| Adhésion et signalisation entre cellules : desmosomes, cadhérines, intégrines |
| Tweet |
|
|
1. Contacts et adhésion entre cellules 2. La matrice extracellulaire et les protéoglycanes 3. Les jonctions trouées ou "gap junction" et les plasmodesmes (cellules végétales) 4. Les desmosomes |
5. Les cadhérines 6. Les intégrines, tétraspanines et β-caténines 7. Les sélectines 8. Communication hydrodynamique collective 9. Liens Internet et références bibliographiques |
1. Contacts et adhésion entre cellules Les tissus sont constitués de cellules. Ces cellules se fixent à des protéoglycanes ou des glycoprotéines situés à la surface d'autres cellules ou dans le "substrat" de la matrice extracellulaire (réseau complexe de macromolécules extracellulaires sécrétées) qui les entoure.
Source : Qiagen Ces interactions permettent de définir la forme, la structure et la fonction des tissus. Ces interactions sont donc nécessaires au maintien de l'intégrité structurale des cellules et à la réorganisation des tissus. Les cellules adhèrent entre elles ainsi qu'à la matrice extracellulaire par l'intermédiaire de molécules d'adhésion. Les sites d'adhésion établissent un lien physique entre le milieu extérieur et le cytosquelette d'une cellule. L'adhésion est un processus fondamental pour la vie cellulaire car elle régule la croissance cellulaire, la division, la différenciation, le déplacement des cellules, la prolifération et la mort des cellules. |
Types de contacts et protéines d'adhésion Les contacts entre cellules sont établis via des assemblages adhésifs de quelques micromètres de long. Les adhésions cellulaires se font et se défont en quelques minutes. Il existe 4 grands types de contacts ou jonctions entre cellules :
Un grand nombre de protéines interviennent dans les phénomènes d'adhésion entre cellules : |
| Famille | Ligand | Jonction cellulaire stable |
| sélectines | glucides | non |
| intégrines | matrice extracellulaire | adhésions focale et hémi-desmosomes |
| membres de la superfamille des immunoglobulines | non | |
| superfamille des immunoglobulines | intégrines | non |
| interactions homophiles : interactions qu'établit une molécule d'adhésion à la surface d'une cellule quand elle se fixe au même type de molécule à la surface d'une autre cellule. | non | |
| cadhérines | interactions homophiles | jonctions adhérentes et desmosomes |
| La base de données "UniProt Knowledgebase" recence plus de 1400 protéines impliquées dans l'adhésion. | ||
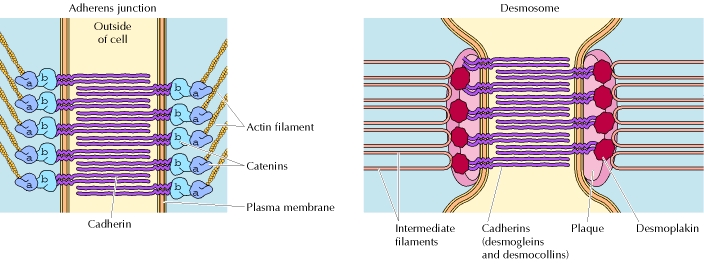
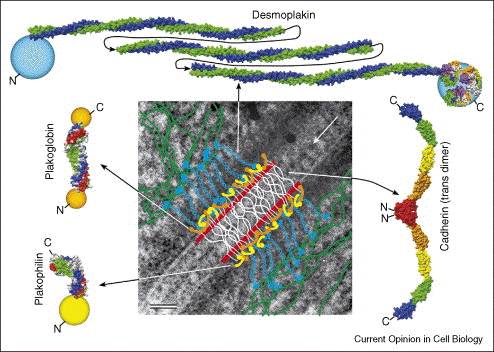
Illustration Les interactions homophiles entre cadhérines médient 2 types d'adhésions stables cellule-cellule (figure ci-dessous) :
Source : NCBI
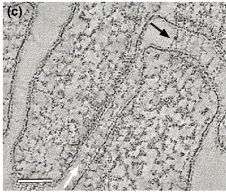
Figure ci-dessous : on distingue des desmosomes (flèche noire) et des jonctions adhérentes (flèche blanche).
Source : Stokes (2007) |
2. La matrice extracellulaire et les protéoglycanes C'est un réseau complexe de macromolécules sécrétées dans le milieu extracellulaire. Elle constitue un espace structuré entre les cellules au sein duquel celles-ci migrent et interagissent. La matrice extracellulaire est composée :
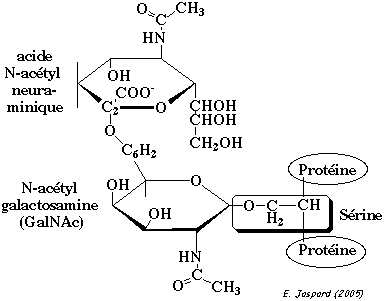
La O-glycosylation est la liaison établie entre :
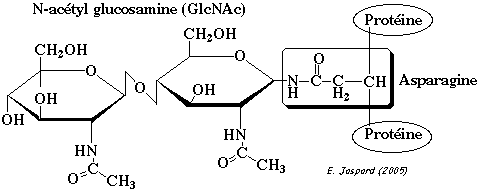
La O-glycosylation est catalysée par des glycoprotéines glycosyltransférases. Les résidus de sucre sont ajoutés directement sur les acides aminés cibles, les uns après les autres et sous forme activée, c'est-à-dire, liés à un nucléotide. La N-glycosylation est un évènement qui se déroule pendant la traduction. Les oses sont fixés à l'azote du groupement amide de l'asparagine. Le sucre immédiatement attachés à l'asparagine est la N-acétylglucosamine.
|
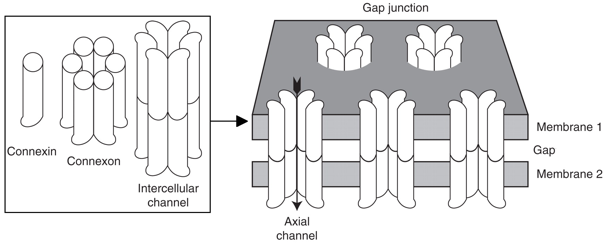
3. Les jonctions trouées ou "gap junction" et les plasmodesmes (cellules végétales) Ce sont des canaux qui permettent la diffusion facilitée de molécules (masse molaire environ < 1KDa) entre les cytoplasmes de cellules ajacentes. Les protéines et les acides nucléiques ne peuvent pas passer. La plupart des cellules des tissus animaux (cellules épithéliales, endothéliales, cardiaques, musculaires, ...) communiquent via des jonctions gap. Dans les cellules électriquement excitables (comme celles du muscle cardiaque), le passage direct d'ions via les jonctions gap couple et synchronise la contraction de cellules adjacentes. Les jonctions gap permettent aussi le passage, entre cellules adjacentes, de molécules de signalisation intracellulaire (comme l'AMPc et le Ca2+) ce qui coordonne la réponse cellulaire dans les tissus. Figure ci-contre : les niveaux multiples de la structure des canaux jonctions gap. Ils sont composés d'hexamères de protéines intrinsèques de taille moyenne : les connexines ou les innexines.
Source : Goodenough & Paul (2009) Les connexines sont synthétisées au niveau du réticulum endoplasmique rugueux puis insérées au réticulum endoplasmique au cours de leur traduction.
Les connexines s'assemblent à l'intérieur de la cellule en hexamères, appelés connexons qui sont dirigés vers la surface de la cellule. Il s'y associent à d'autres connexons de cellules adjacentes : cela forme un canal axial (qui traverse les 2 membranes plasmiques) et un orifice extracellulaire étroit (le "gap"). |
Les plasmodesmes ("plasmodesmata" et "plasmodesma") - cellules végétales L'adhésion entre les cellules de plantes est médiée par leur paroi cellulaire plutôt que via des protéines transmembranaires. En particulier, une région spécialisée riche en pectine de la paroi cellulaire, appelée la lamelle intermédiaire ("middle lamella") agit comme une "colle" pour maintenir accolées les cellules adjacentes. Du fait de la rigidité de la paroi cellulaire des cellules de plantes, les associations stables entre cellules ne requièrent pas l'établissement de liens physiques avec le cytosquelette, tels que ceux qui se forment via les jonctions adhérentes et les desmosomes dans les cellules animales. Cependant, les cellules adjacentes de plantes communiquent via des connections cytoplasmiques appelées plasmodesmes. Elles fonctionnent de manière analogues aux jonctions gap des cellules animales, bien qu'elles n'aient pas d'analogie structurale. Au niveau du plasmodesme, les membranes plasmiques de cellules adjacentes sont continues, formant ainsi des canaux cytoplasmiques au travers de ces cellules. Une extension du réticulum endoplasmique passe au travers de ce canal. Les plasmodesmes jouent un rôle important dans le développement de la plante en contrôlant le traffic de molécules impliquées dans des mécanismes de régulation telles que les facteurs de transcription ou les ARN. |
|
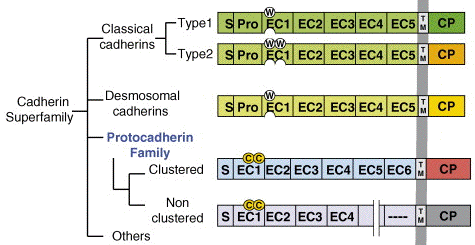
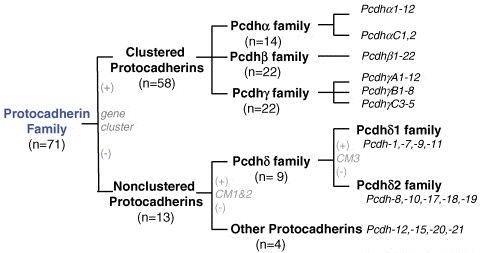
Les cadhérines sont des protéines transmembranaires impliquées dans l'adhésion entre cellules. On trouve les cadhérines depuis l'amibe jusqu'aux mammifères. Leur fixation est généralement homophilique dans les jonctions adhérentes : une cadhérine d'un type s'associe à une cadhérine du même type d'une cellule adjacente. Les cadhérines constituent une famille multigénique :
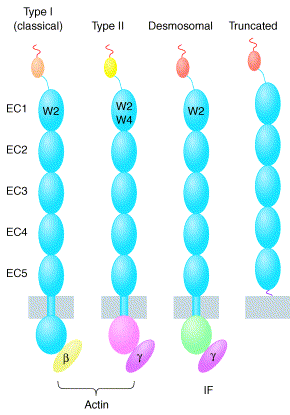
Source : Morishita & Yagi (2007) L'association entre cadhérines s'effectue via une partie de la structure qui leur est commune : leur segment extracellulaire N-terminal qui contient 5 domaines répétés (EC1 à EC5 - 110 acides aminés par domaine). L'association entre cadhérines est dépendante du calcium puisque les interactions entre domaines sont rigidifiées par des sites de fixation du calcium.
Source : Patel et al. (2003) Les cadhérines classiques (type I) ont un tryptophane conservé en position 2. Ce tryptophane est crucial pour l'association entre cadhérines et donc pour l'adhésion entre cellules. (Voir le mécanisme ci-dessous) Les cadhérines de type II ont un plus petit pro-domaine et deux tryptophanes conservés (en position 2 et 4). Les cadhérines de type I et II sont reliées à l'actine du cytosquelette. Les cadhérines desmosomales (desmocollines et desmogleines) sont semblables aux cadhérines de type I mais ont une région cytoplasmique distincte reliée au filament intermédiaire (IF). La cadhérine tronquée (T-cadhérine) est une cadhérine de type I qui ne possède pas de fragment transmembranaire ni le tryptophane conservé en position 2. A l'inverse elle a une ancre GPI. Les E-cadhérines sont des glycoprotéines qui forment, avec les caténines, les jonctions adhérentes dans les cellules épithéliales normales. |
|
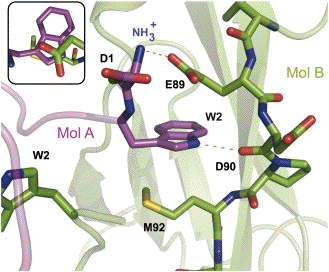
Modèle du mécanisme d'interaction des cadhérines de type I La figure ci-dessous montre les interactions entre l'extrémité N-terminale d'une molécule de cadhérine A (Mol A en magenta) et la poche acceptrice hydrophobe d'une molécule cadhérine B (Mol B en vert) au sein d'un dimère.
Source : Parisini et al. (2007) Les lignes en pointillé indiquent :
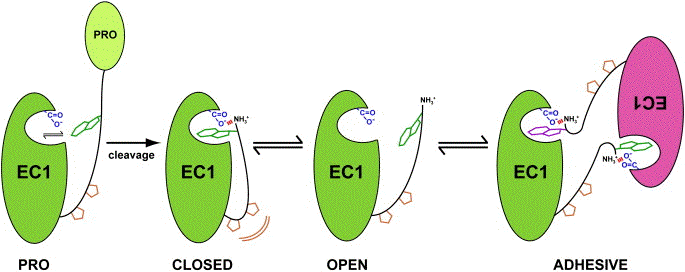
a. les cadhérines sont synthétisées sous forme de précurseur non-adhésif (PRO-cadhérine ci-dessous) puisqu'il n'y a pas encore l'extrémité N-terminale apte à établir le pont salin avec Glu89 (en bleu, figure ci-dessous). b. après protéolyse de la pro-séquence N-terminale (par une protéase de la famille des furines), le "bras" d'adhésion s'ajuste de manière intra-moléculaire dans EC1. L'intercalage du tryptophane en position 2 (en vert) est stabilisée notamment par l'interaction avec Glu89 (E89) et Met92 (M92) et la formation du pont salin (trait rouge) entre Glu89 et le groupement aminé N-terminal. c. cette configuration "fermée" (CLOSED) est en équilibre avec une configuration "ouverte" (OPEN) au sein de laquelle le "bras" adhésif est dissocié de EC1. d. les dimères "adhésifs" sont obtenus par des interactions inter-moléculaires comparables à celles établies dans la forme "fermée".
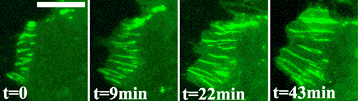
Source : Parisini et al. (2007) On peut localiser la cadhérine et suivre la dynamique des cellules par la technique de couplage à la protéine fluorescente verte (GFP). La figure ci-dessous montre la croissance des contacts (lignes vertes) entre deux cellules : les cadhérines fluorescentes de la cellule de droite montrent l'évolution des zones adhésives.
Source : Brevier et al. (2007) Les cadhérines de la cellule de gauche n'ayant pas été marquées, l'étirement des contacts est uniquement visible sur la cellule de droite bien que le phénomène ait lieu sur les deux cellules. |
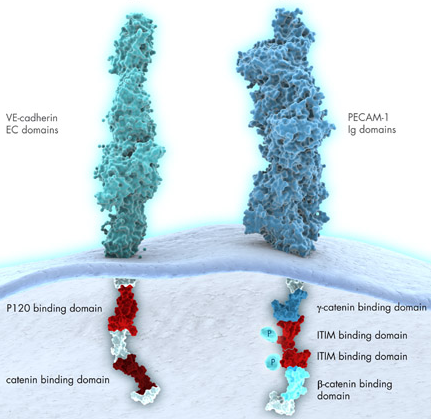
La cadhérine endothéliale vasculaire (CVE) est un élément essentiel des jonctions adhérentes endothéliales et elle joue un rôle capital dans la perméabilité vasculaire. CVE est nécessaire au bon développement vasculaire. Son absence ou une forme tronquée de CVE où il manque le site de liaison de la β-caténine sont létales. Le domaine extracellulaire de CVE est engagé dans une liaison au sein d'une cellule (interaction cis) et dans des liaisons avec des cellules voisines (interaction trans). Comme dans le cas d'autres cadhérines, ce domaine est stabilisé par la fixation d'ions calcium. CVE possède un domaine transmembranaire suivi d'une extrémité cytoplasmique qui s'associe à la β-caténine et à la γ-caténine (plakoglobine). Ces protéines relient CVE au cytosquelette d'actine. La caténine p120 se lie aussi au domaine cytoplasmique de CVE : cette fixation semble réguler la fonction cadhérine, la stabilité et la disponibilité à la surface de la cellule.
Source : Qiagen PECAM-1 ("Platelet Endothelial Cell Adhesion Molecule") est une molécule d'adhésion et de signalisation localisée à la surface des cellules endothéliales, des plaquettes et des leucocytes. C'est une protéine transmembranaire avec une région extracellulaire qui contient six domaines "immunoglobulin-like", un domaine transmembranaire et une courte extrémité cytoplasmique fortement conservée qui contient un motif ITIM immuno-modulateur. Ce motif se compose de deux tyrosines. Une fois phosphorylés, ces tyrosines fixent la "Src homology region 2 domain-containing phosphatase 2" qui inhibe certaine voies d'activation. Dans les cellules endothéliales, PECAM-1 est principalement localisée aux niveau des jonctions intracellulaires et interagit avec d'autres protéines PECAM-1 des cellules voisines. PECAM-1 fixe également d'autres récepteurs comme l'intégrine αvβ3. |
|
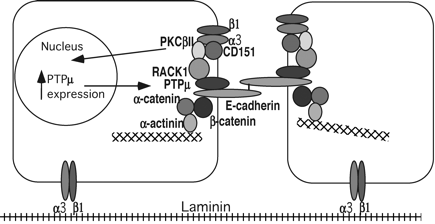
6. Les intégrines, les tétraspanines et les β-caténines Les intégrines forment une large famille de plusieurs dizaines de protéines (notées αiβj avec i de 1 à 7 et j de 1 à 3). La fonction de la plupart des intégrines est de médier l'adhésion des cellules à la matrice extra-cellulaire et l'adhésion entre cellules via les cadhérines. Cependant les intégrines de la famille β2 font exception à cette règle puisqu'elles médient directement l'adhésion des leucocytes aux cellules de l'endothélium. Par ailleurs, certaines intégrines de la famille β1 pourraient aussi médier l'adhésion cellule - cellule quand elles sont complexées aux tétraspanines (CD9, CD53, CD63, ...). [Voir un tableau] Les tétraspanines forment une super-famille de glycoprotéines de la surface cellulaire qui possèdent 4 domaines transmembranaires (TM1 - TM4) et 2 domaines extracellulaires (grande et petite boucles extracellulaires, "large and small extracellular loops (LEL and SEL)" - PDB : 1G8Q ). On leur confère un rôle dans de nombreux processus normaux ou pathologiques (différentiation cellulaire, motilité cellulaire, ...). L'adhésion entre cellules via les cadhérines est renforcée lorsqu'il y a formation de complexes entre les E-cadhérines et les β-caténines (PDB : 1JDH) et que ces complexes s'associent aux molécules d'actine du cytosquelette. La formation de complexes entre les E-cadhérines et les β-caténines est régulée par la phosphorylation d'une tyrosine des β-caténines. Le complexe intégrine - tétraspanine à la surface de la cellule engendre un complexe multimoléculaire [intégrine α3β - CD151 - PKCβII - RACK1 - PTPμ - βcaténine - E-cadhérine].
Source : Chattopadhyay et al. (2003) |
|
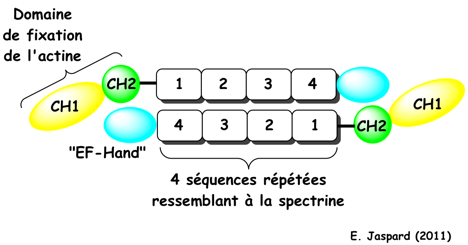
Interactions entre l'α-actinine et les microfilaments d'actine F Dans les celulles du muscle et dans les celulles non - musculaires, l'α-actinine est une protéine d'adhésion qui fixe majoritairement les microfilaments d'actine F et qui relie ces microfilaments directement à des récepteurs d'intégrine. Le domaine de fixation de l'actine de l'α-actinine-3 du muscle humain est formé de deux sous-domaines consécutifs homologues de la calponine. Dans la forme native apo de l'α-actinine-3, ces deux sous-domaines sont dans une conformation dite fermée. Seuls des changements de conformation induisant une certaine orientation de ces deux sous-domaines l'un par rapport à l'autre permettent une fixation efficace aux filaments d'actine. L'aptitude à se fixer à l'actine est régulée par le phosphatidylinositol 4,5-bisphosphate qui se fixe au second sous-domaine homologue de la calponine (Corgan et al., 2004). La structure tridimensionnelle suggère l'existence d'un autre site de fixation du phosphatidylinositol 4,5-bisphosphate (Franzot et al., 2005). Dans ce site, la partie acide gras serait orientée de sorte qu'elle interagirait avec la séquence qui relie le domaine de fixation de l'actine à la première des 4 séquences répétées ressemblant à la spectrine, régulant ainsi la fixation du domaine C-terminal ressemblant à la calmoduline ("EF-hand") à cette séquence de liaison.
|
|
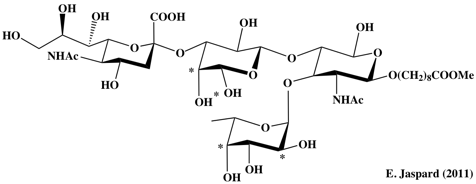
La famille des sélectines (E, P et L) fait partie du groupe des lectines animales de type C qui se fixent à l'épitope sialyl-Lewisx et à d'autres oligosaccharides, notamment l'héparine et le galactosyl - céramide sulfaté. Cette fixation est calcium-dépendante. Structure de l'épitope sialyl-Lewisx : {α}-D-Neu5Ac-[2->3] - β-D-Gal-[1->4]- [{α}-L-Fuc-(1->3)] -β-D-GlcNAc-O-[CH2]8 COO-Me
Code PDB du complexe E-sélectine - épitope : 4KMB La structure des sélectines est constituée :
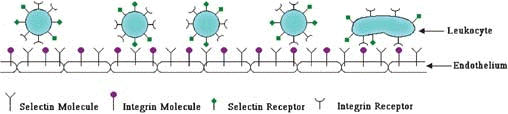
Les P-sélectines sont des protéines d'adhésion multi-domaines à la surface des cellules endothéliales qui interviennent dans la mobilisation des leucocytes au niveau des sites d'inflammation. L'unité de reconnaissance est constituée par le domaine lectine amino-terminal et le domaine EGF.
Les leucocytes "roulent" sur l'endothelium via les sélectines et adhèrent aux intégrines via leur récepteurs respectifs pour finalement atteindre le site d'inflammation |
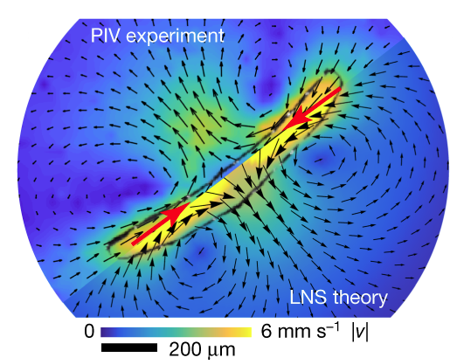
8. Communication hydrodynamique collective Des ondes hydrodynamiques sont déclenchées dans les colonies du protiste unicellulaire Spirostomum ambiguum (longueur 1.1 ± 0.2 mm) et se propagent, de manière similaire à une réaction en chaîne, des centaines de fois plus vite que leur vitesse de nage (0,22 m.s-1 contre 0,2 mm.s-1). En enroulant son cytosquelette, Spirostomum ambiguum peut contracter son corps de 60% en environ 5 ms avec des accélérations pouvant atteindre des forces de 14 g. Une simple contraction cellulaire génère des flux de vortex à longue distance caractérisés par des nombres de Reynolds intermédiaires (environ 50 contre 0,2 au cours de la nage normale) qui peuvent, à leur tour, déclencher des cellules voisines.
Source : Mathijssen et al. (2019) Ce type de signalisation auraient plusieurs conséquences :
|
| 9. Liens Internet et références bibliographiques |
|
"Cell adhesion : proteins involved in the adherence of cells to other cells or to a matrix" : plus de 1400 protéines impliquées dans l'adhésion. |
UniProt Knowledgebase |
|
Stokes (2007) "Protocadherin family : diversity, structure, and function" Curr. Opin. Cell Biol. 19, 565 - 571 Morishita & Yagi (2007) "Protocadherin family : diversity, structure, and function" Curr. Opin. Cell Biol. 19, 584 - 592 Patel et al. (2003) "Cadherin-mediated cell - cell adhesion: sticking together as a family" Curr. Opin. Cell Biol. 13, 690 - 698 Parisini et al. (2007) "The crystal structure of human E-cadherin domains 1 and 2, and comparison with other cadherins in the context of adhesion mechanism" J. Mol. Biol. 373, 401 - 411 |
|
|
Brevier et al. (2007) "Force-Extension Relationship of Cell-Cell Contacts" Physic. Rev. Let. 98, 268101 Chattopadhyay et al. (2003) "alpha3b1 integrin - CD151, a component of the cadherin - catenin complex, regulates PTPµ expression and cell - cell adhesion" J. Cell Biol. 163, 1351 - 1362 Corgan et al. (2004) "Phosphoinositides differentially regulate a-actinin flexibility and function" Biochem. J. 378, 1067 - 1072 Franzot et al. (2005) "The crystal structure of the actin binding domain from alpha-actinin in its closed conformation: structural insight into phospholipid regulation of alpha-actinin" J. Mol. Biol. 348, 151 - 65 |
|
|
Rinnbauer et al. (2003) "Epitope mapping of sialyl Lewisx bound to E-selectin using saturation transfer difference NMR experiments" Glycobiology 13, 435 - 443 Rossier et al. (2012) "Integrins β1 and β3 display distinct dynamic nanoscale organizations inside focal adhesions" Nature Cell Biol. 1057 - 1067 Mathijssen et al. (2019) "Collective intercellular communication through ultra-fast hydrodynamic trigger waves" Nature |
|
![]()