| Les protéases |
| Tweet |
|
|
1. Généralités 2. Principales sources de protéases 3. Classification des protéases a. Classification selon l'Enzyme Commission (EC) 4. Le site de fixation du polypeptide substrat des protéases à sérine 5. Les zymogènes : précurseur des protéases 6. Mécanisme catalytique des protéases à sérine a. Formation d'un acyl-enzyme - "burst" |
c. Le mécanisme "Ping Pong" 7. Les protéases intramembranaires 8. Les protéases intramembranaires des protéines prénylées à motif CAAX a. La protéase Ste24p 9. Les protéasomes 26S, 19S et 20S 10. Les machineries protéolytiques de la famille [AAA+ / ATPase] 11. La protéase ATP-dépendante bactérienne ClpXP 12. Liens Internet et références bibliographiques |
1. Généralités Les protéases sont les enzymes qui hydrolysent la liaison peptidique établie entre 2 acides aminés d'une chaîne polypeptidique (peptides et protéines).
Autres appellations des protéases : peptidases, protéinases ou enzymes protéolytiques. On conçoit la particularité de ces enzymes puisque leurs substrats sont des macromolécules biologiques de même nature. Rappel de certaines caractéristiques des protéines :
Source : Uniprot
Les protéases sont étudiées depuis des décennies :
Les protéases sont classées en 6 groupes : aspartate-, cystéine-, glutamate-, métallo-, sérine- et thréonine-protéases. Elles constituent l'une des plus vastes familles d'enzymes et on estime que 2% des gènes du génome humain codent des protéases (soit environ 550 gènes) :
Caractérisées par une grande diversité structurale et fonctionnelle, les protéases assument une très vaste gamme de fonctions cellulaires qui vont du recyclage des protéines intracellulaires à la digestion des nutriements, en passant par l'amplification de cascades au sein du système immunitaire. Exemples et application :
Les protéases se distinguent notablement par certaines de leurs propriétés :
|
Recherche de médicaments axés sur les protéases On estime que 5 à 10 % de toutes les cibles pharmaceutiques des médicaments en cours de développement sont des protéases.
Le contrôle qualité du bon repliement des protéines Le réseau de contrôle de la qualité des protéines utilise des protéines chaperonnes qui aident des protéines à se replier dans leur conformation native, ou préviennent un mauvais repliement ou "réactivent" des protéines mal repliées et agrégées.
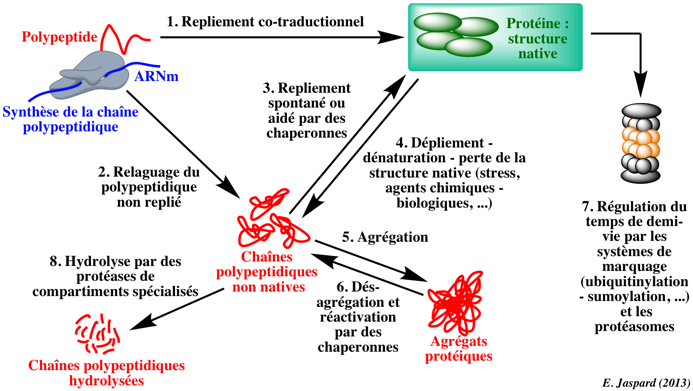
Les chaînes polypeptidiques biosynthétisées se replient dans leur structure native de manière co-traductionnelle avec ou sans aide de protéines chaperonnes (1). Cependant, une fraction des chaînes polypeptidiques peuvent être libérées des ribosomes dans une conformation non native (2). Certaines d'entre elles adoptent alors leur conformation native spontanément ou à l'aide de protéines chaperonnes (3). D'autres n'adoptent pas la conformation native et s'agrègent (5). Des conditions de stress cellulaire (choc thermique, stress oxydatif, certaines pathologies, ...) peuvent induire une dénaturation de protéines correctement repliées (4) qui peuvent également s'agréger et devenir inactives. Ce processus peut être réversé par des protéines chaperonnes (6, 3). Chez les Eucaryotes, les Archées et certaines bactéries, il existe un processus hautement sélectif et régulé qui se déroule dans le noyau et le cytoplasme et qui nécessite de l'énergie (hydrolyse de l'ATP) : il s'agit du système protéolytique ubiquitine-protéasome (7). Des systèmes protéolytiques chez les bactéries ou dans certains compartiments des Eucaryotes (lysosome, peroxysome, mitochondrie, ...) hydrolysent les protéines mal repliées, des protéines membranaires, des protéines extracellulaires et des protéines caractérisées par de trés longs temps de demi-vie (8). Exemple : les protéases Lon (EC = 3.4.21.53 - MEROPS : famille S16 - clan SJ) sont des protéases à sérine ATP-dépendantes qui hydrolysent sélectivement les protéines anormales ou mutées (mal repliées) et certaines protéines régulatrices à demi-vie courte. |
2. Principales sources de protéases Actuellement, le marché mondial des ventes d'enzymes industrielles est supérieur à 3 milliards de dollars et les protéases représentent environ 60 % des ventes totales. Protéases animales : les protéases d'origine animale les plus familières sont la chymotrypsine, la trypsine pancréatique, l'élastase, la pepsine, la rennine.
Protéases de plantes : papaïne, bromélaine, kératinases et ficine sont certaines des protéases les plus connues. Leurs productions à partir de sources végétales est un processus chronophage. D'autant plus que la concentration des protéases dans les tissus est généralement basse.
Protéases microbiennes : la communauté microbienne est privilégiée par rapport aux animaux et aux plantes pour la production de protéases à grande échelle, en raison de leur croissance rapide et de la simplicité de leur cycle de vie pour la génération de nouvelles enzymes recombinantes avec des propriétés modifiées. Les protéases microbiennes représentent les 2/3 de la production commerciale de protéases et jouent un rôle capital dans les détergents, l'industrie pharmaceutique, l'industrie agricole et du cuir. Les protéases alcalines sont le groupe majeur des protéases produites par les bactéries, les champignons, les levures et les actinomycètes. Protéases fongiques : on leur porte un intérêt croissant en raison de la forte diversité et spécificité de substrat et de leur stabilité dans des conditions extrêmes. Elle présentent l'avantage d'être purifiées du mycélium par simple filtration. Les protéases fongiques peuvent être produites par processus de fermentation à l'état solide. Les protéases fongiques sont également utilisés en permettant de modifier les protéines alimentaires. Protéases alcalines bactériennes : elles ont une grande importance commerciale dans les industries de blanchisserie, alimentaires, du cuir et de la soie en raison de leur forte production et de leur forte activité à pH alcalin (pH 8 à pH 12), avec une température optimale entre 50°C et 70°C. Ces propriétés expliquent leur utilisation dans l'industrie des détergents. |
3. Classification des protéases a. Classification selon l'Enzyme Commission (EC) La classification établie par la commission des enzymes de l'Union Internationale de Biochimie et de Biologie Moléculaire (sigle anglais "IUBMB") a été établie sur des critères de spécificité. La nomenclature des enzymes s'écrit de manière générale sous la forme : E.C. X.X.X.X. La liste de toutes les enzymes, leur nomenclature et leur identifiant EC est décrite à : Enzyme Nomenclature and Classification of Enzyme-Catalysed Reactions |
| Le premier "X" correspond aux types de réactions catalysées par les enzymes. | 1er X | Réaction catalysée | exemple de coenzyme impliqué |
| X = 1 : oxydoréductases (E. C. 1.X.X.X) | oxydoréduction | NAD(P)+ | |
| X = 2 : transférases (E. C. 2.X.X.X) | transfert de groupes | phosphate de pyridoxal | |
| X = 3 : hydrolases (E. C. 3.X.X.X) | hydrolyse | aucun | |
| X = 4 : lyases (E. C. 4.X.X.X) | addition de groupe à des atomes engagés dans des doubles liaisons | pyrophosphate de thiamine | |
| X = 5 : isomérases (E. C. 5.X.X.X) | isomérisation (de position de groupe ou de fonction) | phosphate de pyridoxal | |
| X = 6 : ligases (E. C. 6.X.X.X) | condensation de deux molécules | ATP | |
| X = 7 : translocases (E. C. 7.X.X.X) | transfert d'un "côté 1" à un "côté 2" | aucun | |
| Dans les banques de données, on trouve une catégorie "Autres" ; exemples : kinases, phosphoprotéines, "Mutator transposons". | |||
|
Exemples d'hydrolases agissant sur la liaison peptidique Les liens renvoient vers le descriptif détaillé de chaque protéase (fichiers EXPASY) |
|||
| EC 3.4.11 - Aminopeptidases | EC 3.4.13 - Dipeptidases | EC 3.4.14 - Dipeptidyl-peptidases et tripeptidyl-peptidases | EC 3.4.15 - Peptidyl-dipeptidases |
|
3.4.11.1 Leucyl aminopeptidase 3.4.11.2 Membrane alanyl aminopeptidase 3.4.11.3 Cystinyl aminopeptidase 3.4.11.4 Tripeptide aminopeptidase 3.4.11.5 Prolyl aminopeptidase 3.4.11.6 Aminopeptidase B 3.4.11.7 Glutamyl aminopeptidase 3.4.11.9 Xaa-Pro aminopeptidase 3.4.11.10 Bacterial leucyl aminopeptidase 3.4.11.13 Clostridial aminopeptidase 3.4.11.14 Cytosol alanyl aminopeptidase 3.4.11.15 Aminopeptidase Y 3.4.11.16 Xaa-Trp aminopeptidase 3.4.11.17 Tryptophanyl aminopeptidase 3.4.11.18 Methionyl aminopeptidase 3.4.11.19 D-stereospecific aminopeptidase 3.4.11.20 Aminopeptidase Ey 3.4.11.21 Aspartyl aminopeptidase 3.4.11.22 Aminopeptidase I 3.4.11.23 PepB aminopeptidase 3.4.11.24 Aminopeptidase S 3.4.11.25 Beta-peptidyl aminopeptidase 3.4.11.26 Intermediate cleaving peptidase 55 |
3.4.13.4 Xaa-Arg dipeptidase 3.4.13.5 Xaa-methyl-His dipeptidase 3.4.13.7 Glu-Glu dipeptidase 3.4.13.9 Xaa-Pro dipeptidase 3.4.13.12 Met-Xaa dipeptidase 3.4.13.17 Non-stereospecific dipeptidase 3.4.13.18 Cytosol nonspecific dipeptidase 3.4.13.19 Membrane dipeptidase 3.4.13.20 Beta-Ala-His dipeptidase 3.4.13.21 Dipeptidase E 3.4.13.22 D-Ala-D-Ala dipeptidase |
3.4.14.1 Dipeptidyl-peptidase I 3.4.14.2 Dipeptidyl-peptidase II 3.4.14.4 Dipeptidyl-peptidase III 3.4.14.5 Dipeptidyl-peptidase IV 3.4.14.6 Dipeptidyl-dipeptidase 3.4.14.9 Tripeptidyl-peptidase I 3.4.14.10 Tripeptidyl-peptidase II 3.4.14.11 Xaa-Pro dipeptidyl-peptidase 3.4.14.12 Xaa-Xaa-Pro tripeptidyl-peptidase |
3.4.15.1 Peptidyl-dipeptidase A 3.4.15.4 Peptidyl-dipeptidase B 3.4.15.5 Peptidyl-dipeptidase Dcp 3.4.15.6 Cyanophycinase |
| EC 3.4.16 - Serine-type carboxypeptidases | EC 3.4.17 - Métallo-carboxypeptidases | EC 3.4.18 - Cysteine-type carboxypeptidase | EC 3.4.19 - peptidases oméga |
|
3.4.16.2 Lysosomal Pro-Xaa carboxypeptidase. 3.4.16.4 Serine-type D-Ala-D-Ala carboxypeptidase 3.4.16.5 Carboxypeptidase C 3.4.16.6 Carboxypeptidase D |
3.4.17.1 Carboxypeptidase A 3.4.17.2 Carboxypeptidase B 3.4.17.3 Lysine carboxypeptidase 3.4.17.4 Gly-Xaa carboxypeptidase 3.4.17.6 Alanine carboxypeptidase 3.4.17.8 Muramoylpentapeptide carboxypeptidase. 3.4.17.10 Carboxypeptidase E 3.4.17.11 Glutamate carboxypeptidase 3.4.17.12 Carboxypeptidase M 3.4.17.13 Muramoyltetrapeptide carboxypeptidase 3.4.17.14 Zinc D-Ala-D-Ala carboxypeptidase 3.4.17.15 Carboxypeptidase A2 3.4.17.16 Membrane Pro-Xaa carboxypeptidase 3.4.17.17 Tubulinyl-Tyr carboxypeptidase 3.4.17.18 Carboxypeptidase T 3.4.17.19 Carboxypeptidase Taq 3.4.17.20 Carboxypeptidase U 3.4.17.21 Glutamate carboxypeptidase II 3.4.17.22 Metallocarboxypeptidase D 3.4.17.23 Angiotensin-converting enzyme 2 |
3.4.18.1 Cathepsine X |
3.4.19.1 Acylaminoacyl-peptidase 3.4.19.2 Peptidyl-glycinamidase 3.4.19.3 Pyroglutamyl-peptidase I 3.4.19.5 Beta-aspartyl-peptidase 3.4.19.6 Pyroglutamyl-peptidase II 3.4.19.7 N-formylmethionyl-peptidase 3.4.19.9 Gamma-glutamyl hydrolase 3.4.19.11 Gamma-D-glutamyl-meso-diaminopimelate peptidase 3.4.19.12 Ubiquitinyl hydrolase 1 3.4.19.13 Glutathione hydrolase 3.4.19.14 Leukotriene-C(4) hydrolase |
| EC 3.4.21 - endopeptidases à sérine | EC 3.4.22 - endopeptidases à cysteine | EC 3.4.23 - endopeptidases à acide aspartique | EC 3.4.24 - Métallo-endopeptidases |
|
3.4.21.1 Chymotrypsine 3.4.21.3 Metridine 3.4.21.4 Trypsine 3.4.21.5 Thrombine 3.4.21.6 Facteur de coagulation 3.4.21.7 Plasmine 3.4.21.10 Acrosine 3.4.21.12 Alpha-lytic endopeptidase 3.4.21.20 Cathepsine 3.4.21.25 Cucumisine 3.4.21.26 Prolyl oligopeptidase 3.4.21.32 Brachyurine 3.4.21.34 Kallikreine 3.4.21.36 Elastase pancréatique 3.4.21.39 Chymase 3.4.21.45 Complement factor 3.4.21.48 Cerevisine 3.4.21.49 Hypodermine 3.4.21.50 Lysyl endopeptidase 3.4.21.61 Kexine 3.4.21.62 Subtilisine 3.4.21.64 Peptidase K 3.4.21.65 Thermomycoline 3.4.21.69 Protein C 3.4.21.75 Furine 3.4.21.79 Granzyme 3.4.21.80 Streptogrisine 3.4.21.82 Glutamyl endopeptidase II 3.4.21.90 Togavirine 3.4.21.91 Flavivirine 3.4.21.96 Lactocepine 3.4.21.97 Assembline 3.4.21.98 Hepacivirine 3.4.21.99 Spermosine 3.4.21.100 Sedolisine 3.4.21.101 Xanthomonalisine 3.4.21.102 C-terminal processing peptidase 3.4.21.105 Rhomboid protease 3.4.21.106 Hepsine 3.4.21.109 Matriptase 3.4.21.112 Site-1 protease 3.4.21.120 Oviductine |
3.4.22.1 Cathepsine B 3.4.22.2 Papaine 3.4.22.3 Ficaine 3.4.22.6 Chymopapaine 3.4.22.7 Asclepaine 3.4.22.8 Clostripaine 3.4.22.10 Streptopaine 3.4.22.14 Actinidaine 3.4.22.15 Cathepsine L 3.4.22.16 Cathepsine H 3.4.22.24 Cathepsine T 3.4.22.25 Glycyl endopeptidase 3.4.22.27 Cathepsine S 3.4.22.28 Picornaine 3C 3.4.22.30 Caricaine 3.4.22.31 Ananaine 3.4.22.32 Bromelaine 3.4.22.34 Legumaine 3.4.22.35 Histolysaine 3.4.22.36 Caspase 1 3.4.22.37 Gingipain R 3.4.22.39 Adenaine 3.4.22.40 Bleomycin hydrolase 3.4.22.41 Cathepsine F 3.4.22.42 Cathepsine O 3.4.22.43 Cathepsine V 3.4.22.47 Gingipaine K 3.4.22.48 Staphopaine 3.4.22.51 Cruzipaine 3.4.22.52 à 3.4.22.54 Calpaine 1 à Calpaine 3 3.4.22.55 à 3.4.22.64 Caspase 2 à Caspase 11 3.4.22.66 Calicivirine 3.4.22.67 Zingipaine 3.4.22.70 Sortase A |
3.4.23.1 Pepsine A 3.4.23.2 Pepsine B 3.4.23.3 Gastricsine 3.4.23.4 Chymosine 3.4.23.5 Cathepsine D 3.4.23.12 Nepenthesine 3.4.23.15 Renine 3.4.23.16 HIV-1 retropepsine 3.4.23.17 Pro-opiomelanocortine converting enzyme 3.4.23.18 Aspergillopepsine I 3.4.23.20 Penicillopepsine 3.4.23.21 Rhizopuspepsine 3.4.23.22 Endothiapepsine 3.4.23.23 Mucorpepsine 3.4.23.24 Candidapepsine 3.4.23.25 Saccharopepsine 3.4.23.26 Rhodotorulapepsine 3.4.23.28 Acrocylinedropepsine 3.4.23.29 Polyporopepsine 3.4.23.30 Pycnoporopepsine 3.4.23.31 Scytalidopepsine A 3.4.23.32 Scytalidopepsine B 3.4.23.34 Cathepsine E 3.4.23.35 Barrierpepsine 3.4.23.36 Signal peptidase II 3.4.23.38 Plasmepsine 3.4.23.40 Phytepsine 3.4.23.41 Yapsine 3.4.23.42 Thermopsine 3.4.23.43 Prepiline peptidase 3.4.23.44 Nodavirus endopeptidase 3.4.23.45 Memapsine 3.4.23.47 HIV-2 retropepsine 3.4.23.48 Activateur du plasminogen Pla 3.4.23.49 Omptine 3.4.23.52 Preflagelline peptidase |
3.4.24.1 Atrolysine A 3.4.24.3 Collagenase 3.4.24.6 Leucolysine 3.4.24.11 Neprilysine 3.4.24.12 Envelysine 3.4.24.13 IgA-specific metalloendopeptidase 3.4.24.14 Procollagen N-endopeptidase 3.4.24.16 Neurolysine 3.4.24.17 Stromelysine 3.4.24.18 Meprine A 3.4.24.21 Astacine 3.4.24.23 Matrilysine 3.4.24.24 Gelatinease A 3.4.24.25 Vibriolysine 3.4.24.26 Pseudolysine 3.4.24.27 Thermolysine 3.4.24.28 Bacillolysine 3.4.24.29 Aureolysine 3.4.24.30 Coccolysine 3.4.24.31 Mycolysine 3.4.24.32 Beta-lytic metalloendopeptidase 3.4.24.43 Atroxase 3.4.24.44 Atrolysine 3.4.24.46 Adamalysine 3.4.24.47 Horrilysine 3.4.24.48 Ruberlysine 3.4.24.49 Bothropasine 3.4.24.50 Bothrolysine 3.4.24.51 Ophiolysine 3.4.24.54 Mucrolysine 3.4.24.55 Pitrilysine 3.4.24.56 inesulysine 3.4.24.57 O-sialoglycoproteine endopeptidase 3.4.24.58 Russellysine 3.4.24.61 Nardilysine 3.4.24.62 Magnolysine 3.4.24.68 Tentoxilysine 3.4.24.69 Bontoxilysine 3.4.24.71 Endotheline-convertineg enzyme 1 3.4.24.72 Fibrolase 3.4.24.73 Jararhagine 3.4.24.75 Lysostaphine 3.4.24.76 Flavastacine 3.4.24.79 Pappalysine 3.4.24.80 Membrane-type matrix metalloproteinease-1 3.4.24.83 Anthrax lethal factor endopeptidase 3.4.24.84 Ste24 endopeptidase 3.4.24.85 S2P endopeptidase |
| EC 3.4.25 - endopeptidases à thréonine | 3.4.25.1 Complexe du proteasome 3.4.25.2 HslU-HslV peptidase |
||
b. Classification selon les [homologies / similarités] de [séquences / structures] & quelques bases de données Famille : groupe de protéases dans lequel chaque membre possède une relation évolutive avec au moins un autre membre, soit sur l'ensemble de la séquence en acides aminés, soit au moins dans la partie de la séquence liées à l'activité catalytique. Chaque famille est identifié par une lettre majuscule qui représente le type catalytique (S : sérine, T : thréonine ; C : cystéine, A : aspartate, M : métallo, U : inconnu), suivi d'un numéro unique. Clan : groupe de familles considérées comme ayant des ancêtres communs. Les membres sont le plus souvent regroupés par similitudes de structure tertiaire, mais l'ordre des résidus catalytiques dans la séquence peut être utilisé. Chaque clan est identifié par 2 lettres majuscules : la première lettre spécifie le type de catalyse (comme pour les familles) / la deuxième lettre est propre au clan.
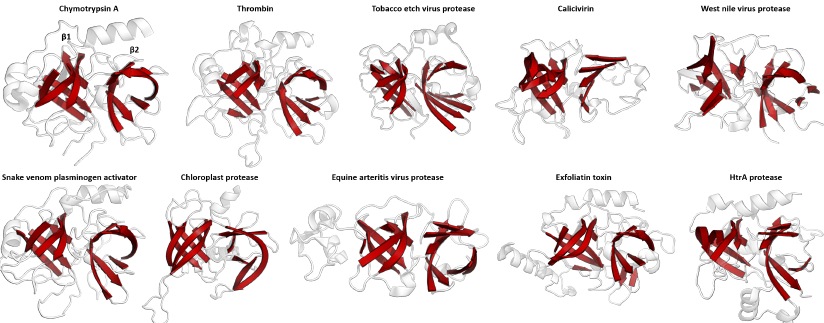
Source : Wikipedia Figure ci-dessus : homologie structurale au sein de la superfamille des protéases du clan PA (clan PA : "Proteases of mixed nucleophile, superfamily A"). Le double tonneau β qui caractérise cette superfamille est en rouge. Exemples : chymotrypsine (PDB 1GG6), thrombine (1MKX), protéase du chloroplaste (4FLN). Le clan PA contient les protéases à sérine et les protéases à cystéine. On trouve des protéases du clan PA chez les plantes, les animaux, les champignons, les eubactéries, les archées et les virus. Le clan PA est un exemple :
La base de données MEROPS classe les protéases sur la base de leur similarité structurale :
Base de données PFAM - Exemples de familles de protéases : |
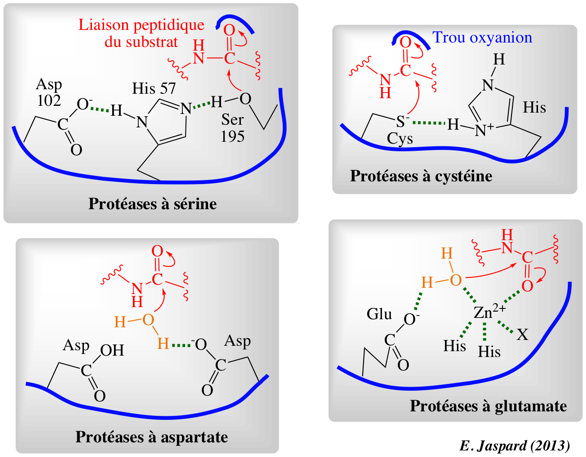
c. Classification des protéases du point du mécanisme catalytique Elles se répartissent en 4 grandes classes :
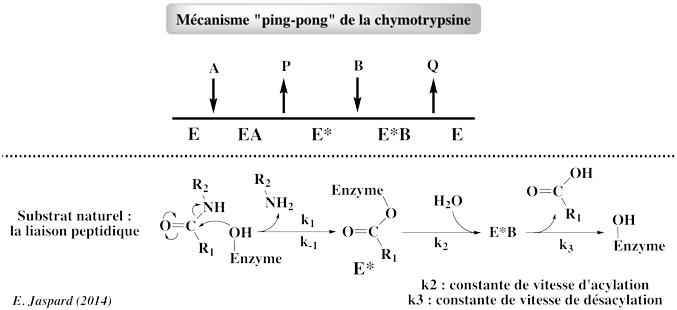
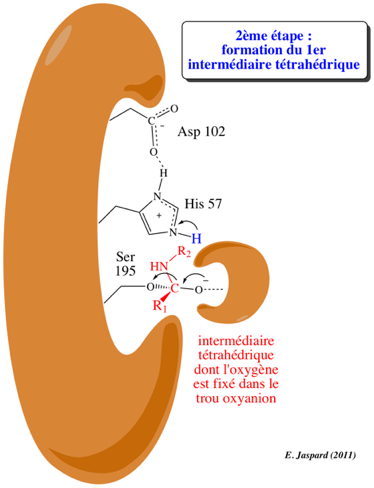
a. Le mécanisme des protéases à sérine : voir le cours sur le site actif des enzymes. b. Le mécanisme des protéases à cystéine est semblable à celui des protéases à sérine par l'utilisation d'un groupement nucléophile et la formation d'un complexe covalent [enzyme-substrat]. Le nucléophile est l'atome de soufre d'une cystéine (c'est un atome d'oxygène d'une sérine dans le cas des protéases à sérine). Dans la papaine, les groupements NH du squelette carboné de Gln 19 et Cys 25 forment le trou oxyanion.
c. Les protéases à acide aspartique contiennent 2 Asp qui agissent selon un mécanisme [acide-base] général. Une molécule d'eau coordonnée par les 2 Asp est activée par l'élimination d'un proton, ce qui permet à cette molécule d'eau polarisée d'attaquer le carbone carbonyle de la liaison peptidique. L'état de transition tétraédrique qui résulte de cette attaque nucléophile semble non chargé, contrairement à l'oxyanion formé lors de la catalyse par les protéases à sérine et à cystéine. d. Les métallo-protéases utilisent un métal coordonné (souvent le zinc) dans leur mécanisme catalytique. Dans beaucoup de métalloprotéases solubles (comme la thermolysine et les métallo-protéases de la matrice), cette coordination s'effectue avec 3 His ou 2 His et 1 chaîne latérale acide. Une molécule d'eau est un ligand supplémentaire du zinc. Cette molécule d'eau établit une liaison hydrogène avec un Glu qui lui arrache un proton et permet ainsi l'attaque. L'ion zinc stabilise l'oxyanion. Voir une animation du mécanisme catalytique des protéases à sérine. |
|
Visualisation de l'élastase pancréatique de Sus scrofa à une résolution de 1,65 Å Code PDB : 3EST Les 3 acides aminés His 57, Asp 102 et Ser 195 forment la triade catalytique.
|
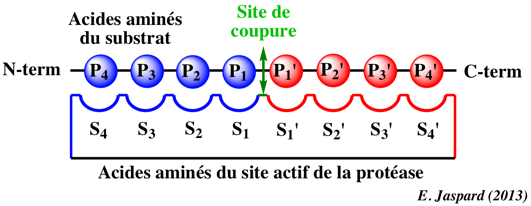
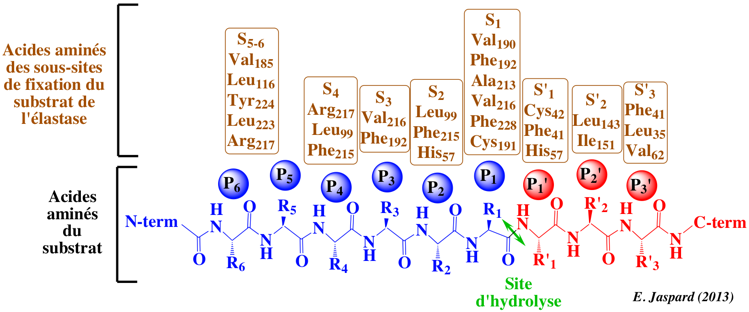
4. Le site de fixation du polypeptide substrat des protéases à sérine Le coeur du site de fixation du polypeptide substrat est la chaîne des carbones α des acides aminés 214 à 216 qui forment un feuillet β antiparallèle avec la chaîne des carbones α des acides aminés P1 à P3 du substrat.
Dans la chymotrypsine, des liaisons hydrogènes sont établies entre :
Ces interactions sont une caractéristique générale des protéases à sérine de type chymotrypsine et sont cruciales pour l'hydrolyse du substrat. Il est à noter que les acides aminés 214 à 216 forment une partie du sous-site S1 et que le groupement carbonyle de Ser 214 forme une liaison hydrogène avec His 57: il existe donc un lien structural direct entre le site de fixation du polypeptide substrat, le sous-site S1 et les acides aminés de la triade catalytique. Les boucles inhibitrices des protéines inhibiteurs des protéases sont bloquées dans la conformation étendue requise pour former le feuillet β antiparallèle avec le site de fixation du polypeptide substrat. Cette conformation étendue est observée dans de nombreux complexes [protéase-inhibiteur], y compris les complexes formés avec des analogues de l'état de transition et des peptidyl-acyl-enzymes. La conformation étendue inclue les acides aminés P3 à P'3, voire le groupement NH de l'acide aminé 218 et l'oxygène carbonyle en P5 comme dans le cas de l'élastase. La structure en feuillet β oriente les chaînes latérales du polypeptide substrat dans des directions alternées. |
|
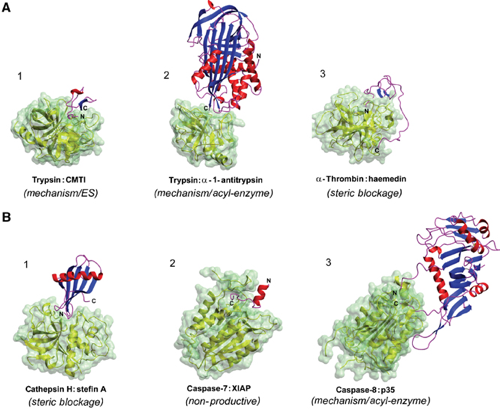
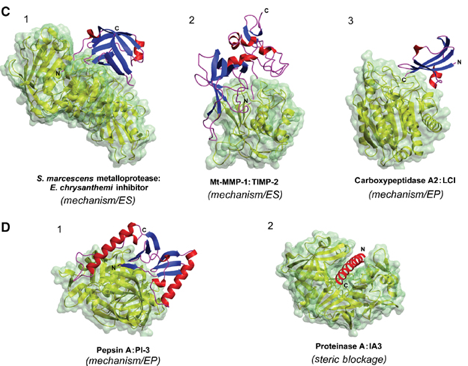
Exemples de complexes [protéase - inhibiteur] Figures ci-dessous :
Source : Otlewski et al. (2005) (A) Exemples de complexes [protéase à sérine - inhibiteur] : (1) trypsine - CMTI (PDB 1PPE) / (2) trypsine - α1-antitrypsine (PDB 1EZX) / (3) α-thrombine - haemedine (PDB 1E0F)
Source : Otlewski et al. (2005) (C) Exemples de complexes [métalloprotéase - inhibiteur] : (1) métalloprotease de Serratia marcescens - inhibiteur de Erwinia chrysanthemi (PDB 1SMP), (2) MMP-1 de type membranaire - TIMP-2 (PDB 1BQQ), (3) carboxypeptidase A2 humaine - LCI (PDB 1DTD) |
Le sous-site S1 Le sous-site S1 de fixation du substrat des protéases à sérine de type chymotrypsine est une poche située à côté de Ser195 et formée par les acides aminés 189-192, 214-216 et 224-228. La spécificité de reconnaissance du substrat est généralement déterminée par les acides aminés de l'enzyme situés aux positions 189, 216 et 226.
Les sous-sites S2-Sn Les sous-sites S2-S3 de la chymotrypsine sont peu discriminants vis-à-vis du polypeptide substrat, en accord avec la fonction de protéase digestive de chymotrypsine.
Les sous-sites S2-Sn des autres protéases à sérine peuvent être, au contraire, des déterminants majeurs de leur spécificité, comme l'élastase par exemple pour laquelle l'interaction S3/P3 est dominante ou l'entérokinase (appelée aussi entéropeptidase) qui reconnaît la séquence P1 à P5 [Asp-Asp-Asp-Asp-Lys] avec une extrême précision. Les sous-sites S'1-S'3 Les interactions entre le groupe partant du substrat et les sous-sites S'1-S'n sont en général déduites des structures des protéines inhibiteurs qui ne contiennent pas les acides aminés optimaux P'1-P'n.
Voir un descriptif détaillé de la spécificité de reconnaissance du polypeptide substrat d'un très grand nombre de protéases et des règles d'hydrolyse : "PeptideCutter" (Expasy). |
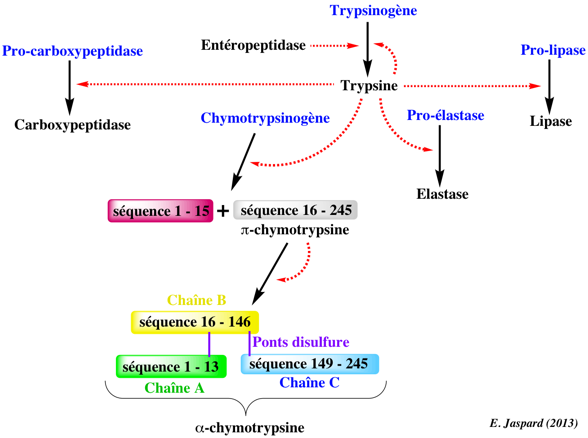
5. Les zymogènes : précurseur des protéases En 1876 - 1877, Wilhelm Kühne a découvert la trypsine dans le liquide pancréatique de l'intestin de boeuf au cours de ses études sur la digestion intermédiaire des protéines dans le tractus gastro-intestinal. Il a conclu que la trypsine était initialement inactive puis convertie en sa forme active. Cette observation est à la base de la notion de précurseur inactif des protéases que l'on appelle zymogène et de l'activation de ce zymogène qui peut être auto-catalytique ou catalysée par d'autres protéases. On distingue :
La taille du fragment hydrolysé varie d'un dipeptide (exemples : les granzymes, le trypsinogène) à des domaines entiers (exemples : cathepsines à cystéine, furine). Les pro-domaines de ces dernières sont censés agir également comme des chaperonnes intramoléculaires et sont nécessaires au trafic cellulaire de ces enzymes. Les enzymes protéolytiques sont biosynthétisées sous forme de précurseur inactif que l'on appelle le zymogène (exemples de zymogènes de protéases à sérine : chymotrypsinogène, trypsinogène, pro-élastase) dont la séquence polypeptidique contient une extrémité N-terminale qui est ultérieurement (auto-)hydrolysée.
Parfois, les protéases sont activées avec l'aide de complexes d'activation (exemples : certaines caspases situées en amont de cascades qui sont activées par DISC ("Death-Inducing Signalling Complex"), l'apoptosome, l'inflammasome, certaines protéases à sérine du plasma sanguin). Il existe des exceptions où la protéolyse n'est pas nécessaire pour l'activation, par exemples :
L'oligomérisation est un facteur important de l'activité enzymatique de certaines protéases : en effet, certaines sont inactives sous forme de monomère voire n'existent que sous forme d'oligomères comme le protéasome, la β-tryptase, la bléomycine hydrolase, les méprines ou la cathepsine C. L'inhibiteur tissulaire des métalloprotéases (TIMP-2 - "Tissue Inhibitor of MetalloProteases") est un cas très particulier : en plus de sa fonction inhibitrice, il peut participer à l'activation de MMP-2 ("Matrix MetalloProteinase-2" ou collagénase de type IV ou gélatinase A) par formation d'un complexe tri-moléculaire avec MT1-MMP ("Membrane Type 1-Matrix MetalloProteinase" - endopeptidase à zinc de type I transmembranaire) au sein duquel [TIMP-2/MT1-MMP] sert de récepteur pour réguler l'activation de MMP-2. Le domaine d'activation de zymogène Quatre segments de la chaîne polypeptidique sont déformés dans les zymogènes de la chymotrypsine et de la trypsine : les 19 premiers acides aminés N-terminaux, les acides aminés 142 à 152, 184 à 193 et 216 à 223. Ces quatre segments sont appelés collectivement domaine d'activation et ils incluent le sous-site S1 et le le trou oxyanion, ce qui explique la faible activité du zymogène. L'hydrolyse des 15 premiers acides aminés N-terminaux active le zymogène et fait apparaître un nouvel acide aminé N-terminal (Ile 16 pour la chymotrypsine, Val 16 pour l'élastase) qui forme un pont salin (enfoui dans la protéine) avec Asp 194. Ces événements induisent des changements locaux de conformation : le sous-site S1 et le le trou oxyanion sont formés, créant ainsi la forme active de la protéase. Voir une animation JMol : "Proteolytic cleavage of chymotrypsinogen by trypsin produces chymotrypsin". |
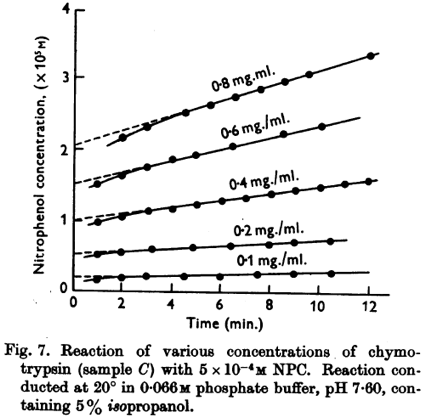
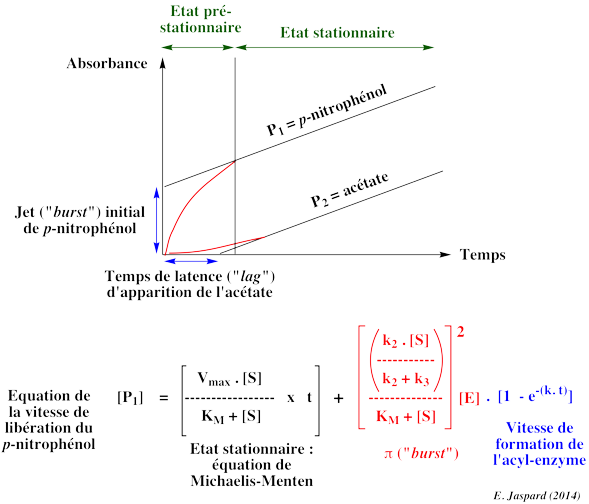
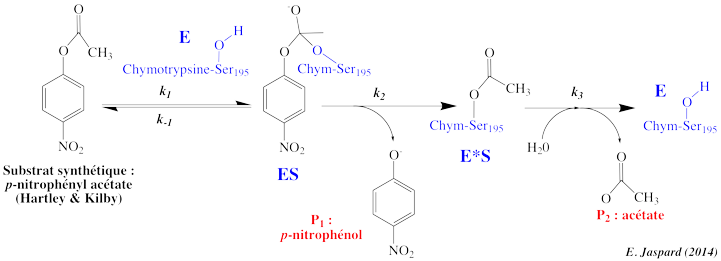
6. Mécanisme catalytique des protéases à sérine En 1953 - 1954, Brian Hartley et B. Kilby ont étudié la cinétique d'hydrolyse par la chymotrypsine en utilisant comme substrat le p-nitrophényl acétate (NPA) et le p-nitrophényléthyl carbonate (NPC), deux esters qui ressemblent à un acide aminé aromatique. L'hydrolyse du groupe carbonyle du NPA forme l'acétate et le p-nitrophénol qui absorbe à 400 nm (coloration jaune), et dont la concentration peut donc être mesurée par simple mesure spectrophotométrique. La réaction avec le NPA se déroule en 2 étapes :
Source : Hartley & Kilby (1954) Texte original de la figure : "In all measurements of the hydrolysis of NPA or NPC by chymotrypsin, it was observed that the extrapolated linear hydrolysis plot did not pass through the origin at zero time (Fig. 7). This suggested that the slow linear hydrolysis was preceded by a rapid initial reaction."
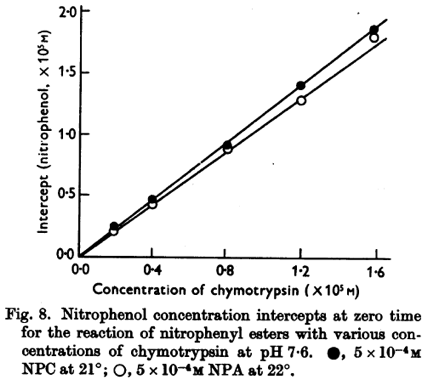
Source : Hartley & Kilby (1954) Texte original de la figure : "The linear portion of these curves gave rates proportional to enzyme concentration. In Fig. 8, the intercepts obtained by extrapolating the linear hydrolysis to zero time are plotted against enzyme concentration. The intercept is proportional to enzyme concentration. Such values are consistent, within experimental error, with a rapid initial reaction liberating 1 mole nitrophenol/mole enzyme." |
| Paramètres cinétiques de l'hydrolyse par la chymotrypsine de dérivés du N-acétyl-L-Trp (N-acétyl-L-Trp-X) | ||
| groupement X | kcat (s-1) | KM (μM) |
| -O-CH2-CH3 | 27 | 0.097 |
| -O-CH3 | 28 | 0.095 |
| -p-nitrophénol | 31 | 0.002 |
| -NH2 | 0.026 | 7.3 |
Influence de la nature du groupement partant sur l'hydrolyse par la chymotrypsine |
||
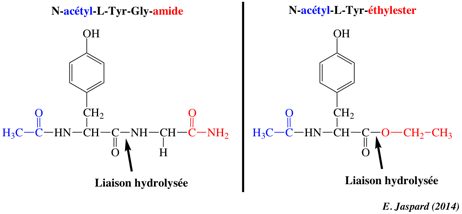
| Paramètre | Acétyl-Tyr-Gly-amide | Acétyl-Tyr-O éthyl-ester |
| kcat (s-1) | 0,50 | 193 |
| KM (M) | 0,023 | 0,0007 |
| kcat/KM (M-1.s-1) | 21,7 | 275.700 |
On observe une formation trés rapide de p-nitrophénol durant l'état pré-stationnaire : c'est ce que l'on appelle le jet ou "burst" (figure ci-dessous et figure 7 ci-dessus).
Cette phase est suivie d'une formation plus lente de p-nitrophénol à l'état stationnaire. Au bout d'un temps appelé temps de latence (ou "lag"), on observe la formation d'acétate. L'étape d'hydrolyse est la plus lente : c'est l'étape limitante. La cinétique d'apparition des 2 produits reflète la formation rapide d'une forme intermédiaire de l'enzyme : l'acyl-enzyme.
|
|
Le mécanisme réactionnel s'écrit : k1 k2 k3 Hypothèse de l'état stationnaire : d[E*S]/dt = 0 => (k2 . [ES]) - (k3 . [E*S]) = 0 |
| [S0] vi = kcat . [E0] . -------------- KMapp + [S0] |
Paramètres cinétiques : k2 . k3 k3
Si k2 << k3 : l'étape limitante est l'acylation - dans ce cas : kcat = k2 et KMapp = KM Si k2 >> k3 : l'étape limitante est la désacylation - dans ce cas : kcat = k3 et KMapp = (k3/k2) KM |
|
Voir un cours sur les cinétiques à 2 (ou plus) substrats. Exemples d'enzymes qui suivent un mécanisme "Ping Pong" : les oxydoréductases (exemple : thiorédoxine peroxydase), les transférases (exemple : acylneuraminate cytydilyltransférase), les protéases à sérine. Ce mécanisme est aussi appelé réaction à double déplacement : un ou plusieurs produits sont libérés avant que tous les substrats se soient fixés à l'enzyme (figure ci-dessous - selon la représentation de William Cleland) :
Une caractéristique fondamentale de ce type de mécanisme est donc l'existence d'une forme intermédiaire E* de l'enzyme, chimiquement modifiée : l'acyl-enzyme. Dans le cas des protéases à sérine :
Voir un détail structural du mécanisme catalytique des protéases à sérine et la formation des acyl-enzymes. |
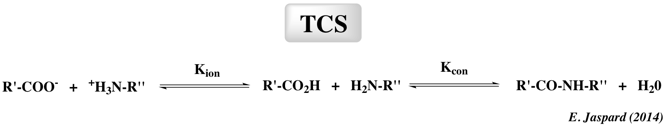
d. Synthèse peptidique par les protéases Il y a plus de 100 ans, Jacobus van't Hoff (1898) a souligné la possibilité de formation enzymatique de la liaison peptidique par réversion de la réaction d'hydrolyse des protéases. Depuis lors, la synthèse catalysée par les protéases a été largement établie. Des petits peptides synthétisés par voie enzymatique sont utilisés pour la nutrition humaine ou animale et aussi comme produits agrochimiques ou pharmaceutiques. Exemples : l'aspartame (édulcorant non calorique), la kyotorphine, l'angiotensine, l'enképhaline et la dynorphine et quelques dipeptides ou tripeptides de nutritionnels. La synthèse de peptides par des protéases s'effectue selon 2 mécanismes : le contrôle thermodynamique et le contrôle cinétique. La synthèse de peptides thermodynamiquement contrôlée ("Thermodynamically Controlled Synthesis" - TCS) avec des protéases représente l'inverse de l'hydrolyse de la liaison peptidique.
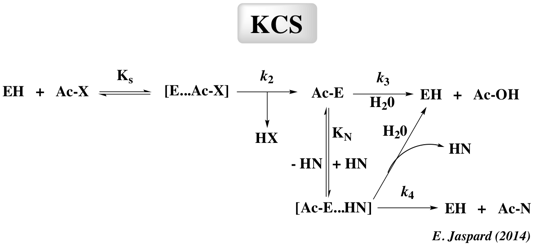
Kion : constante d'équilibre d'ionisation En effet, selon le principe de réversibilité microscopique, la formation et l'hydrolyse de la liaison peptidique suivent le même mécanisme et mettent en jeu les mêmes intermédiaires. La formation de l'intermédiaire acyle à partir d'un acide carboxylique est une réaction très lente et représente l'étape limitante de la TCS. Le contrôle cinétique de la synthèse des peptides ("Kinetically Controlled Synthesis" - KCS)
Comme le montre le schéma, le donneur d'acyle (qui doit être activé sous forme d'un ester, d'une amide ou d'un nitrile) se fixe en premier à l'enzyme pour former un complexe [enzyme- substrat] tétrahedrique [E...AC-X] qui à son tour forme l'acyl-enzyme intermédiaire covalent [Ac-E]. Cet intermédiaire subit une attaque nucléophile par l'eau ou par un nucléophile (HN) qui peut être une amine, un alcool ou un thiol, en compétition avec l'eau pour la réaction de désacylation. La réussite de la réaction de synthèse dépend des cinétiques de ces réactions nucléophiles, ce qui explique la notion de contrôle cinétique de ce type de synthèse. A l'inverse de la stratégie TCS, seules les protéases à sérine ou à cystéine peuvent être utilisés pour la stratégie KCS, car dans ce cas, l'enzyme agit comme une transférase et catalyse le transfert d'un groupement acyle du donneur d'acyle à des acides aminés nucléophiles. Généralement, la stratégie KCS est plus rapide et nécessite un rapport [enzyme/substrat] plus faible que la stratégie TCS parce que le donneur d'acyle est sous forme d'un acide carboxylique activé. Il est souhaitable que le produit peptidique de la réaction soit retiré du milieu réactionnel pour éviter une hydrolyse secondaire indésirable. Le rendement en peptide synthétisé dépend du rapport apparent des constantes de vitesse des activités [transférase / hydrolase] et de la vitesse à laquelle le produit peptidique est hydrolysé. Les protéases les plus utilisées dans la stratégie KCS sont la thermolysine, la papaïne, la trypsine et l'α-chymotrypsine avec des des rapport d'activités [transférase / hydrolase] de l'ordre de 102-104. Voir : Guzman et al. (2007) "Peptide synthesis: chemical or enzymatic" Elec. J. Biotechnol. 10, 279 - 314 |
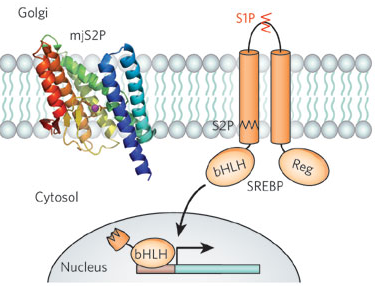
7. Les protéases intramembranaires L'étude des protéases est entrée dans une nouvelle phase avec la découverte de protéases intramembranaires qui catalysent l'hydrolyse de la liaison peptidique dans les membranes. Jusqu'à lors, on croyait les réactions hydrolytiques restreintes aux seules enzymes cytosolubles. La protéolyse intramembranaire fournit une stratégie simple et irréversible pour la signalisation : le clivage et la libération de domaines protéiques attachés à la membrane. Ces domaines peuvent, par exemple, pénétrer dans le noyau et agir comme facteurs de transcription ou être excrétés afin d'activer les récepteurs de cellules voisines. La protéolyse intramembranaire a été décrite pour la première fois comme une activité essentielle dans l'homéostasie des stérols :
Source : Erez et al. (2009) Le domaine cytoplasmique de la protéine de fixation de l'élément de régulation des stérols des mammifères ("mammalian Sterol Regulatory Element-Binding Protein" - mSREBP) est attaché par un segment transmembranaire à la membrane du réticulum endoplasmique. Lorsque le niveaux de cholestérol de la cellule chute, mSREBP est transportée vers l'appareil de Golgi où deux métallo-protéases intramembranaires ("Site-1 Protease" - S1P puis "Site-2 Protease" - S2P) clivent son domaine cytoplasmique. Le domaine cytoplasmique (bHLH) ainsi libéré est dirigé vers le noyau, où il active les gènes de la synthèse du cholestérol et des acides gras. Plusieurs protéases intramembranaires ont depuis été identifiées et elles constituent 3 groupes :
|
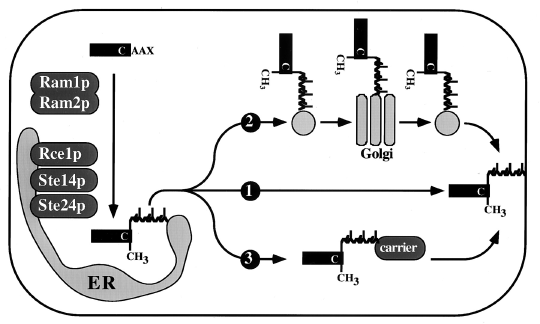
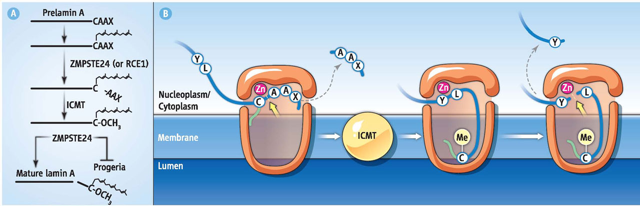
8. Les protéases intramembranaires des protéines prénylées à motif CAAX Voir un cours sur la prénylation. Un grand nombre de protéines (superfamille RAS des petites GTPases, les lamines du noyau, la sous-unité γ des GTPases hétérotrimériques et plusieurs protéines kinases et protéines phosphatases) qui ont un motif C-terminal CAAX (C : Cys, A : un acide aminé aliphatique, X : n'importe quel acide aminé) subissent des modifications post-traductionnelles par des lipides (prénylation) isoprénoïdes (groupes farnésyl ou géranyl-géranyl). Ces groupes sont attachés à la cystéine. L'isoprènylation est suivie par l'hydrolyse des acides aminés AAX et la méthylation de la cystéine prénylée. La biogénèse des protéines prénylées à motif CAAX nécessite un routage à partir de la membrane du réticulum endoplasmique (RE). Bien que la maturation des protéines prénylées à motif CAAX ait lieu au niveau de cette membrane, ces protéines sont destinées à d'autres types de membranes (plasmique par exemple). Le transport des protéines prénylées à motif CAAX telles que Ras de la membrane du RE vers la membrane plasmique peut impliquer l'un des 3 mécanismes suivants :
Source : Schmidt et al. (1998)
Actuellement, on a identifié 2 protéases intramembranaires des protéines prénylées à motif CAAX. |
|
Remarque préalable : diverses appellations de Ste24p (EC 3.4.24.84) selon l'origine
Ste24p est une métallo-protéase à zinc (endopeptidase de type I). Elle a un rôle spécifique dans la maturation de la prélamine A (précurseur de la lamine A, protéine filament intermédiaire du noyau) chez les Eucaryotes et du facteur a chez la levure.
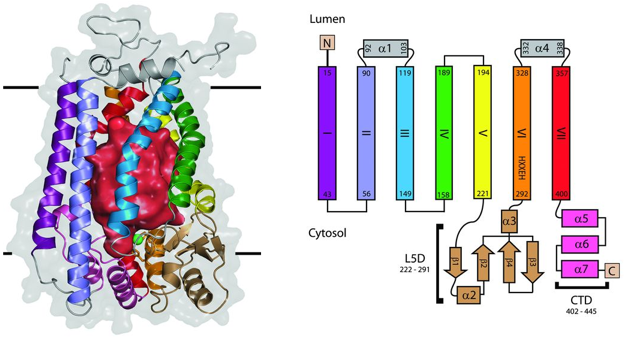
Source : Pryor et al. (2013) SmSte24p de la levure Saccharomyces mikatae est une protéine intégrale de la membrane composée de 7 hélices transmembranaires (de 28 à 44 acides aminés) avec des écarts jusqu'à 10 Å de diamètre entre les hélices (figure ci-dessous).
Figure ci-dessous : structure de la protéase SmSte24p de la levure Saccharomyces mikatae.
Source : Pryor et al. (2013) La surface de la protéine exposée au cytoplasme contient 2 domaines supplémentaires :
Actuellement, les acides aminés du site actif de SmSte24p ne sont pas clairement définis.
|
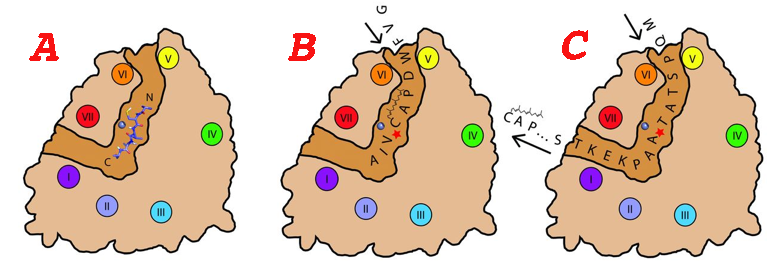
Mécanisme catalytique supposé de la protéase SmSte24p Actuellement, le mécanisme de l'hydrolyse s'appuie sur l'hypothèse d'un processus d'insertion du substrat, de sa translocation puis de l'éjection du premier produit. Le sillon de fixation du substrat, formé par L5D et CTD, se situe sur la face exposée au cytoplasme de la cavité interne transmembranaire de SmSte24p. Ce sillon, presque coplanaire à la surface de la membrane a une longueur d'environ 40 Å et traverse la cavité (figure ci-dessous - mécanisme catalytique proposé pour SmSte24p). Figure A : le fort degré de similitude entre les sites actifs de SmSte24p et de la thermolysine suggère que la polarité (direction N- vers C-) des substrats respectifs est identique dans les deux enzymes. En conséquence, les substrats de SmSte24p seraient orientés dans le sillon avec leurs extrémités C-terminale pointant vers l'embouchure située entre les hélices I et VII et leurs extrémités N-terminale pointant vers l'embouchure située entre les hélices V et VI.
Source : Pryor et al. (2013) Figure B : l'étoile rouge indique le site d'hydrolyse du motif CAAX. Figure C : translocation du substrat qui suit cette hydrolyse, puis évacuation du produit d'hydrolyse par l'embouchure située entre les hélices I et VII et enfin positionnement du deuxième site d'hydrolyse. La consommation d'une molécule d'eau à chaque acte catalytique suggère que la cavité interne volumineuse constitue un "réservoir d'eau", maximisant ainsi la concentration effective de l'eau et augmentant la catalyse des protéines prénylées à motif CAAX. D'autres protéases intramembranaires adoptent des mécanismes d'accès de l'eau au site catalytique différents.
|
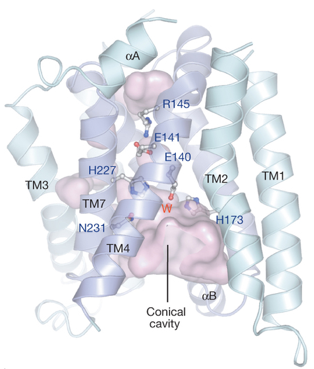
b. La protéase Rce1 ("Ras and a-factor converting enzyme 1") Elle a été identifiée chez Saccharomyces cerevisiae : elle serait l'un des premiers membres identifiés d'une classe de protéases intramembranaires à glutamate (endopeptidase de type II - EC 3.4.22) du réticulum endoplasmique. Sa spécificité est beaucoup plus large dans la maturation de toutes les protéines prénylées à motif CAAX. Rce1 appartient à la famille ABI ("ABortive Infection") des protéases intramembranaires potentielles et possède des homologues dans les 3 règnes du vivant. La famille ABI est définie par trois motifs conservés qui constituent leur site catalytique. Rce1 contient 8 hélices α transmembranaires dont la structure et celle de son site catalytique sont différentes des autres protéases intramembranaires. Le domaine ABI contient environ 100 acides aminés qui correspondent aux hélices α transmembranaires TM4 à TM7. Ces quatre hélices forment un faisceau d'hélices antiparallèles (symétrie d'ordre 2) entouré par les hélices moins conservées TM1, TM2, TM3 et TM8. 7 hélices α transmembranaires forment une large cavité catalytique conique (d'un volume d'environ 1400 Å3) qui accomode les protéines prénylées à motif CAAX. Les acides aminés catalytiques (E140, H173, H227 et N231) sont situés dans la partie supérieure de cette cavité, à environ 10 Å à l'intérieur de la membrane. Figure ci-dessous : structure de la protéase Rce1
Source : Manolaridis et al. (2013) Cette conformation des hélices α transmembranaires permet un accès illimité du solvant du cytoplasme jusqu'au site catalytique de cette cavité. Un côté de la cavité est ouvert côté membrane par un espace situé entre TM2 et TM4. L'accès au périplasme (la lumière du réticulum endoplasmique) est bloqué par l'acide aminé conservé R145, qui interagit avec T210 et acide aminé conservé E141. Le modèle structural actuel propose que le groupe farnésyle se fixe sur un site situé à l'ouverture de 2 hélices α transmembranaires, positionnant ainsi la liaison peptidique à hydrolyser à côté d'une molécule d'eau nucléophile activée par Glu140. |
| 12. Liens Internet et références bibliographiques |
|
The MEROPS database : information resource for peptidases (also termed proteases, proteinases and proteolytic enzymes) and the proteins that inhibit them PROSPER: Protease substrate specificity webserver Serine Proteases |
|
|
Hartley & Kilby (1954) "The reaction of p-nitrophenyl esters with chymotrypsin and insulin" Biochemical Journal 56, 288 - 297 Schechter & Berger (1967) "On the size of the active site in proteases" Biochem. Biophys. Res. Com. 27, 157 - 162 Schechter & Berger (1968) "On the active site of proteases. 3. Mapping the active site of papain; specific peptide inhibitors of papain" Biochem. Biophys. Res. Commun. 32, 898 - 902 Jaspard E. (2000) "Role of protein-solvent interactions in refolding: effects of cosolvent additives on the renaturation of porcine pancreatic elastase at various pHs" Arch. Biochem. Biophys. 375, 220 - 228 Hedstrom L. (2002) "Serine protease mechanism and specificity" Chem. Rev. 102, 4501 - 4524 |
|
|
Vocadlo et al. (2001) "Catalysis by hen egg white lysozyme proceeds via a covalent intermediate" Nature, 412, 835 - 3838 Feng et al. (2007) "Structure of a Site-2 Protease Family Intramembrane Metalloprotease" Science 318, 1608 - 1612 Erez et al. (2009) "How intramembrane proteases bury hydrolytic reactions in the membrane" Nature 459, 371 - 378 Li et al. (2013) "Commercial proteases: Present and future" FEBS Let. 587, 1155 - 1163 |
|
|
Schmidt et al. (1998) "Endoplasmic reticulum membrane localization of Rce1p and Ste24p, yeast proteases involved in carboxyl-terminal CAAX protein processing and amino-terminal a-factor cleavage" PNAS 95, 11175 - 11180 Pryor et al. (2013) "Structure of the integral membrane protein CAAX protease Ste24p" Science 339, 1600 - 1604 Quigley et al. (2013) "The Structural Basis of ZMPSTE24-Dependent Laminopathies" Science 339, 1604 - 1607 Manolaridis et al. (2013) "Mechanism of farnesylated CAAX protein processing by the intramembrane protease Rce1" Nature 504, 301 - 305 |
|
|
Otlewski et al. (2005) "The many faces of protease–protein inhibitor interaction" EMBO J. 24, 1303 - 1310 Drag & Salvesen (2010) "Emerging principles in protease-based drug discovery" Nat. Rev. Drug Discov. 9, 690 - 701 Vandermarliere et al. (2013) "Getting intimate with trypsin, the leading protease in proteomics" Mass Spectro. Rev. 32, 453 - 465 |
![]()