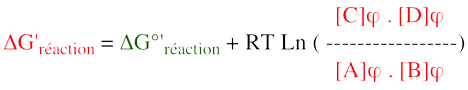| Variation d'Energie Libre de Gibbs - conditions standard (ΔG'°) et physiologiques (ΔG') Constante d'équilibre - ATP et couplage énergétique de réactions |
| Tweet |
|
| I. Introduction au métabolisme et à la bioénergétique
II. Rappels des principes de la thermodynamique et de fonctions d'état III. Variation d'énergie libre de Gibbs et constante d'équilibre d'une réaction |
IV. L'ATP : principale source d'énergie dans la cellule 1. Couplage des réactions biochimiques 2. Structure et hydrolyse de l'ATP 3. Couplage de l'hydrolyse de l'ATP et de la biosynthèse de molécules par transfert de groupement phosphoryle 4. L'énergie d'hydrolyse de certains métabolites est couplée à la synthèse de l'ATP 5. Le coenzyme A : un autre composé à haut potentiel énergétique impliqué dans le transfert de groupe acyle 6. Les enzymes et l'abaissement de l'énergie d'activation |
| I. Introduction au métabolisme et à la bioénergétique
1. Pour vivre les organismes doivent extraire de l'énergie à partir de la matière environnante et la convertir en d'autres formes d'énergie propres à leur existence. Les organismes peuvent être divisés en deux classes étroitement liées :
2. Toute cellule est le siège de milliers de réactions biochimiques qui mettent en jeu des transferts de matière et d'énergie. Cet ensemble de réactions s'appelle le métabolisme. Les réactions forment un réseau de voies très ramifiées le long desquelles les molécules, que l'on appelle des métabolites, sont transformées.
L'ensemble de ces réactions se déroulent à une très grande vitesse, bien supérieure à celles qu'elles auraient isolément dans la nature, grâce à des catalyseurs biologiques que sont les enzymes. Voir un cours sur la métabolomique. 3. En conclusion, la cellule va constamment :
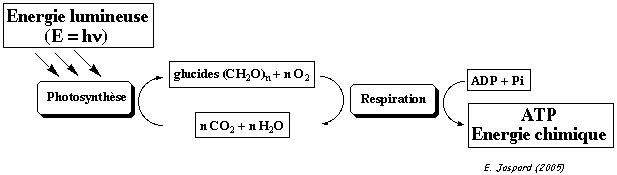
Tous les êtres de la biosphère dépendent de la transformation de l'énergie lumineuse en énergie chimique. Exemples de travaux cellulaires :
Le travail chimique est développé dans ce cours. |
|
II. Rappels des principes de thermodynamique et de fonctions d'état On ne peut expliquer les principes qui régissent les mouvements de matière et d'énergie entre les organismes et leur environnement et au sein des organismes eux-mêmes qu'en faisant appel aux lois de la thermodynamique. L'application des lois de la thermodynamique aux réactions biochimiques constitue la bioénergétique. Rappelons tout d'abord les deux formes principales d'énergie :
|
|
3. Second principe : Entropie (S) Selon le second principe de la thermodynamique, tout échange ou transformation d'énergie dans un système ouvert augmente son entropie, c'est-à-dire le degré de désordre de ce système. On peut aussi définir l'entropie de la manière suivante : tout processus spontané s'accompagne d'une augmentation de l'entropie de l'Univers, c'est-à-dire de l'entropie du système et du milieu extérieur. Cette augmentation de l'entropie vient du fait qu'au cours de la plupart des transformations énergétiques, une partie de l'énergie est convertie en chaleur qui est perdue en se dispersant dans le milieu extérieur, ce qui augmente son degré de désordre. De ce fait, en accomplissant leurs fonctions, les cellules convertissent inévitablement une partie de l'énergie en chaleur, et le meilleur exemple que l'on puisse donner de ce phénomène est la chaleur vite étouffante qui règne dans un amphithéâtre bondé. Le haut degré d'organisation des organismes biologiques contredit-il le second principe de la thermodynamique ? NON : car bien que la variation nette de l'entropie de tout processus soit positive, l'entropie d'un système donné (comme une cellule par exemple) peut décroître à condition que l'accroissement de l'ordre de ce système soit au moins compensé par l'accroissement du désordre (donc de l'entropie) du milieu extérieur. En effet, selon le second principe, c'est l'entropie de l'Univers, donc l'entropie totale du système et du milieu extérieur qui doit s'accroître. Par exemple, le repliement d'une chaîne polypeptidique ou la formation d'une bicouche lipidique des membranes aboutissent à des états dont l'entropie est plus faible (ces 2 processus sont mûs essentiellement par une énergie qui émane des interactions de caractère hydrophobe). En fait, cette diminution d'entropie est contrebalancée par une augmentation importante de celle des molécules d'eau environnantes. On peut donc conclure que :
Illustrations biologiques de l'augmentation de l'entropie :
|
|
L'entropie n'est pas une fonction directement utilisable puisqu'il est impossible de mesurer l'entropie de l'Univers. Le physicien américain, Josiah Willard Gibbs, a introduit à la fin du siècle dernier une nouvelle fonction d'état que l'on appelle (en son hommage) l'énergie libre de Gibbs ou enthalpie libre ou G. Cette fonction s'écrit à une température donnée : ΔG = ΔH - TΔS
En conséquence, la capacité d'un système à fournir un travail utile diminue à mesure que ce système se rapproche de son état d'équilibre, puisque son énergie potentielle (son enthalpie) est de plus en plus faible. A l'équilibre, le système ne peut plus fournir de travail : ΔG est NUL.
Entropie d'un système et degré de désordre qui en découle : exercice d'illustration N°2 |
|
Voir un développement expliquant : (i) la constante d'équilibre; (ii) les conditions d'état standard et ΔG°'; (iii) les conditions de concentrations physiologiques et ΔG'. |
|
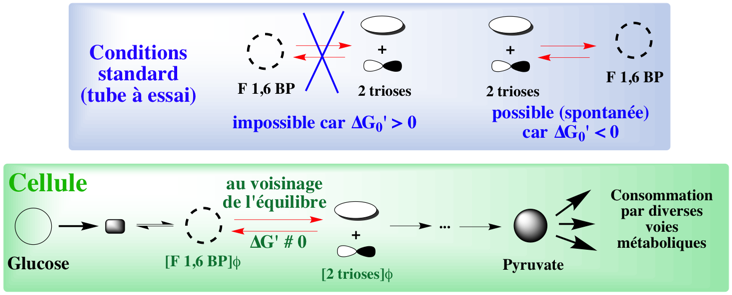
2. Processus à l'état stationnaire ou à l'état d'équilibre Une notion extrêmement importante est celle d'état stationnaire des voies métaboliques. Chaque voie métabolique correspond à un grand ensemble de réactions biochimiques dont chacune a pour substrat le produit de la réaction qui la précède. Considérons le cas le plus simple, un enchaînement de trois réactions : A -> B -> C. La concentration de l'intermédiaire métabolique B est dans un état dit stationnaire quand la vitesse de formation de B à partir de A est égale à la vitesse de disparition de B pour former C. A l'état stationnaire, la concentration de B est donc constante, alors que A disparaît et que C s'accumule : il y a un apport constant du premier métabolite de cette voie et une consommation constante du dernier métabolite. Cette situation est donc distincte d'une réaction à l'équilibre (A <=> B), pour laquelle les concentrations des 2 métabolites sont constantes. En conséquences, les voies métaboliques sont dans un état stationnaire qui n'est pas forcément un état d'équilibre. En d'autres termes, les réactions d'une voie métabolique sont plus ou moins éloignées de l'état d'équilibre. Quelle en est la cause thermodynamique ? C'est l'augmentation de l'entropie d'un système qui le pousse vers un état d'équilibre, et cet état d'équilibre s'accompagne d'une augmentation du désordre de ce système. Mais dans les systèmes métaboliques, les forces de l'entropie sont dominées par des processus cellulaires qui sont créateurs d'ordre, c'est-à-dire par des processus qui diminuent l'entropie, donc qui ont tendance à éloigner le système de l'état d'équilibre. Cette balance entre rapprochement et éloignement de l'état d'équilibre est l'état stationnaire. L'état stationnaire est une notion capitale dans un domaine en voie de développement : la reconstruction métabolique à l'échelle d'un génome. |
|
3. Irréversibilité d'une réaction et régulation des voies métaboliques De la relation :
il découle un concept primordial : la concentration physiologique des métabolites régit la variation d'énergie libre de Gibbs d'une réaction ΔG', donc la spontanéité de cette réaction et le sens dans lequel elle se déroule.
Réactions réversibles - flèches doubles :
Réactions irréversibles - une flèche dans le sens substrat(s) -> produit(s) :
Cette dernière caractéristique explique aussi pourquoi les réactions réversibles ne peuvent pas constituer des points de contrôle du métabolisme : l'activité des enzymes qui catalysent les réactions réversibles n'est pas modulable par des effecteurs. |
|
2. Structure et hydrolyse de l'ATP L'ATP est un triphosphate de nucléoside dans lequel :
|
|
L'énergie libre de Gibbs contenue dans l'ATP est transférée au moment où les liaisons phosphoanhydride sont scindées pour former :
Bien que les groupes phosphoryle de l'ATP soient généralement transférés à d'autres accepteurs que l'eau, les réactions d'hydrolyse donnent des valeurs utiles pour évaluer les variations d'énergie libre qui découlent de ces coupures. Dans les conditions standard :
Dans les conditions de concentrations physiologiques (dans la cellule), le rapport de la concentration de l'ATP à celles de ses produits d'hydrolyse est très différent du rapport observé quand l'équilibre est atteint dans les conditions standard. Cela se traduit par une valeur plus importante : ΔG' ≈ - 14 kcal.mol-1 |
Explications du caractère fortement exergonique de l'hydrolyse de l'ATP 1er facteur : la répulsion électrostatique Les trois groupements phosphate contigus chargés négativement constituent une disposition instable et riche en énergie potentielle. L'hydrolyse de l'ATP est donc exergonique car, en premier lieu elle diminue la répulsion électrostatique entre les atomes d'oxygène chargés négativement des groupes phosphate de l'ATP.
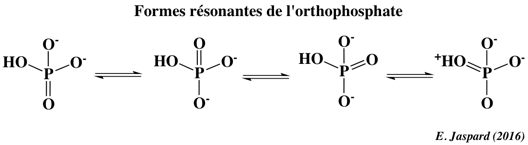
2eme facteur : l'hydratation Les effets de la solvatation et en l'occurrence de l'hydratation des ions contribuent notablement à l'énergie libre négative fournie par l'hydrolyse de l'ATP. En effet, plus un ion est hydraté, plus il s'entoure d'une enveloppe de molécules d'eau qui lui sert d'écran et le protège des effets de répulsion électrostatiques. Or les produits de l'hydrolyse de l'ATP (l'ADP et le Pi ou l'AMP et le PPi) sont plus hydratés que l'ATP, donc mieux protégés et par conséquent plus stables du point de vue thermodynamique, ce qui est en faveur de la réaction dans le sens de l'hydrolyse de l'ATP. 3eme facteur : les structures en résonance L'ion orthophosphate (PO43−) est obtenu quand l'acide orthophosphorique (H3PO4) a perdu ses 3 protons. Il existe plusieurs structures résonantes de l'ion orthophosphate et ces structures ont une énergie similaire (figure ci-dessous).
L'hydrolyse de l'ATP libère un groupe orthophosphate très stable puisqu' il existe plusieurs structures en résonance. En revanche, l'ATP possède moins de structures en résonance car celle qui a un atome d'oxygène chargé positivement adjacent à un atome de phosphore chargé aussi positivement correspond à une configuration défavorable sur le plan électrostatique.
L'ADP, et encore plus le Pi, sont donc davantage stabilisés par résonance que ne l'est l'ATP. En d'autres termes : les produits (ADP et Pi) ont donc un niveau énergétique moins élevé que le réactif (ATP) ce qui favorise la réaction d'hydrolyse de l'ATP. Cette différence de stabilisation liée aux formes en résonance est la principale force qui rend compte du caractère fortement exergonique de l'hydrolyse de l'ATP. |
|
3. Couplage de l'hydrolyse de l'ATP et de la biosynthèse de molécules par transfert de groupement phosphoryle La cellule est capable d'utiliser l'énergie dégagée par l'hydrolyse de l'ATP pour effectuer des processus endergoniques. L'un de ces processus est la biosynthèse de molécules :
Exemple : la synthèse de la glutamine à partir du glutamate par la glutamine synthétase se déroule de la manière suivante :
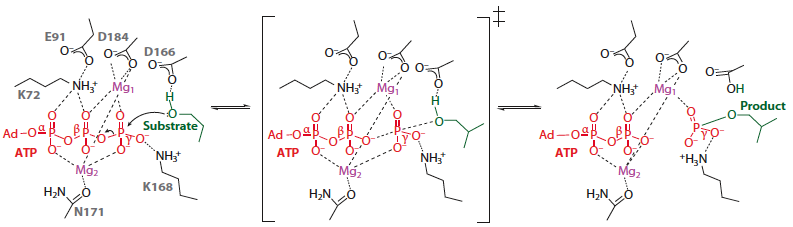
Phosphorylation des protéines Les protéines kinases ou phosphotransférases catalysent le transfert du groupe phosphoryle γ de l'ATP (ou plus rarement d'un autre nucléoside triphosphate) à un autre substrat. Ces enzymes catalysent des réactions irréversibles. Figure ci-dessous : mécanisme catalytique (simplifié) du transfert du groupement phosphoryle de l'ATP par les protéines kinases. L'état de transition est au milieu. Le produit est une tyrosine ou une serine/threonine phosphorylée de la protéine cible. Source : Endicott et al. (2012) |
|
4. L'énergie d'hydrolyse de certains métabolites est couplée à la synthèse de l'ATP Certains métabolites libèrent une quantité d'énergie libre supérieure à celle qui est fournie par l'hydrolyse de l'ATP dans les mêmes conditions. Cette énergie peut donc être utilisée pour la synthèse d'ATP. Les enzymes qui transfèrent le groupe phosphoryle de ces molécules pour former de l'ATP s'appellent aussi des kinases. Il existe 2 molécules dont l'hydrolyse peut être couplée à la synthèse de l'ATP : le phosphoénolpyruvate et la phosphocréatine. 4a. Le phosphoénolpyruvate La liaison phosphate la plus énergétique dans la cellule est celle du phosphoénolpyruvate (PEP), un intermédiaire de la glycolyse : ΔG°' ≈ - 14,8 kcal/mol. Le PEP est un énol ester dont l'hydrolyse se déroule en 3 étapes :
On peut donc rendre compte du fort caractère énergétique du PEP en considérant que cette molécule correspond à un énol bloqué par le groupe phosphoryle : le départ de ce groupe autorise la molécule à prendre la forme cétone qui est beaucoup plus stable. Dans la glycolyse, la pyruvate kinase catalyse le transfert du groupe phosphoryle du PEP à l'ADP pour former de l'ATP et du pyruvate est la pyruvate kinase.
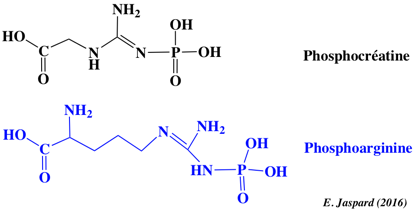
L'autre mode est la synthèse d'ATP par l'ATP synthase à l'issue d'un ensemble de réactions qui ont lieu dans la mitochondrie : la chaîne respiratoire. 4b. La phosphocréatine Le cycle de l'ATP s'effectue à un rythme ahurissant. Une cellule musculaire au travail, par exemple, renouvelle la totalité de son stock d'ATP environ une fois par minute. Cela représente 10 millions de molécules d'ATP utilisées et régénérées par seconde et par cellule. Au début d'un effort musculaire, dans les muscles des vertébrés, la créatine kinase qui catalyse l'approvisionnement en ATP en transférant le groupe phosphoryle activé de la phosphocréatine. La phosphocréatine et la phosphoarginine sont des phosphagènes. Leurs liaisons sont phosphoamides et non pas phosphoanhydrides.
5. Le coenzyme A : composé à haute énergie libre impliqué dans le transfert de groupe acyle Le coenzyme A ou CoA ou CoASH est la molécule qui permet les réactions de transfert des groupes acyles (R-C=O), par exemple lors du catabolisme des acides gras.
|
|
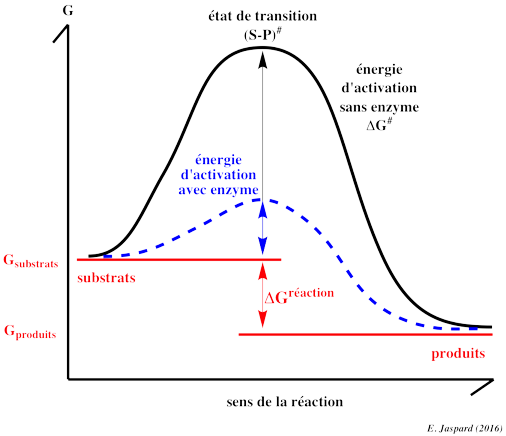
6. Les enzymes et l'abaissement de l'énergie d'activation Les enzymes sont les catalyseurs des réactions biochimiques. Ce sont des protéines et chacune d'entre elle est spécifique d'une réaction donnée. Les molécules qui réagissent avec les enzymes sont appelées les substrats ou réactifs et les molécules obtenues après la réaction sont les produits. Un substrat et un produit sont caractérisés par des liaisons chimiques. Au cours d'une réaction, des échanges d'énergie avec le milieu environnant ont lieu : certaines liaisons du substrat sont rompues en absorbant de l'énergie et certaines liaisons du produit sont formées en libérant de l'énergie. Dans le cas d'une réaction exergonique, l'énergie nécessaire pour rompre les liaisons est inférieure à l'énergie libérée lors de la formation des liaisons. L'énergie requise pour que la réaction ait lieu s'appelle l'énergie libre d'activation ou énergie d'activation : ΔG#activation. C'est l'énergie nécessaire pour que les liaisons du substrat qui doivent être rompues le soient.
Les enzymes augmentent la vitesse de la réaction en abaissant l'énergie d'activation ΔG# : à la même température, les réactifs franchissent plus facilement et donc plus fréquemment la barrière d'activation.
Les enzymes ne modifient pas la variation d'énergie libre de Gibbs de la réaction : ΔGréaction = Gproduits - Gsubstrats. Il faut noter que même dans le cas d'une réaction exergonique, les réactifs doivent franchir la barrière d'activation. Cette barrière est essentielle pour la vie : sans elle, les macromolécules (protéines, acides nucléiques ...) à fort potentiel énergétique se décomposeraient spontanément. Même si les lois de la thermodynamique indiquent que cette décomposition est favorisée, peu de molécules peuvent franchir cette barrière à la température caractéristique de la vie cellulaire. Cette particularité permet aux organismes vivants d'en contrôler l'utilisation à l'aide des enzymes. Remarque : certaines molécules d'ARN, les ribozymes, sont capables de catalyser des réactions d'auto-épissage. |
![]()