C-5 du noyau imidazole est un nucléophile activé car il fait partie d'une ènamine [H2N-C=C] dont le -NH2 est équivalent à un -OH d'un énol. Cette réaction est catalysée par la phosphoribosylamino-imidazole carboxylase. Chez Escherichia coli, 2 enzymes distinctes sont nécessaires pour catalyser cette réaction. Réaction 7 : Le groupe aminé de l'aspartate est introduit dans le cycle purique en cours de biosynthèse. L'aspartate se condense en entier pour former d'abord un succinyli-carboxamide. Cette réaction est catalysée par la phosphoribosylaminoimidazole-succinocarboxamide synthase. Les étapes 6 et 7 sont catalysées par une 2ème enzyme multi-fonctionnelle qui catalyse les réactions correspondant aux activités enzymatiques EC 4.1.1.21 et EC 6.3.2.6. Le gène qui code cette seconde multienzyme est sous le contrôle du même promoteur que le gène qui code l'amidophosphoribosyl transférase (réaction 1 et enzyme-clé de la voie de biosynthèse de l'IMP). Il y a donc une régulation coordonnée au niveau de la transcription de ces deux gènes. Réaction 8 : L'adénylosuccinate lyase catalyse le clivage non hydrolytique qui élimine le fumarate. Ce processus aboutit au transfert d'un groupe aminé qui formera l'azote N-1 de l'IMP. Réaction 9 : Elle rappelle l'étape 3 : le 10-formyltétrahydrofolate cède son groupe formyle au groupe aminé nucléophile du ribonucléotide d'amino-imidazole-carboxamide. Cette réaction est catalysée par la phosphoribosylaminoimidazolecarboxamide formyltransférase. Réaction 10 : L'azote amide de cet intermédiaire final se condense au groupe formyle et ferme le cycle : l'inosine monophosphate est synthétisée. La base purique de l'IMP est l'hypoxanthine. Cette réaction est catalysée par l'IMP cyclohydrolase. Les étapes 9 et 10 sont catalysées par une 3ème enzyme multi-fonctionnelle qui catalyse les réactions correspondant aux activités enzymatiques EC 2.1.2.3 et EC 3.5.4.10. Bilan : la synthèse de l'IMP nécessite 5 ATP, 2 glutamines, 1 glycine, 1 molécule de CO2, 1 aspartate et 2 molécules d'acide formique. |
|
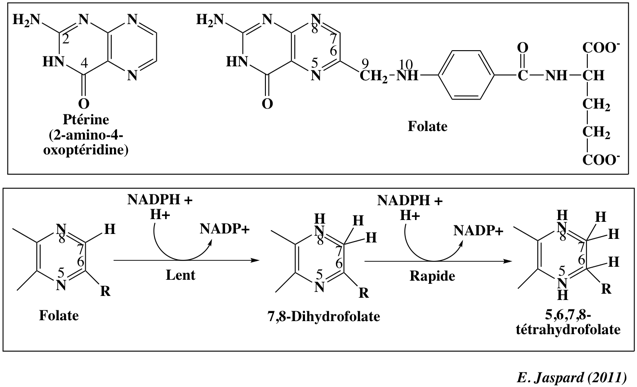
Le folate est composé de la ptérine, d'un résidu acide p-aminobenzoïque et d'un résidu glutamate (à droite dans la structure). Le tétrahydrofolate dérive du folate par addition d'atomes d'hydrogène en position 5, 6, 7 et 8 de la ptérine.
c. Le domaine catalytique des Glutamine Amido Transférases (GATase) De très nombreuses enzymes biosynthétiques catalyse le clivage de l'ammoniac de la glutamine, puis de transférer ce groupe à un substrat pour former un nouveau groupe carbone-azote. Cette activité catalytique est connue sous le nom de Glutamine Amido Transférase ou GATase. Le domaine structural qui porte cette activité catalytique peut-être :
Il existe 2 classes de domaines GATase : la classe-I (ou type trpG ou triade) et la classe II (ou type purF ou Ntn). Exemples d'enzymes qui ont un domaine GATase de la classe II :
Le site actif du domaine GATase de la classe II est constitué par une Cys située à l'extrémité N-terminale de la chaîne polypeptidique des formes matures. La position N-terminale et le repliement autour de cette Cys catalytique diffèrent de la triade [Cys-His-Glu] du site actif du domaine GATase de la classe I. Deux autres acides aminés conservés (Asn et Gly) forment un trou oxyanion qui stabilise l'intermédiaire tétraédrique. L'architecture 3D du domaine GATase de la classe II correspond à 4 couches [α/β/β/α]. Ce repliement est semblable à celui des hydrolases nucléophiles d'acides aminés N-terminaux qui sont capables d'attaque nucléophile et d'auto-catalyse. |
|
Visualisation de la Glutamine PhosphoRibosylPyroPhosphate (5-PRPP) Amido Transférase de Escherichia coli à une résolution de 2,4 Å. Code PDB : 1EEC
|
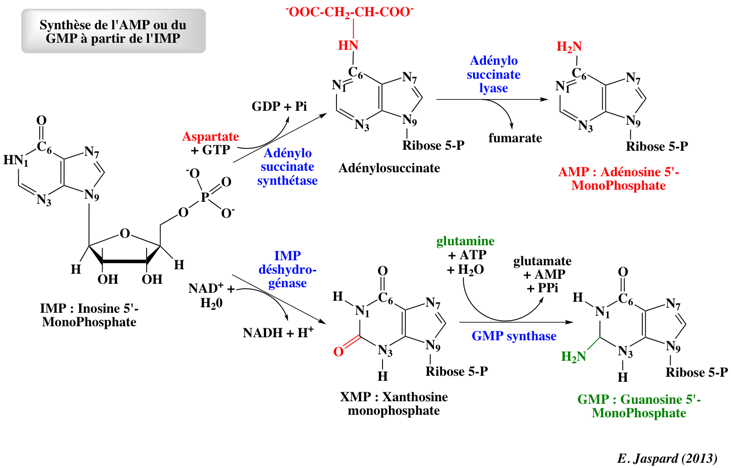
d. Synthèse de l'AMP ou du GMP à partir de l'IMP L'IMP est un point de branchement dans la biosynthèse des nucléotides de purine car il peut être converti en AMP ou en GMP via 2 suites de réactions distinctes.
L'utilisation du GTP pour la synthèse d'AMP permet à la cellule d'ajuster les proportions d'AMP et de GMP à des niveaux à peu près équivalent :
e. La voie de sauvetage des nucléotides de purine Au cours de la voie de récupération, les nucléotides de purine sont synthétisés à partir des intermédiaires (les bases puriques et les nucléosides) du catabolisme (dégradation) des nucléotides. Les bases puriques libres (adénine, guanine et hypoxanthine) peuvent ainsi être re-converties en leur nucléotide correspondant par phosphoribosylation. Ce processus est nettement moins coûteux sur le plan énergétique que la biosynthèse de novo : la synthèse d'AMP par exemple nécessite l'hydrolyse de 7 liaisons à haut potentiel énergétique. Deux transférases clé sont impliqués dans le sauvetage des purines :
Une autre enzyme joue un rôle crucial de sauvetage des purines dans les cellules à division rapide : l'adénosine désaminase qui catalyse la désamination de l'adénosine en inosine. |
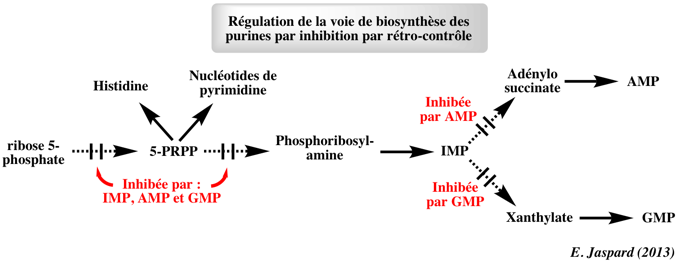
5. Régulation de la synthèse des purines La biosynthèse des nucléotides est régulée par rétro-inhibition ou inhibition par rétro-contrôle ("feedback inhibition") de manière similaire à la régulation de la biosynthèse des acides aminés.
Cette inhibition régule la vitesse de biosynthèse des purines et l'équilibre entre la formation d'AMP et de GMP. Or ces deux métabolites (et d'autres nucléotides) sont des rétro-inhibiteurs. Les deux premières réactions de la voie de biosynthèse des purines sont les étapes dont les vitesses sont limitantes. a. La synthèse du 5- PRPP par la phosphoribosyl pyrophosphate synthétase est rétro-inhibée par les nucléotides de purine (surtout par l'AMP et le GMP). Les effets de ces 2 nucléotides sont synergiques : l'inhibition est maximale quand la concentration optimale d'AMP et de GMP sont atteintes. b. La vitesse de catalyse de la glutamine phosphoribosylpyrophosphate amidotransférase, enzyme qui catalyse la formation de la phosphoribosyl-amine (5-PRA) à partir du 5-PRPP, est finement régulée par de nombreux ribonucléotides de purine :
c. Le GTP est un substrat de la synthèse de l'AMP et l'ATP est un substrat de la synthèse du GMP : cette réciprocité de substrat tend à équilibrer la synthèse des ribonucléotides d'adénine et de guanine. d. La synthèse d'AMP et de GMP est contrôlée par rétro-inhibition :
e. La réduction des ribonucléotides en désoxyribonucléotides est catalysée par la ribonucléotide réductase (E.C. 1.17.4.1). La ribonucléotide réductase forme un hétérodimère d'une grande (RRM1) et d'une petite (RRM2) sous-unité.
Cette sophistication de la régulation du flux global d'une voie métabolique complexe est un splendide cas d'école pour des études d'enzymologie. |
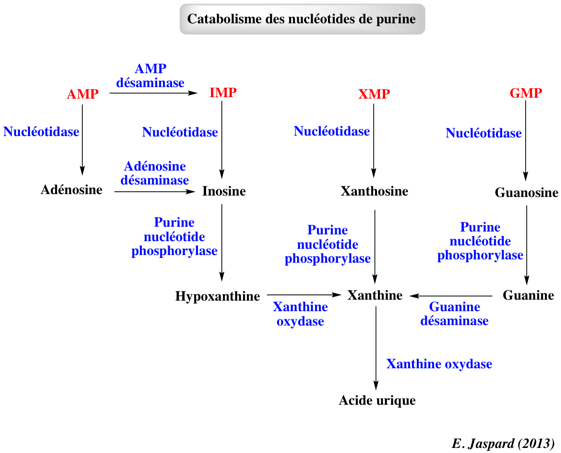
6. Dégradation des nucléotides de purine Le catabolisme des nucléotides de purine aboutit à la formation d'acide urique, insoluble et excrété dans l'urine sous forme de cristaux d'urate de sodium.
L'origine des atomes de l'acide urique Des expériences utilisant des précurseurs marqués par des isotopes radioactifs ont permis de déterminer l'origine des atomes de carbone et d'azote constitutifs de l'acide urique. Ces expériences ont montré que l'acide urique et les purines qui entrent dans la composition des acides nucléiques sont issus des mêmes précurseurs et de la même voie métabolique.
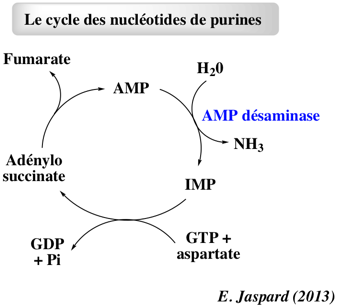
b. Le cycle des nucléotides de purine et son rôle dans l'activité musculaire La synthèse d'AMP à partir d'IMP et la voie de sauvetage d'IMP via le catabolisme d'AMP ont pour conséquence de désaminer l'aspartate en fumarate. Ce processus est appelé cycle des nucléotides de purine.
Ce cycle est trés important dans les cellules du muscle : une forte activité musculaire augmente l'activité du cycle de Krebs afin de produire davantage de NADH pour une synthèse accrue d'ATP. Le muscle reconstitue donc les intermédiaires du cycle de Krebs à partir du fumarate généré par le cycle des nucléotides de purine. |
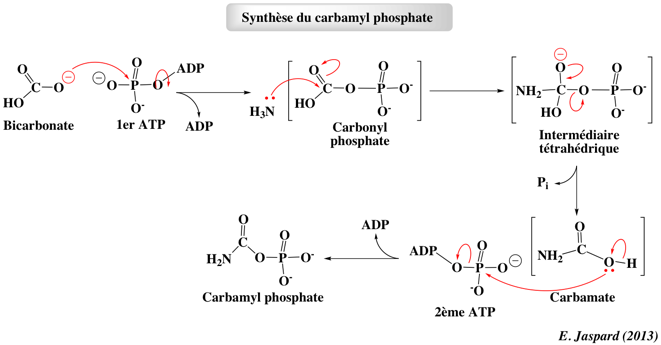
7. Biosynthèse des nucléotides de pyrimidine a. Synthèse du carbamyl-phosphate La synthèse des pyrimidines est moins complexe que celle des purines. En premier lieu, le carbamyl-phosphate est synthétisé par la carbamyl-phosphate synthétase à partir de bicarbonate, de NH3 et d'ATP.
Au delà de la biosynthèse des nucléotides de pyrimidine, cette réaction est la première étape de la biosynthèse de l'arginine (procaryotes et eucaryotes) et du cycle de l'urée chez la plupart des vertébrés terrestres. |
| Exemple de différentes carbamyl-phosphate synthétases | ||
| Carbamoyl-phosphate synthase (CPSase I) | la source de NH3 est l'ammoniac dans le cas du cycle de l'urée dans la mitochondrie | E.C. 6.3.4.16 |
| Carbamoyl-phosphate synthase glutamine-hydrolysing (CPSase II) | la source de NH3 est la glutamine dans le cytosol | E.C. 6.3.5.5 |
| Carbamoyl-phosphate synthase large chain | Escherichia coli | E.C. 6.3.5.5 |
| Carbamoyl-phosphate synthase arginine-specific large chain | P03965 | E.C. 6.3.5.5 |
| Carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase | P27708 | E.C. 6.3.5.5 |
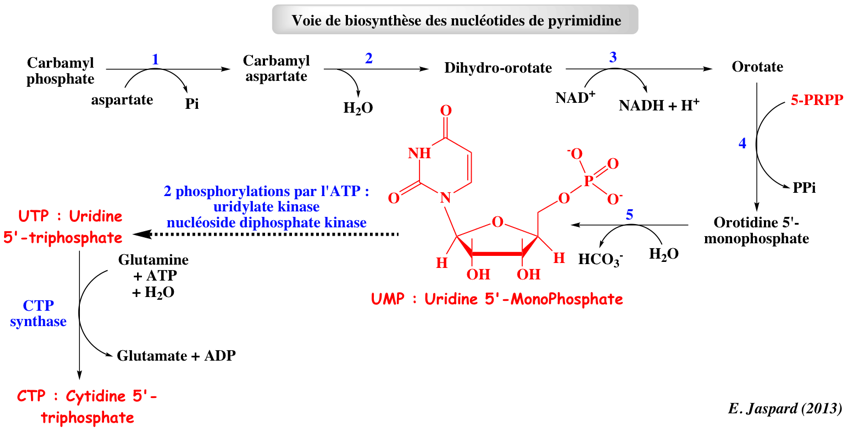
b. Suite de la synthèse des nucléotides de pyrimidine La synthèse des pyrimidines diffère sur deux points importants de celle des purines :
Réaction 1. L'aspartate transcarbamylase (ATCase) est l'archétype de l'enzyme sujette à régulation allostérique (comme la phosphofructokinase 1). La transition allostérique T <===> R correspond à :
L'ATCase est aussi l'une des enzymes les plus étudiées sur le plan mécanistique :
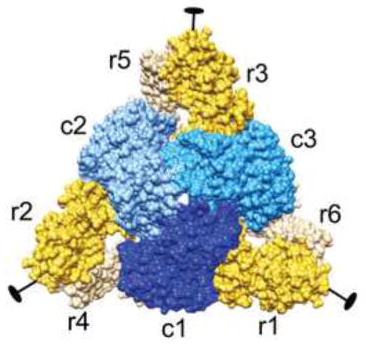
Nomenclature : carbamyl aspartate ou N-carbamyl-L-asparte ou acide uréido-succinique. Chez Escherichia coli, l'ATCase est composée de 6 sous-unités catalytiques (C) et 6 sous-unités régulatrices (R) :
Source : Kantrowitz (2012)
On recense plusieurs dizaines de structures cristallographiques de l'ATCase dans la PDB. Réaction 2. La carbamoyl aspartate déshydratase contient 3 domaines :
Réaction 3. La dihydro-orotate déshydrogénase :
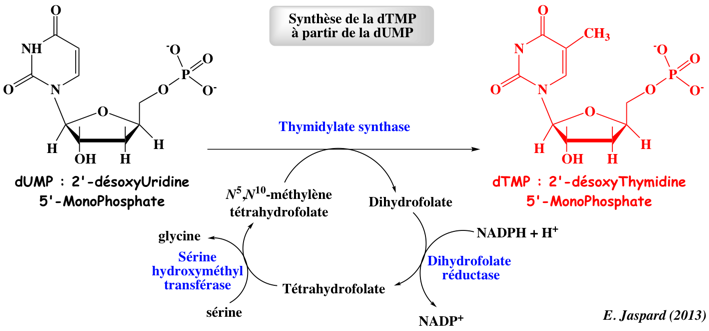
Réaction 4. L'orotate phosphoribosyl transférase (E.C. 2.4.2.10) : chez la levure et les bactéries c'est une enzyme indépendante. Chez les mammifères et les organismes multicellulaires, elle correspond à un domaine d'une enzyme bi-fonctionnelle, l'uridine monophosphate synthetase (UMP synthase). Réaction 5. L'orotidine-5'-phosphate décarboxylase (E.C. 4.1.1.23). Les nucléotides de thymine sont à leur tour obtenus :
|
8. Régulation de la synthèse des nucléotides de pyrimidines Régulation allostérique de la carbamoyl-phosphate synthase I (tous les sites de fixations des effecteurs sont situés dans la grande sous-unité) :
Régulation allostérique de l'aspartate transcarbamylase (ATCase) :
Carbamoyl-phosphate synthase glutamine-hydrolysing (CPSase II) :
9. Catabolisme des nucléotides de pyrimidines Le catabolisme des nucléotides de pyrimidine aboutit finalement à la β-alanine (catabolisme de CMP et UMP) ou au β-amino-isobutyrate (catabolisme de dTMP) et à NH3 et CO2. La β-alanine et le β-amino-isobutyrate sont des donneurs de groupement -NH2 dans les réactions de transamination de l'α-cétoglutarate en glutamate. Une réaction subséquente convertit les produits en malonyl-CoA (qui peuvent être redirigés vers la synthèse des acides gras) ou en méthylmalonyl-CoA qui est converti en succinyl-CoA qui rejoint le cycle de Krebs. |
| 10. Liens Internet et références bibliographiques |
|
"Section 25.2 : Purine Bases Can Be Synthesized de Novo or Recycled by Salvage Pathways" "Section 25.4 : Key Steps in Nucleotide Biosynthesis Are Regulated by Feedback Inhibition" Purine Metabolism - KEGG Pathway Cours : "Nucleotide Metabolism" - The Medical Biochemistry Page |
|
|
Purinosome An et al. (2008) "Reversible Compartmentalization of de Novo Purine Biosynthetic Complexes in Living Cells" Science 320, 103-106 Zhao et al. (2013) "The purinosome, a multi-protein complex involved in the de novo biosynthesis of purines in humans" Chem. Commun. 49, 4444-4452 |
|
| Kantrowitz (2012) "Allostery and cooperativity in Escherichia coli Aspartate Transcarbamoylase" Arch. Biochem. Biophys. 519, 81 - 90 | Article |
![]()