| La phosphorylation et les protéines kinases |
| Tweet |
|
|
1. La phosphorylation : généralités 2. Mécanisme catalytique des protéines kinases a. Schéma catalytique de la phosphorylation par les protéines kinases 3. Les familles de protéines kinases 4. Les spécificités de substrat et quelques motifs consensus de phosphorylation 5. La sous-unité calalytique des protéines kinases 6. La déphosphorylation par les protéines Ser/Thr ou Tyr phosphatases |
7. Mécanisme d'activation de certaines protéines kinases 8. Les protéines kinases calcium/calmoduline dépendantes - CaM kinases II (animaux) 9. Les protéines kinases calcium dépendantes - CDPK (plantes) 10. La protéine kinase A 11. Généralités sur les MAP-kinases 12. Les MAP kinases p38 13. Modulation de l'activité et de la localisation des facteurs de transcription NFAT 14. Liens Internet et références bibliographiques |
|
1. La phosphorylation : généralités C'est une modification post-traductionnelle capitale des protéines, qui intervient dans un trés grand nombre de processus cellulaires (différenciation, division, prolifération, apoptose, ...) et en particulier dans les mécanismes de signalisation. Le processus de phosphorylation est connu depuis plus d'un siècle (Levene & Alsberg, 1906) suivi de la mise en évidence d'une phosphosérine dans la vitelline (Lipmann & Levene, 1932) puis de la première description de la phosphorylation d'une protéine (Burnett & Kennedy, 1954). Dans les années 50, Edwin Gerhard Krebs et Edmond Fischer ont déterminé le mécanisme par lequel l'AMP sert d'activateur de la phosphorylase b dans le muscle squelettique. Ils ont constaté que l'ATP est nécessaire pour l'activation de la phosphorylase et que le calcium est un cofacteur important. Ils ont démontré que le phosphate est incorporé dans un résidu sérine spécifique de la phosphorylase b, conduisant à une forme active appelée phosphorylase a. Ils ont confirmé que cette phosphorylation est médiée par une autre enzyme (la kinase de la phosphorylase b), elle-même contrôlée par une autre kinase dépendante de l'AMPc (la protéine kinase A). Ces données ont conduit à la notion de cascade de phosphorylation par les kinases. En 1968, Edmond Fischer a purifié la protéine kinase A. E. Krebs et E. Fischer ont reçu le Prix Nobel en 1992. La phosphorylation induit des modifications structurales donc fonctionnelles trés importantes de la protéine cible qui ont pour conséquences (entre autres) :
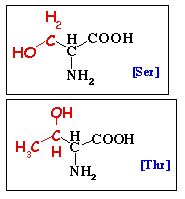
La phosphorylation est une modification qui est réversée par la déphosphorylation catalysée par les protéines phosphatases. La phosphorylation est la réaction d'estérification de la chaîne latérale de la sérine, de la thréonine ou de la tyrosine (chez les Eucaryotes), par addition d'un ou plusieurs groupement(s) phosphate.
Chez les Eucaryotes, le taux de phosphorylation de la sérine, de la thréonine ou de la tyrosine est respectivement de 1000 / 100 / 1.
Bien que quantitativement moins importante, la phosphorylation de la tyrosine a une incidence biologique importante. Par exemple, l'activité d'un grand nombre de facteur de croissance est contrôlée par la phosphorylation de cet acide aminé. Autres acides aminés phosphorylés
|
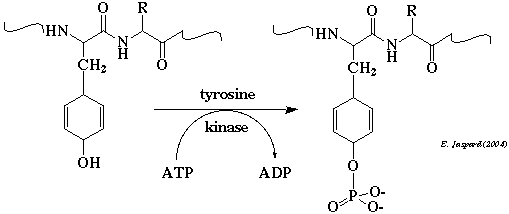
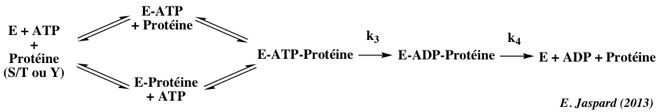
2. Mécanisme catalytique des protéines kinases a. Schéma catalytique de la phosphorylation par les protéines kinases Les protéines kinases catalysent les réactions de phosphorylation. Elles catalysent le transfert du groupe phosphoryle en position γ de l'ATP sur le groupement OH de la sérine, de la thréonine ou de la tyrosine des protéines cibles : protéine (S/T ou Y)-OH + ATP4-.Mg2+ -> protéine (S/T ou Y)-O-PO32- + ADP3-.Mg2+ + H+ Rappel : EC 2 = transférases - EC 2.7 = transfert de groupe contenant un phosphore. Des études cinétiques dans des solvants de viscosités différentes permettant de distinguer les étapes de liaison et de dissociation des étapes catalytiques chimiques ont montré que l'étape catalytique est rapide (k3 ∼ 300 à 500 s-1). La libération des produits est relativement lente (k4 ∼ 20 à 30 s-1) et constituerait l'étape limitante.
Voir le détail du mécanisme catalytique de la protéine kinase FAK-1. Représentation simplifiée du mécanisme catalytique de la phosphorylation par les protéines kinases La réaction comporte 3 étapes principales :
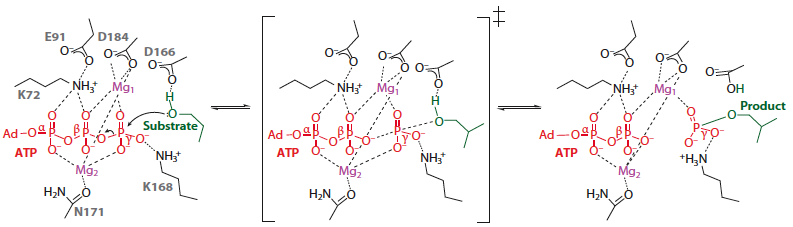
Ci-dessous : la réaction est décrite à partir du complexe [enzyme/substrat], via l'état de transition (au centre) jusqu'aux complexe [enzyme/produit].
Source : Endicott et al. (2012) Le groupe OH de la [Ser/Thr ou Tyr] de la protéine substrat (en vert) est orienté de sorte que la paire d'électrons de l'oxygène est alignée, via le phosphore γ, à l'oxygène [Pβ-O-Pγ] de l'ATP. L'état de transition implique un intermédiaire métaphosphate dans lequel la rupture de la liaison [Pβ-O...Pγ] de l'ATP est bien avancée, tandis que la liaison nucléophile "naissante" liant le groupe phosphoryle γ ne fait que commencer. La charge négative du phosphate γ est compensée par les ions Mg2+ et le groupement aminé d'une lysine (K168). Quand la réaction se déroule, l'acidité du groupement hydroxyle substrat augmente et son pKa devient plus bas que le pKa de l'aspartate catalytique voisin, ce qui permet le transfert d'un proton du groupement hydroxyle (pKa normal ∼12) à l'aspartate (pKa normal ∼ 4.5). Ce proton est probablement transféré par la suite au dianion phosphate du produit, ce qui restaure l'état carboxylate de l'aspartate catalytique. |
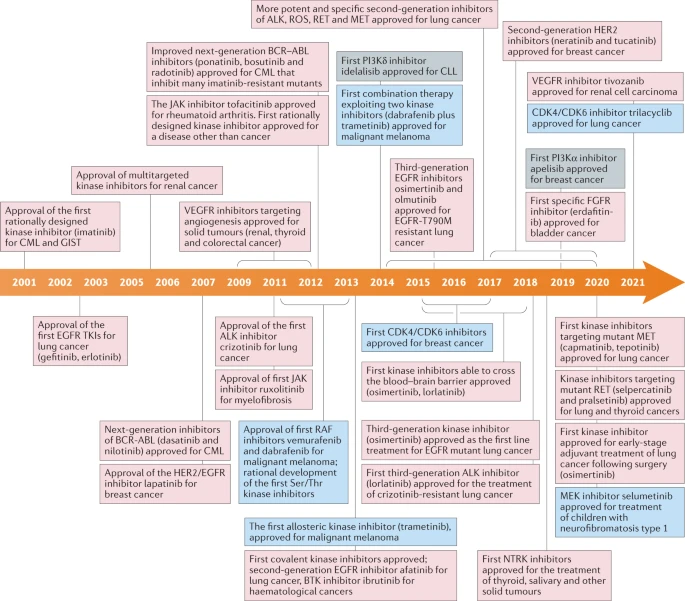
b. Inhibition des protéines kinases et développement de médicaments L'imatinib est le premier inhibiteur de kinases ayant reçu l'approbation par l'agence américaine "US Food and drug Administration" (FDA) en 2001. L'imatinib inhibe la tyrosine kinase d'Abelson (ABL) exprimée sous forme d'une protéine de fusion dérégulée (appelée BCR-ABL) dans presque tous les cas de leucémie myéloïde chronique. Bien que le fasudil (un inhibiteur des protéines kinases RHO-dépendantes) et la rapamycine (sirolimus, un inhibiteur de la protéine kinase TORC1) aient été approuvés avant 2001, ces composés ont été développés et approuvés sans connaître l'identité de leur cible protéique. L'imatinib a donc été le premier médicament développé en ciblant une protéine kinase spécifique. La figure ci-dessous est une chronologie des événements importants dans le développement et l'approbation d'inhibiteurs de kinase depuis 2001.
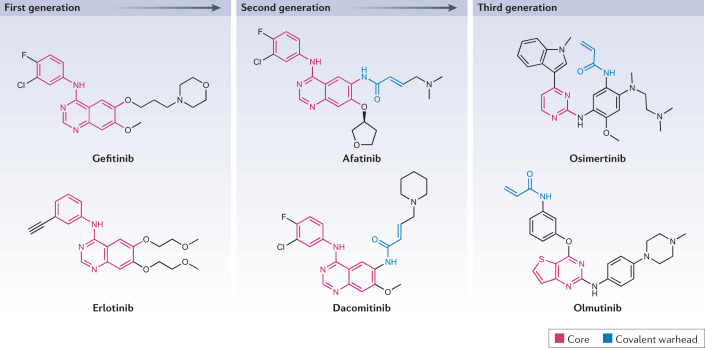
Source : Cohen et al. (2021) Les protéines kinases et les gènes qui les codent sont donc des cibles thérapeutiques majeures pour lutter contre un très grand nombre de pathologies très graves. Les inhibiteurs de la kinase du récepteur du facteur de croissance épidermique ("Epidermal Growth Factor Receptor" - EGFR) illustrent le développement d'inhibiteurs dont la structure de mieux en mieux adaptées améliore leurs propriétés thérapeutiques.
Source : Cohen et al. (2021)
|
|
3. Les familles de protéines kinases Les kinases sont classées en groupes eux-mêmes contenant plusieurs familles. Les 27 familles de protéines kinases constituent l'une des plus vastes et importantes super-familles de protéines.
|
| Il existe de nombreuses bases de données dédiées aux protéines kinases et/ou aux protéines phosphorylées. | |||||
Base de données |
Protéines (en 2016) |
Nombre de sites de phosphorylation | pS | pT | pY |
| dbPAF | 54148 | 483001 | 318016 | 108615 | 56370 |
| Phospho.ELM 9.0 | 10601 | 41070 | 30653 | 7232 | 3185 |
| dbPTM 3.0 | 24601 | 147851 | 102310 | 28797 | 16744 |
| PHOSIDA | 15924 | 64118 | 51318 | 10782 | 2018 |
| PhosphositePlus | 37568 | 310779 | 194217 | 69647 | 46915 |
| PhosphoPep 2.0 | 16234 | 75278 | 57762 | 13492 | 4024 |
| PhosphoGRID 2.0 | 3121 | 19831 | 14871 | 4343 | 617 |
| SysPTM 2.0 | 13867 | 54224 | 41875 | 9126 | 3223 |
| HPRD 9 | 8280 | 51733 | 36052 | 11388 | 4293 |
| UniProt | 14904 | 50713 | 42189 | 6943 | 1581 |
| Source : Ullah et al. (2016) | |||||
| protéines tyrosine kinases | protéines sérine / thréonine kinases (EC 2.7.11.1) |
|
Les protéines tyrosine kinases recencées dans le génome humain forment une trés vaste famille multigènique. Les protéines tyrosine kinases sont divisés en 2 groupes : a. les protéines tyrosine kinases cytoplasmiques (EC 2.7.10.2). b. les protéines tyrosine kinases récepteurs transmembranaires ("transmembrane receptor-linked kinases" / EC 2.7.10.1 - EC 2.7.1.112). Exemple : le récepteur de l'insuline. Elles sont constituées d'un domaine extracellulaire qui fixe un ligand spécifique, d'un domaine transmembranaire et d'un domaine catalytique intracellulaire qui fixe et phosphoryle les protéines cibles. La fixation du ligand au domaine extracellulaire entraîne des modifications structurales des protéines tyrosine kinases, ce qui les activent. |
Quelques exemples :
|
|
protéines histidine kinases (EC 2.7.13.3) Elle n'ont pas de point commun du point de vue structural avec les autres protéines kinases et sont trouvées essentiellement chez les procaryotes et les plantes. Elles fonctionnent en "relai" avec un résidu aspartate qui reçoit le groupement phosphoryle préalablement transféré de l'ATP sur l'histidine. |
|
|
Différences avec Arabidopsis thaliana (plantes) Le génome de Arabidopsis thaliana code pour environ 1100 protéines kinases, soit 4% des 25.500 gènes prédits. Cette proportion est donc le double de celle observée chez l'homme, chez Saccharomyces cerevisiae ou chez Caenorhabditis elegans. Chez Arabidopsis thaliana, les protéines kinases récepteurs transmembranaires phosphorylent les résidus Ser et Thr (elles phosphorylent essentiellement les Tyr chez les animaux - voir ci-dessus). Les deux principaux types de kinases qui médient le signal calcique chez les animaux, les protéines kinases calcium / calmoduline dépendantes (CaM kinase) et la protéine kinase C, semblent absentes ou sous-représentées chez Arabidopsis thaliana. |
|
4. Les spécificités de substrat et quelques motifs consensus de phosphorylation Des analyses bioinformatiques et statistiques des séquences primaires d'un trés grand nombres de substrats de protéines kinases ont montré que les 4 acides aminés (au moins) en amont et en aval du site phospho-accepteur ont une influence sur la sélection du substrat par les protéines kinases. Les caractéristiques physico-chimiques des chaînes latérales de ces 8 acides aminés indiquent que : a. les protéines sérine / thréonine kinases peuvent être schématiquement classées en 3 catégories :
b. les protéines tyrosine kinases sont davantage acidophiles |
| Exemples de motifs consensus de phosphorylation (Schwartz & Gygi, 2005) | ||
| sérine kinases | thréonine kinases | tyrosine kinases |
| RXRXXSP (cas général) | TPP
T180[EPG]Y182 (MAP kinases - double phosphorylation) |
EXXY |
| S/TPX[KR] (kinases cycline dépendantes) | Y[DS] | |
| S/TDX[DE] (caséine kinase II) | YXXP | |
| Bases de données de sites de phosphorylation | Programme de pédiction de sites de phosphorylation (et bases de données) |
| Phospho.ELM : A database of S/T/Y phosphorylation sites | KinasePhos : A web tool to computationally predict phosphorylation sites within given protein sequences |
| PhosPhAt : The Arabidopsis Protein Phosphorylation Site Database | PlantPhos : A web tool for predicting potential phosphorylation sites in plant proteins |
| PhosphoSitePlus : A comprehensive resource for investigating the structure and function of experimentally determined post-translational modifications in man and mouse | NetPhos 2.0 server : Produces neural network predictions for serine, threonine and tyrosine phosphorylation sites in eukaryotic proteins |
| PHOSIDA : Phosphorylation site database | |
| dbPTM : An informative resource for protein post-translational modifications | Analyse du phospho-protéome |
| P3DB : A plant protein phosphorylation database | SWISS-2DPAGE Viewer : recherche interactive dans le phospho-protéome de plusieurs organismes (analyse par spectromètrie de masse) |
|
5. La sous-unité calalytique des protéines kinases La sous-unité catalytique des protéines kinases (qu'elles soit Tyr ou Ser / Thr) est extrêmement conservée du point de vue structural, ce qui permet de mettre au point un grand nombre d'inhibiteurs dans le but de traiter des maladies. La séquence consensus du site catalytique des protéines kinases est : [LIVMFYC]-x-[HY]-x-D-[LIVMFY]-K-x(2)-N-[LIVMFYCT](3) Les protéines kinases sont parmi les enzymes dont on dispose le plus de structures cristallographiques. 510 séquences de protéines kinases de l'homme ont été comparées et fournissent les données suivantes (Source : Kostich et al., 2002) :
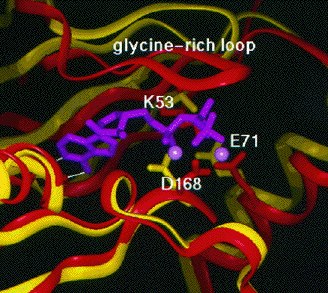
Figure ci-dessus : superposition des sites de fixation de l'ATP et du magnésium (magenta) de la MAP-kinase p38 (jaune) et de la protéine kinase dépendante de l'AMPc (rouge). Source : Wilson et al. (1996)
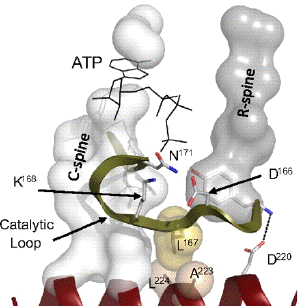
Figure ci-dessus : boucle catalytique et boucle d'activation de la protéine kinase A. Trois résidus catalytiques sont mis en évidence : D166, K168 et N171. Source : Kornev et al. (2008)
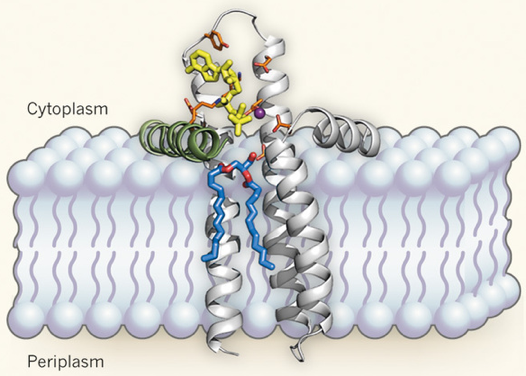
De tous les motifs, c'est le motif riche en Gly qui est le plus variable entre les séquences de protéines kinases. Les autres sont conservés à 95%. Remarque : la numérotation des acides aminés pour chaque sous-domaine mentionné ci-dessus est celle de la sous-unité catalytique de la protéine kinase A. Un cas particulier : le site actif de la diacylglycérol kinase La diacylglycérol kinase bactérienne est une protéine intégrale de la membrane qui catalyse une étape clé dans la synthèse d'oligosaccharides (la conversion du diacylglycérol en acide phosphatidique). Elle forme un homotrimère et elle est fonctionnellement et structuralement distincte des autres kinases. De plus sa petite taille (seulement 121 acides aminés) la rend encore plus intrigante : comment une si petite enzyme peut-elle traverser la membrane tout en structurant un site actif qui doit accommoder un substrat lipidique encombrant et une molécule d'ATP hydrophile ? Les données cristallographiques récentes (Li et al., 2013) sont en faveur de l'hypothèse que chaque monomère "emprunte un composant structural" au monomère voisin et peut ainsi créer un site actif composite. Cet élément emprunté est une hélice amphiphile (hydrophobe et hydrophile) N-terminale. Cette hélice est stratégiquement située à l'interface entre la membrane et le cytoplasme et fournit un résidu catalytique crucial. Figure ci-dessous : schéma de la diacylglycérol kinase dans la membrane interne de Escherichia coli. Bâtonnets oranges : acides aminés catalytiques / sphère pourpre : ion magnésium.
Source : Zheng & Jia (2013) L'hypothèse formulée par Li et al. (2013) est que le site actif qui lie l'ATP (jaune) et le substrat (bleu et rouge), comprend à la fois la membrane et une hélice N-terminale d'un autre monomère (vert). La poche de fixation de l'ATP est dans la partie cytoplasmique de la protéine : elle est formée par les trois hélices transmembranaires d'un monomère et l'hélice N-terminale du monomère voisin. La poche de liaison du substrat est formée en grande partie par la membrane. Ainsi la membrane constituerait une partie du site actif de cette enzyme. |
6. La déphosphorylation par les protéines Ser/Thr ou Tyr phosphatases La phosphorylation est le plus souvent transitoire. Le ou les groupement(s) ajoutés sont ensuite clivés : la déphosphorylation est catalysée par les protéines phosphatases.
La plupart des protéines kinases ont une phosphatase qui leur est associée. La phosphorylation - déphosphorylation est donc un mécanisme impliqué dans la régulation de l'activité biologique de certaines protéines et également un moyen de contrôler finement le flux d'une voie métabolique. Sur la base de leur homologie de séquences en acides aminés, de leurs structures 3D et de leurs sensibilités aux inhibiteurs, les protéines phosphatases sont classées en 4 familles* :
Bien qu'il y ait peu d'homologie entre les séquences en acides aminés des protéines Ser phosphatases et des protéines Thr phosphatases, la structure de leur domaine catalytique est similaire. Elle est en revanche trés différente de la structure des protéines Tyr phosphatases. *On trouve de nombreuses classifications selon les sources qui ne regroupent pas les phosphatases selon les mêmes critères. Figure ci-dessous : comparaison des 3 familles de protéines Ser/Thr phosphatases.
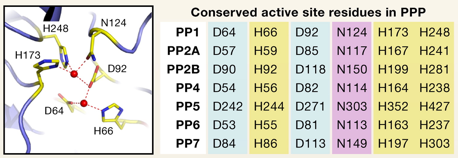
Source : Shi Y. (2009) |
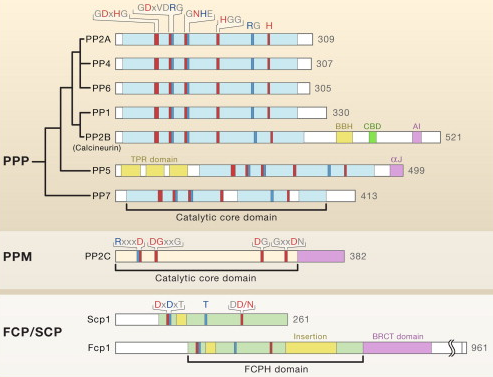
Légende de la figure ci-dessus : toutes les protéines sont d'origine humaine sauf PP7 (Arabidopsis thaliana).
|
|
| BBH : hélice de fixation de CNB | AI : séquence auto-inhibitrice |
| CTD : domaine C-terminal | TPR : "tetratricopeptide repeat" |
| CBD : motif de fixation de la calmoduline | FCPH : domaine homologue à FCP |
Les séquences des motifs signature sont indiquées au dessus des diagrammes. Les domaines catalytiques de chaque protéine sont indiqués en dessous des diagrammes. Les familles PPP et PPM contiennent 3 motifs caractéristiques au sein du domaine catalytique conservé : GDxHG, GDxVDRG, et GNHE. Les acides aminés impliqués dans la coordination au métal et dans la fixation du phosphate sont colorés en rouge et en bleu, respectivement. |
|
|
Différentes phosphatases et leur nomenclature
A l'exception de PP2B et PP2C, elles sont inhibées spécifiquement par les microcystines (famille de toxines peptidiques cyclique des cyanobactéries). Les protéines Ser/Thr phosphatases de type 1 (PP1) Ce sont des phosphatases majeures et ubiquitaires dans toutes les cellules eucaryotes. |
| Chaque PP1 est constituée d'une sous-unité catalytique et d'une sous-unité régulatrice R. | ||
Les séquences des sous-unités catalytiques sont très conservées au sein des eucaryotes (environ 70% ou plus d'identité de séquences). Elles ont une structure 3D conservée. |
Plus de 100 sous-unités régulatrices ont été identifiées. L'analyse des sous-unités R d'un grand nombre d'eucaryotes suggère un accroissement du nombre de sous-unités R concomitant avec l'évolution des organismes multicellulaires. Les sous-unités R pourraient :
|
|
Les associations in vivo des sous-unités catalytiques de PP1 (ou de PP2A) avec différentes sous-unités R génèrent un vaste ensemble de protéines appelées holoenzymes. Ces holoenzymes sont différentiellement exprimées. En conséquence, les interactions entre ces deux types de sous-unités sont capitales pour les trés nombreux processus cellulaires dans lesquels les PP1 sont impliquées :
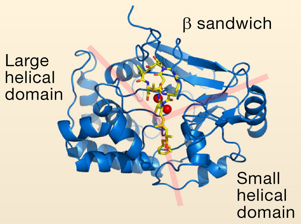
Structure de la sous-unité catalytique de PP1
Source : Shi Y. (2009)
Remarque : la calcineurine ou PP2B met en jeu un ion Fe3+ et un ion Zn2+.
Source : Shi Y. (2009)
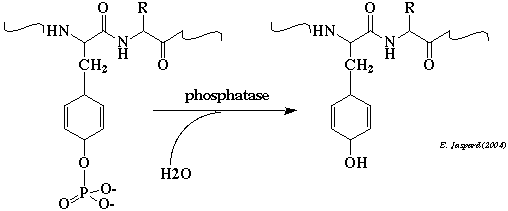
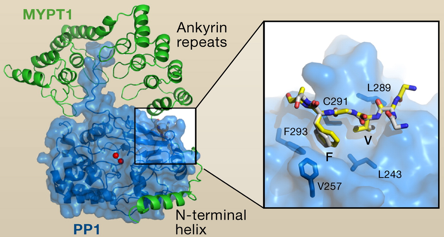
Remarque : les protéines tyrosine phosphatases mettent en jeu une cystéine qui forme un intermédiaire phospho-cystéine avant l'attaque nucléophile de l'atome de phosphore par une molécule d'eau. Figure ci-dessous : structure du domaine catalytique de PP1 (en bleu) lié à la sous-unité régulatrice de la protéine d'adressage de la phosphatase de la myosine (MYPT1 - en vert).
Source : Shi Y. (2009) 3 motifs de MYPT1 sont impliqués dans les interactions :
|
|
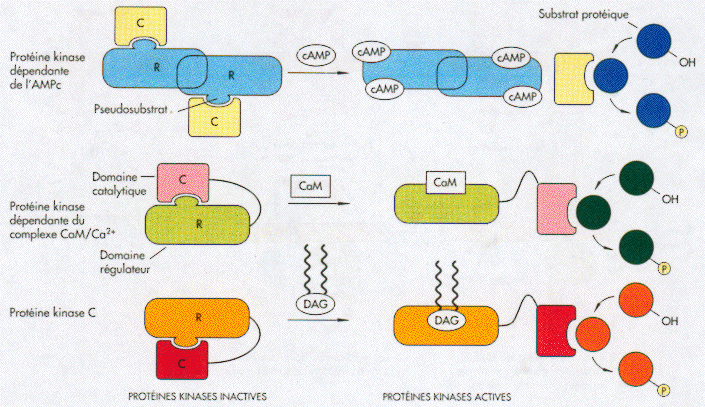
7. Mécanisme d'activation de certaines protéines kinases Chaque protéine kinase dans la figure ci-dessous possède une sous-unité régulatrice (R) qui contient un domaine pseudo-substrat mimant un substrat réel. Ce domaine pseudo-substrat occupe le site catalytique de la sous-unité catalytique (C) et l'inhibe.
Source : "ADN recombinant" Watson et al. (1994) - Ed. DeBoeck
Dans chacun des cas, la fixation des ligands qui activent ces kinases induit un changement de structure qui dissocie le domaine pseudo-substrat du site catalytique. Voir une animation de l'activation des protéines kinases A par l'AMPc. Ces ligands sont respectivement l'AMP cyclique (AMPc), la calmoduline et le diacylglycérol (DAG). Les protéines tyrosine kinases peuvent s'autophosphoryler au niveau du segment d'activation de leur domaine kinase. Ce processus induit un changement de conformation qui les active. Exemples de domaines de fixation dépendants de la phosphorylation : protéines 14-3-3, "Src homology 2" (SH2), "phosphotyrosine binding" (PTB), "BRCA1 C-terminal" (BRCT). |
|
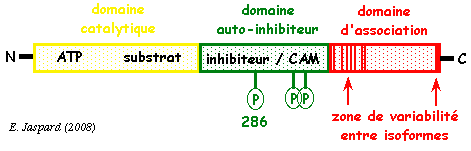
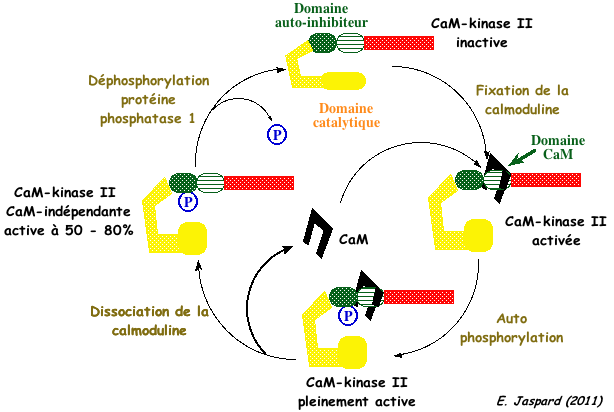
8. Les protéines kinases calcium/calmoduline dépendantes ou CaM kinases II (animaux) Quand la calmoduline a fixé 4 ions calcium, elle adopte une nouvelle conformation qui lui permet d'interagir avec d'autres protéines cibles afin de les activer. En particulier, la kinase II calcium/calmoduline-dépendante ou CaM-kinase II (EC 2.7.1.123) :
La CaM kinase II possède :
|
|
Visualisation de la Cam-kinase II de Homo sapiens à une résolution de 2,3 Å (2010). PDB : 2VZ6
|
a. La fixation de la calmoduline (compléxée au calcium) au domaine de régulation de la CaM-kinase II lève le pouvoir auto-inhibiteur de ce domaine et active le domaine catalytique. b. La CaM-kinase II ainsi activée s'autophosphoryle sur la thréonine 286. c. Une diminution du taux de Ca2+ entraîne une dissociation de la calmoduline de la CaM-kinase II, cette dernière demeurant active. d. L'hydrolyse du groupement phosphate du domaine auto-inhibiteur par la protéine phosphatase 1 inactive de nouveau la CaM-kinase II. e. La protéine kinase A régule l'activité déphosphorylante de la protéine phosphatase 1. L'un des meilleurs motifs consensus de phosphorylation par les CaM-kinases est : [basique-basique-X-basique]-hydrophobe-X(4)-S/T-X-basique (où X est n'importe quel acide aminé). |
|
9. Les protéines kinases calcium dépendantes - CDPK (plantes) Chez les plantes et les protozoaires, les protéines kinases calcium dépendantes (" Calcium-dependent protein kinases" - CDPK - EC 2.7.11.1) médient le signal calcique.
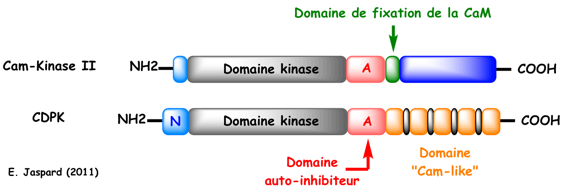
Les CDPK sont des des Ser/Thr kinases. Elles ont une structure hautement conservée. Leur structure est différente de celle des CaM kinases. En effet, les CDPK possèdent (figure ci-dessous) :
En conséquence, les CDPK sont activées directement par le calcium sans l'intervention de calmoduline exogène. Le mécanisme d'activation des CDPK est trés semblable à celui des CaM-kinases. Comme le domaine catalytique des CDPK est hautement homologue de celui des CaM kinases d'animaux (exemple : 44% et 43% d'identité avec, respectivement, la CaM-kinase II α et β de souris), on suppose que les CDPK résultent de la fusion de gènes codant une CaM kinase et une calmoduline. Les différences de séquences en acides aminés entre les isoformes des CDPK sont principalement observées au niveau de l'extrémité trés variable du domaine N-terminal. Ces différences sont aussi liées à l'existence de nombreux sites de modification post-traductionnelle par des acides gras (myristoylation et palmitoylation) qui servent à ancrer les CDPK dans la membrane. Spécifité de substrats des CDPK : voir un tableau des cibles potentielles. L'un des meilleurs motifs consensus de phosphorylation par les CDPK est : [basique-hydrophobe-X-basique]-X(2)-S-X(3)-hydrophobe-basique (où X est n'importe quel acide aminé). |
|
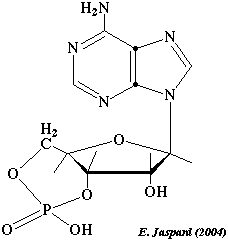
La protéine kinase A (Ser/Thr protéine kinase) est contrôlée par l'AMP cyclique (figure ci-dessous) synthétisé par l'adénylate cyclase.
En son absence, la protéine kinase A est sous forme d'un tétramère constitué de deux types de sous-unités (2 sous-unités catalytiques et 2 sous-unités régulatrices - R2C2) :
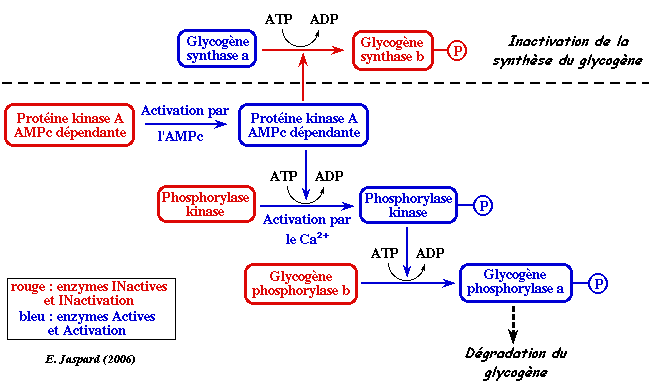
Exemple de cascade de phosphorylation : la glycogènolyse. La protéine kinase A est impliquée dans la régulation de la synthèse du glycogène.
Exemples de protéines phosphorylées : la glycogène synthase et la glycogène phosphorylase des cellules hépatiques en réponse au relarguage du glucagon du pancréas. La phosphorylation :
La protéine kinase A phosphoryle les protéines sur un résidu Ser ou Thr contenu dans un motif consensus : R-X-X-S/T (X est n'importe quel acide aminé). Les sous-unités catalytiques de la protéine kinase A peuvent être régulées par phosphorylation. Mais l'activation de la protéine kinase A est essentiellement contrôlée par un mécanisme de rétro-inhibition :
|
|
11. Généralités sur les MAP-kinases Les voies de signalisation qui impliquent la famille des MAP kinases ("Mitogen-activated protein kinases" - MAPK) transforment des stimuli extracellulaires en réponses intracellulaires par le biais d'un grand nombre de types de récepteurs, tels que :
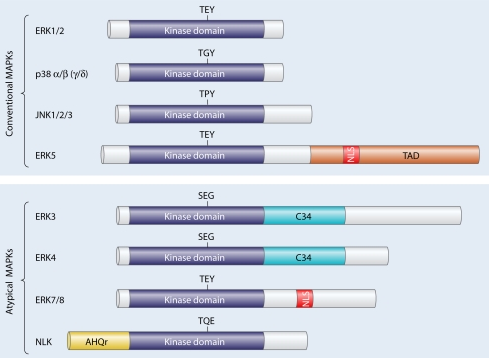
Les MAPK régulent de manière coordonnée la prolifération cellulaire, la différenciation, la mobilité et bien d'autres processus : voir la figure interactive du site Biocarta. Chez les mammifères, on comptabilise actuellement 5 groupes de MAPK :
Source : Cargnello & Roux (2011)
Chaque MAPK possède ses propres substrats, activateurs et inactivateurs :
|
| Exemples de cascade de signalisation de différentes MAPK chez les mammifères | ||||
| Stimulus | Facteur de croissance | Intégrine | Stress oxydatif | IL-1 |
| Activateur | Ras-GTP | Rac1 | Src | TRAF6-TAB1/2 |
MAPK kinase kinase |
c-Raf1 | MEKK1 | MEKK2 | TAK1 |
| MAPK kinase (MAPK - K) |
MKK1 | MKK4 | MKK5 | MKK6 |
| MAPK | ERK1 | JNK1 | ERK5 | p38-α |
| Substrat | Rsk 90 | c-Jun | MEF2 | MNK1 |
| Vue la complexité de la terminologie des MAPK, voici quelques exemples de synonymes | |
| MAP kinase 1 : MAPK1, ERK2 , p42MAPK (gène MAPK1) | MAP kinase 11 : MAPK11, p38 beta, SAPK2B |
| MAP kinase 3 : MAPK3, ERK1, p44MAPK (gène MAPK3) | MAP kinase 12 : MAPK13, p38 gamma, SAPK3 |
| MAP kinase 4 : MAPK4, ERK4, p63MAPK (gène MAPK4) | MAP kinase 13 : MAPK13, p38 delta, SAPK4 |
| MAP kinase 6 : MAPK6, ERK3, p97MAPK (gène MAPK6) | MAP kinase 14 : MAPK14, p38 alpha, SAPK2A |
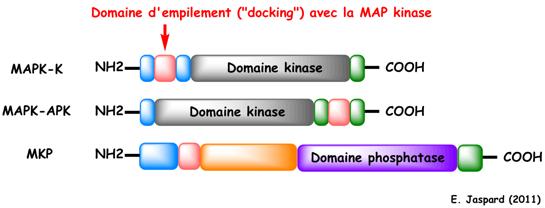
Mécanisme d'activation des MAP-kinases a. Interaction d'empilement ("docking interaction") Les protéines qui interagissent avec les MAPK (leurs protéines substrat, leurs kinases et leurs phosphatases) possèdent des régions appelées domaines ou sites d'empilement ("docking sites" ou "D-sites") qui médient leurs interactions. Les sites d'empilement sont de courts segments d'acides aminés :
Source : Ho et al. (2006)
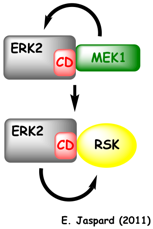
Voir : Whisenant et al. (2010) Figure ci-dessous : exemple d'interactions d'empilement via le domaine d'empilement ("CD domain") d'une MAPK de la famille ERK.
ERK2 : "Extracellular-signal-regulated kinase 2"; MEK1 : [MAPK/ERK kinase 1] ou MAPK-K1. Presque toutes les protéines qui interagissent avec les MAPK (leurs substrats et les MAPK-K) possèdent un motif conservé ("MAPK-docking site" - figure ci-dessous) responsable de cette interaction avec le domaine d'empilement (voir un article sur l'interaction ERK2 / caspase). Le point important est que ce motif se trouve en dehors du domaine catalytique (kinase ou phosphatase). L'interaction protéine - protéine des MAPK et de ces molécules n'est donc pas semblable à l'interaction enzyme-substrat "classique" observée au niveau d'un site actif.
En haut : position du motif dans le cas des MAPK-K; au milieu : position du motif dans le cas des MAPK-APK; en bas : position du motif dans le cas des MKP. |
b. L'échaffaudage ("scaffolding") Les protéines d'échaffaudage s'associent simultanément aux différents composants de la voie de signalisation des MAPK. |
||
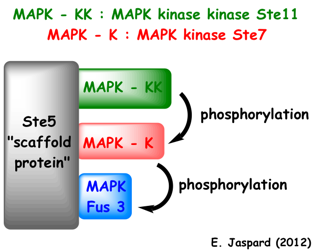
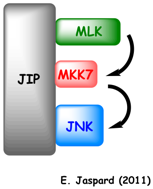
Exemple : chez la levure, la protéine d'échaffaudage ("scaffold protein") Ste5 possède un site de fixation pour la MAPK-KK (Ste11), un site de fixation pour la MAPK-K (Ste7) et un site de fixation pour la MAPK (Fus3).
La MAPK-KK active la MAPK-K qui à son tour active la MAPK. De plus, Ste5 change la conformation de la MAPK Fus3, ce qui permet qu'elle soit phosphorylée et activée par la MAPK-K Ste7. |
Autre exemple :
Voir : Dhanasekaran et al. (2007) |
|
|
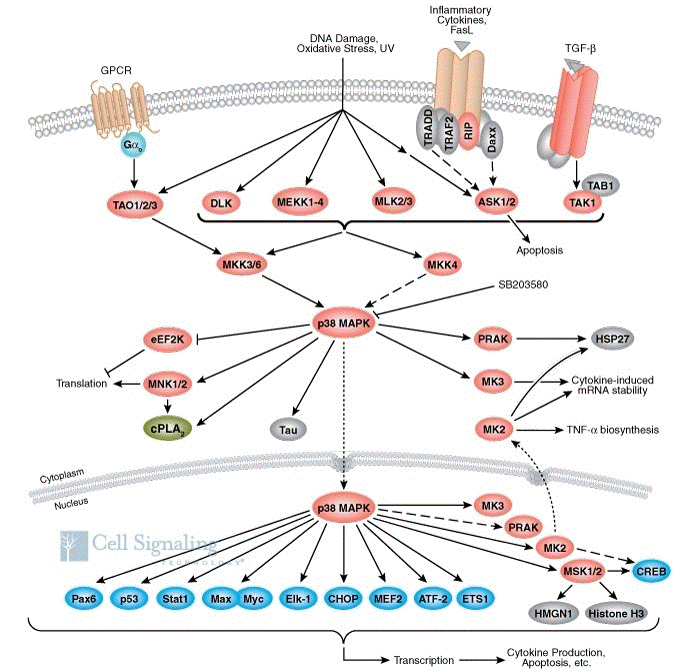
Les MAP kinases p38 ("Mitogen-activated protein kinase p38") des mammifères sont activées par de nombreux stress cellulaires (lipopolysaccharides, choc osmotique, exposition aux ultra-violets) mais aussi en réponse à des cytokines liées à l'inflammation. La sous-famille des MAP kinases p38 contient 4 membres : p38α, p38β, p38γ et p38δ. Elles ont une homologie de séquence d'environ 60%. Cependant, elles différent quant à leurs profils d'expression, leurs spécificités de substrat et leurs sensibilités aux inhibiteurs chimiques tel que le SB203580. L'activité des MAP kinases p38 est capitale pour une réponse immunitaire et inflammatoire normales : en effet, la voie de signalisation des MAP kinases p38 est un élément clé de la régulation de la biosynthèse des cytokines pro-inflammatoires au niveau transcriptionnel et traductionnel. En conséquence, nombre de molécules impliquées dans cette voie sont des cibles thérapeutiques pour la mise au moint de médicaments contre certaines maladies auto-immunes et contre l'inflammation. Des études plus récentes ont montré un rôle plus large encore des MAP kinases p38 puisqu'elles participent au contrôle du cycle cellulaire et du remodelage du cytosquelette. La MAP-kinase p38 (p38 MAPK dans la figure ci-dessous) est activée par phosphorylation de la Thr 180 et de la Tyr 182 par les kinases MKK3 et SEK.
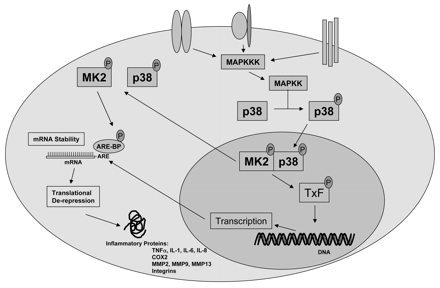
Source : "Cell signaling" La MAP-kinase p38 ainsi modifiée phosphoryle à son tour et active la MAPK-APK 2 (voir ci-après) qui active un certain nombre de facteurs de transcription dont ATF-2. Activation des MAP-kinases par d'autres kinases La voie des MAP-kinase p38 et les processus auxquelles elles sont liés, impliquent le fonctionnement d'une autre famille de kinases dont on découvre de plus en plus de membres : les protéines kinases activées par les MAP-kinases ("(MAP kinases)-activated protein kinases" - MAPK-APK) qui sont des sérine/thréonine kinases. Elles répondent à des stimuli mitogènes ou des stress via une phosphorylation dirigée par une proline ("proline-directed phosphorylation"). Cette activation du domaine kinase des MAPK-APK est catalysée par des kinases 1 and 2 et des MAP kinases p38 qui sont elles-mêmes régulées par des signaux extracellulaires. On compte au moins 11 MAPK-APK chez les vertébrés, réparties en plusieurs sous-familles :
Elles semblent jouer un rôle important dans la traduction des ARN messagers, la prolifération cellulaire et la réponse au niveau du noyau à des agents mitogènes ou des stress cellulaires. Illustration de la régulation de l'expression des gènes de l'inflammation par la MAP-kinase p38 et la MAPK-APK 2 (dénommée MK2 dans la figure ci-dessous). Les ARN messagers qui ont un "turn-over" rapide contiennent souvent des éléments riches en A et U ("AU-rich elements" - ARE) dans la partie 3' non traduite.
Source : Schindler et al. (2007) |
|
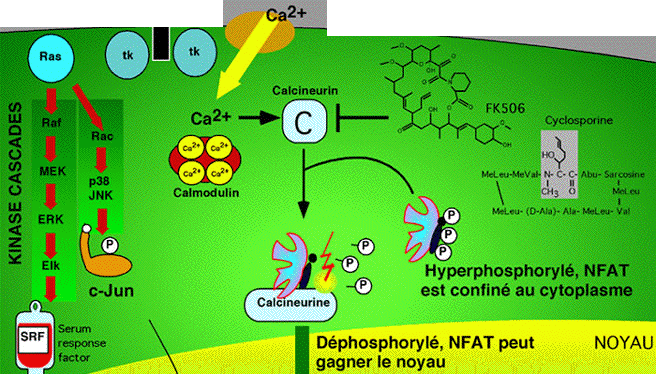
13. Modulation de l'activité et de la localisation des facteurs de transcription NFAT ("Nuclear Factor of Activated T cells") La phosphorylation et la déphosphorylation modulent l'activité de certains facteurs de transcription en les activant directement (exemple : facteur AP-1) et en modifiant leur localisation cellulaire (exemple : facteurs NFkB et NFAT). NFAT est une famille de facteurs de transcription : NFAT1 (NFATc2 ou NFATp); NFAT2 (NFATc1 ou NFATc); NFAT3 (NFATc4); NFAT4 (NFATc3 ou NFATx). Le facteur de transcription NFAT3 est impliqué dans la formation des adipocytes et dans la régulation de l'expression du gène PPAR-γ2.
Source : "Biochimie des protéines" Sa localisation cellulaire dépend de la phosphorylation des sérines en position 168 et 170 par la MAP-kinase p38.
|
| 14. Liens Internet et références bibliographiques |
|
Levene P.A. & Alsberg C.L. (1906) "The cleavage products of vitellin" J. Biol. Chem. 2, 127 - 133 Lipmann F.A. & Levene P.A. (1932) "Serinephosphoric acid obtained on hydrolysis of vitellinic acid" J. Biol. Chem. 98, 109 - 114 Burnett G. & Kennedy E.P (1954) "The enzymatic phosphorylation of proteins" J. Biol. Chem. 211 , 969 - 980 Krebs & Fischer (1956) "The phosphorylase b to a converting enzyme of rabbit skeletal muscle" Biochim. Biophys. Acta 20, 150 - 157 Tanoue T. & Nishida E. (2003) "Molecular recognitions in the MAP kinase cascades" Cell. Signalling 15, 455 - 462 |
|
|
Logiciel de prédiction en ligne : "pkaPS - Prediction of protein kinase A phosphorylation sites using the simplified kinase binding model" Neuberger et al. (2007) "pkaPS: prediction of protein kinase A phosphorylation sites with the simplified kinase-substrate binding model" Biol Direct. 2, 1 "Cell Signaling" : signalisation de voies métaboliques "MPR: Mammalian Phosphorylation Resource" NCI, Bethesda "Phospho.ELM" : a database of experimentally verified phosphorylation sites in eukaryotic proteins. |
|
|
Schwartz, D. & Gygi, S.P. (2005) "An iterative statistical approach to the identification of protein phosphorylation motifs from large-scale data sets" Nature Biotechnol. 23, 1391 - 1398 Kuhn et al. (2007) "Functional Classification of Protein Kinase Binding Sites Using Cavbase" ChemMedChem 2, 1432 - 1447 |
|
|
Kostich et al. (2002) "Human members of the eukaryotic protein kinase family" Genome Biology 3, research0043.1 Wilson et al. (1996) "Crystal Structure of p38 Mitogen-activated Protein Kinase" J. Biol. Chem. 271, 27696 - 27700 Ho et al. (2006) "Interacting JNK-docking Sites in MKK7 Promote Binding and Activation of JNK Mitogen-activated Protein Kinases" J. Biol. Chem. 281, 13169 - 13179 Kornev et al. (2008) "A helix scaffold for the assembly of active protein kinases" PNAS 105, 14377 - 14382 Won et al. (2011) "Recruitment interactions can override catalytic interactions in determining the functional identity of a protein kinase" PNAS 108, 9809 - 9814 |
|
|
Cheng et al. (2002) "Calcium Signaling through Protein Kinases. The Arabidopsis Calcium-Dependent Protein Kinase Gene Family" Plant Physiol. 129, 469 - 485 Dhanasekaran et al. (2007) "Scaffold proteins of MAP-kinase modules" Oncogene 26, 3185 - 3202 Whisenant et al. (2010) "Computational Prediction and Experimental Verification of New MAP Kinase Docking Sites and Substrates Including Gli Transcription Factors" PLoS Comput Biol. 6, e1000908 Cargnello & Roux (2011) "Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases" Microbiol. Mol. Biol. Rev. 75, 50 - 83 |
|
|
Schindler et al. (2007) "p38 Pathway Kinases as Anti-inflammatory Drug Targets" J. Dent. Res. 86, 800 - 811 Shi Y. (2009) "Serine/threonine phosphatases: mechanism through structure" Cell 139, 468 - 484 Endicott et al. (2012) "The Structural Basis for Control of Eukaryotic Protein Kinases" Ann. Rev. Biochem. 81, 587 - 613 |
|
|
Zheng & Jia (2013) "Crystal structure of the integral membrane diacylglycerol kinase" Nature 497, 445 - 446 Li et al. (2013) "Crystal structure of the integral membrane diacylglycerol kinase" Nature 497, 521 - 524 Trentini et al. (2016) "Arginine phosphorylation marks proteins for degradation by a Clp protease" Nature 539, 48–53 |
|
|
Cohen P. (2002) "Protein kinases - the major drug targets of the twenty-first century ?" Nat. Rev. Drug Discov. 1, 309 - 315 Cohen et al. (2021) "Kinase drug discovery 20 years after imatinib: progress and future directions" Nat. Rev. Drug Discov. 20, 551 - 569 |
|
![]()