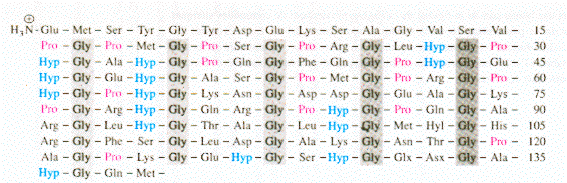| Les types de collagène |
| Tweet |
|
|
1. Les types de collagène 2. Bioynthèse des procollagènes et du tropocollagène 3. Hydrolyse des propeptides des procollagènes 4. Le tropocollagène 5. Les aggrégats de fibrilles et structure périodique des collagènes |
6. Les séquences télopeptides (lysine et des hydroxylysine) 7. Modification des lysines par des oses 8. Hydroxyproline 9. La matrice extracellulaire 10. Liens Internet et références bibliographiques |
1. Les types de collagène Les collagènes sont les protéines majeures de la plupart des tissus conjonctifs. Les collagènes sont les protéines prépondérantes chez les vertébrés (30% des protéines totales des Mammifères : peau, os, dents, tendons, vaisseaux sanguins).
Source : Aslan et al. (2008)
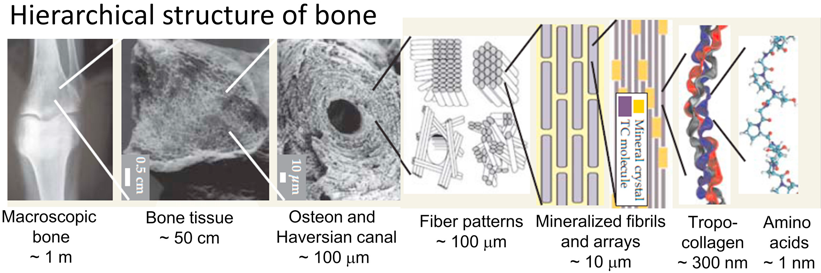
Source : Sherman et al. (2015) Dans le génome humain, il existe 44 gènes qui codent une chaîne polypeptidique précurseur d'un type de collagène : ces chaînes polypeptidiques précurseurs se combinent de diverses façons pour créer 28 types différents de fibrilles de collagène (désignés par des chiffres romains). |
| Type de collagène | Composition de la triple hélice | Symbole du gène | Structure | Remarques |
| I: précurseur de fibrilles | [α1(I)]2[α2(I)] * | COL1A1 COL1A2 |
300 nm fibrilles 67 nm** |
peau, tendon, os, ... / type de collagène le plus abondant dans le corps humain - jusqu'à 90% de toutes les protéines de collagène |
| II: précurseur de fibrilles | [α1(II)]3 | COL2A1 | 300 nm fibrilles courtes 67 nm |
homotrimère / cartilage, vitreous humor |
| III: précurseur de fibrilles | [α1(III)]3 | COL3A1 | 300 nm fibrilles courtes 67 nm |
homotrimère / peau, muscle, associé au type I |
| IV: formeur de feuillets | [α1(IV)]2[α2(IV)] | COL4A1 à COL4A6 |
domaine globulaire C-terminal 390 nm non fibrillaire |
Toute lame basale. C'est la composante structurale majeure de toutes les membranes basales. COL4A3 code pour l'antigène lié au syndrome appelé de « Goodpasture » |
| V: précurseur de fibrilles | [α1(V)][α2(V)][α3(V)] | COL5A1 COL5A2 COL5A3 |
domaine globulaire N-terminal 390 nm fibres courtes |
la plupart des tissus interstitiels, associé au type I |
| VI: précurseur de fibrilles | [α1(VI)][α2(VI)][α3(VI)] | COL6A1 COL6A2 COL6A3 |
domaine globulaire N- et C-terminaux 150 nm microfibrilles |
la plupart des tissus interstitiels, associé au type I |
| VII: ancrage | [α1(VII)]3 | COL7A1 | 450 nm | homotrimère / épithelium |
| VIII | [α1(VIII)]3 | COL8A1 COL8A2 |
homotrimère / certaines cellules endothéliales | |
| IX: ancrage | [α1(IX)][α2(IX)][α3(IX)] | COL9A1 COL9A2 COL9A3 |
domaine globulaire N-terminal 200 nm protéoglycane |
cartilage de type "FACIT collagen" ("Fibril Associated Collagens with Interrupted Triple helices"), associé au type II |
| X | [α1(X)]3 | COL10A1 | domaine globulaire C-terminal 150 nm | homotrimère / cartilage hypertrophique et minéralisant |
| XI | [α1(XI)][α2(XI)][α3(XI)] | COL11A1 COL11A2 |
300nm fibres courtes |
cartilage |
| XII: ancrage | [α1(XII)] | COL12A1 | homotrimère / collagène FACIT/ tendons, peau, placenta / interagit avec les types I et III | |
|
Ne sont décrits dans ce tableau que les 12 principaux types de collagène sur les 28 types recensés. *Exemple : deux des trois protéines de la triple hélice de collagène de type I sont codées par le gène COL1A1 et la troisième protéine est codée par le gène COL1A2. Les fibrilles de collagène de type I sont donc désignées [α1(I)]2[α2(I)] où le chiffre romain désigne le type de fibrille. **Les interactions latérales au sein de la triple hélices de collagène entraînent la formation de fibrilles d'environ 50 nm de diamètre. La compaction du collagène est telle que les molécules adjacentes sont déplacées d'environ 1/4 de leur longueur (soit environ 67nm). Cette matrice décalée produit un effet strié qui est visible en microscopie électronique. Source : "The Extracellular Matrix " - The Medical Biochemistry Page |
||||
| Classe de collagène | Type* | Gène | Remarques |
| Fibrillaire | I | COL1A1, COL1A2 | fibrilles principalement dans la matrice stromale fibreuse (exemple : peau, os, tendons et ligaments) - les collagènes fibrillaires de types I et III sont les principaux composants de la matrice extracellulaire cardiaque |
| II | COL2A1 | ||
| III | COL3A1 | ||
| V | COL5A1, COL5A2, COL5A3 | ||
| XI | COL11A1, COL11A2, COL11A3 | ||
| XXIV | COL24A1 | ||
| XXVII | COL27A1 | ||
| FACIT | IX | COL9A1, COL9A2, COL9A3 | "ponts moléculaires" associés aux fibrilles de collagène de type I (XII, XVI, XIX, XXI) et de type II (IX, XVI, XIX) |
| XII | COL12A1 | ||
| XIV | COL14A1 | ||
| XVI | COL16A1 | ||
| XIX | COL19A1 | ||
| XX | COL20A1 | ||
| XXI | COL21A1 | ||
| XXII | COL22A1 | ||
| Membrane basale | IV | COL4A1, COL4A2, COL4A3, COL4A4, COL4A5, COL4A6 | structure en réseau composée de laminines et de protéines de la membrane basale |
| Longue chaîne | VII | COL7A13 | fibrilles ancrées associées à la membrane basale |
| Filamenteux | VI | COL6A1, COL6A2, COL6A3, COL6A5, COL6A6 | microfibrilles "perlées" |
| Courte chaîne | VIII | COL8A1 | structure en treillage hexagonal |
| X | COL10A1 | ce type de collagène régule et est régulé par une hypertrophie du cartilage | |
| Multiplexines | XV | COL15A1 | domaines à triple hélice multiples avec des interruptions contenant des glycosaminoglycanes (sulfate de chondroïtine et sulfate d'héparine) |
| XVIII | COL18A1 | ||
| MACIT | XIII | COL13A1 | molécules de surface de la cellule avec des domaines extracellulaires, membranaires et intracellulaires |
| XVII | COL17A1 | ||
| XXIII | COL23A1 | ||
|
*Les types de collagène sont classés sur la base de l'homologie de la structure du domaine et de l'assemblage supra-structural. MACIT - "Membrane-Associated Collagens with Interrupted Triple helices" (domaine transmembranaire) Source : Mouw et al. (2014) |
|||
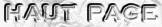
|
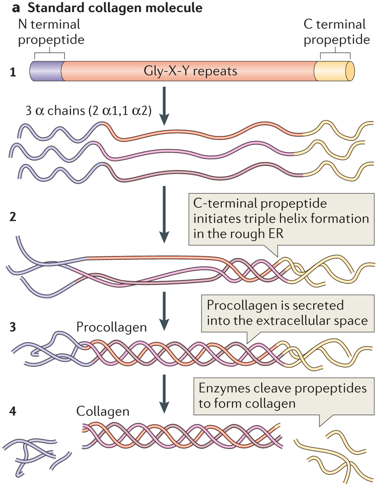
2. Bioynthèse des procollagènes et du tropocollagène Les collagènes fibrillaires (de types I, II, III, V et XI) sont synthétisés sous forme de précurseurs solubles, les procollagènes. Ces chaînes précurseur sont transportées de manière co-traductionnelle dans le lumen du réticulum endoplasmique (RE) où elles subissent des modifications pour maturer les molécules des différents types de collagène. Ci-dessous, schéma général des principales étapes de biosynthèse et de maturation des chaînes polypeptidiques qui forment un type de collagène fibrillaire :
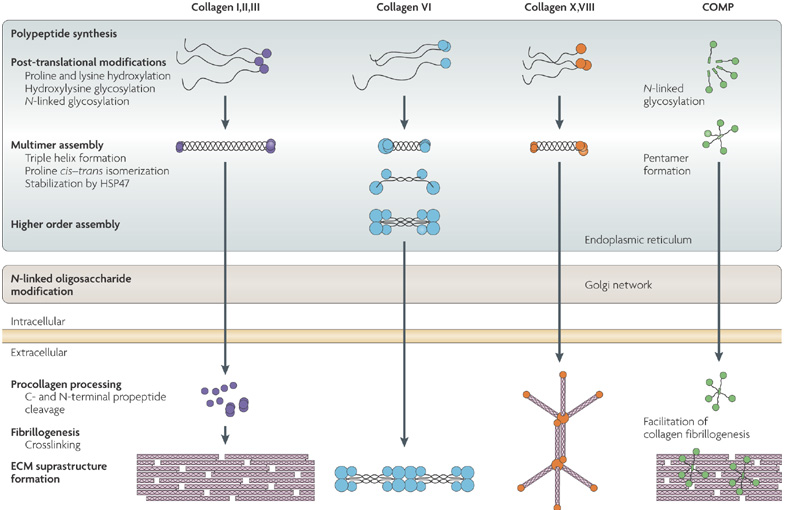
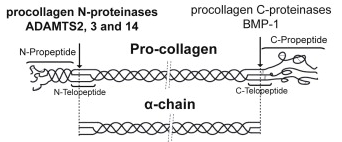
Source : Mouw et al. (2014) La maturation continue dans l'appareil de Golgi puis le procollagène de type I est sécrétée par exocytose. Des procollagène peptidases hydrolysent les propeptides N- et C-terminaux dans le milieu extra-cellulaire :
Le rythme de renouvellement du réseau de collagène est estimé à 80 - 120 jours (équilibre synthèse - dégradation des fibres de collagène). Voir un schéma récapitulatif de ces évènements. Figure ci-dessous : principales similitudes et différences dans les voies de synthèse et d'assemblage de plusieurs classes de collagène
Source : Bateman et al. (2009) |
|
3. Hydrolyse des propeptides des procollagènes Les procollagènes sont constitués d'une région centrale en triple hélice en forme de tige (environ 300 kDa) avec des propeptides globulaires aux extrémités N-terminale (environ 50 kDa) et C-terminale (environ 90 kDa). Ces domaines propeptides sont hydrolysés par des métalloendopetidases.
Source : Fan et al. (2012) "Fibrogenesis Tissue Repair" Les protéases BTP ("Bone morphogenetic protein-1 (BMP-1)/Tolloid-like Proteinases") - EC 3.4.24.19
Source : Bekhouche & Colige (2015) Hydrolyse du propeptide N-terminal : ADAM / ADAMTS Les métalloendopeptidases ADAM2, 3 et 14 clivent le propeptide N-terminal des procollagènes fibrillaires. Il existe une forte homologie de séquence des sites de clivage par ADAMTS2 des procollagènes fibrillaires de types I à III de différentes espèces de mammifères : PGMP435/A436NQD. Le site de clivage potentiel semble se situer à la position 435/436 en amont du domaine de la triple hélice. ADAM / ADAMTS ("A Disintegrin And Metalloproteinase domain-containing protein" / "A Disintegrin And Metalloproteinase with ThromboSpondin type I domain") :
Hydrolyse du propeptide C-terminal : BMP-1 La métalloendopetidase à zinc BMP-1 ("Bone Morphogenetic Protein-1") est constituée d'un domaine catalytique de type astacine suivi de plusieurs domaines CUB ("Complement C1r / C1s, uEGF, BMP-1") et de domaines EGF. BMP-1 appartient au clan metzincine lui-même constitué :
Chez les vertébrés, BMP-1 est impliquée dans la maturation d'une grande variété de précurseurs de la matrice extracellulaire nécessaires à l'assemblage normal de la matrice extracellulaire et des tissus. Ces substrats incluent :
La laminine-5 est un élément clé des filaments d'ancrage qui aident les kératinocytes à adhérer à la membrane basale. La laminine-5 est un trimère composé de chaînes α3, β3 et γ2 : BMP1 sépare les chaînes γ2 de la laminine-5 de l'homme et celle du rat. |
|
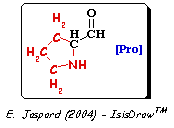
C'est l'unité de base du collagène. Dimensions : longueur : environ 300 nm - diamètre : 1.5 nm. Le tropocollagène est composé de 3 chaînes polypeptidiques hélicoïdales gauches torsadées l'une autour de l'autre en une superhélice de pas droit. Ces hélices ne sont pas comparables aux hélices α, incompatibles avec la présence d'une proline. En effet, la proline est un acide α-iminé (groupe aminé secondaire). Sa chaîne latérale est liée à la fois au groupe α-carboxyle et au groupe α-aminé.
Source : Protein Data Bank |
|
Visualisation du tropocollagène de l'hommeà une résolution de 2 Å Code PDB : 1BKV Les 3 chaînes (A, B et C) sont colorées.
|
|
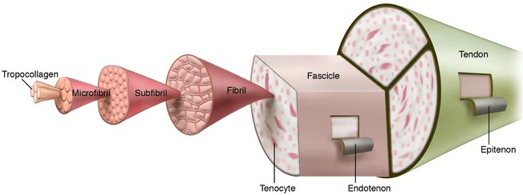
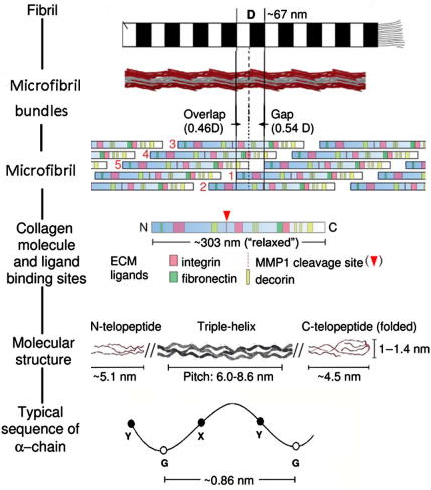
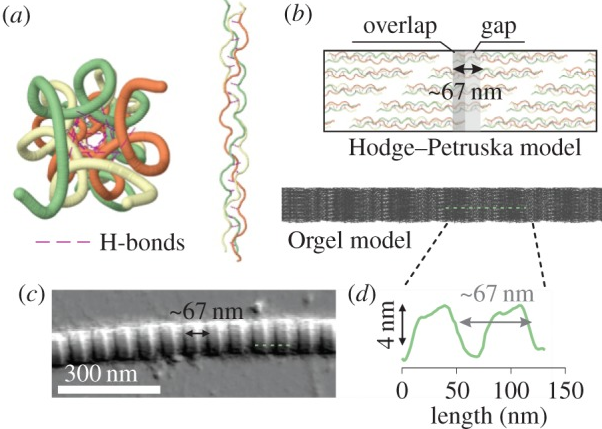
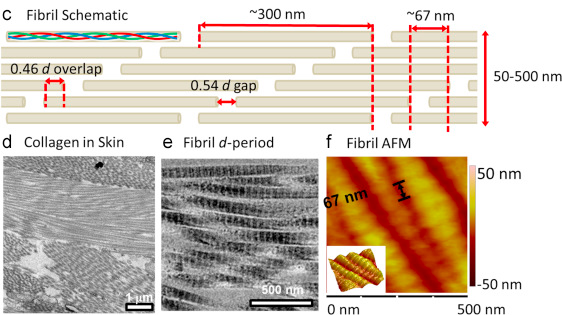
5. Les aggrégats de fibrilles et structure périodique des collagènes Le tropocollagène compose une molécule bien plus grande : les aggrégats de fibrilles.
Source : Bertassoni et al. (2012) Dans les fibrilles, les segments de triple hélices sont décalés les uns par rapport aux autres. Cette disposition créée :
Ces 2 régions ont une périodicité (appelée période D) de ≈ 67 nm. C'est la raison pour laquelle les fibrilles de collagène ont une apparence en forme de bandes.
Source : Andriotis et al. (2015)
Source : Sherman et al. (2015) |
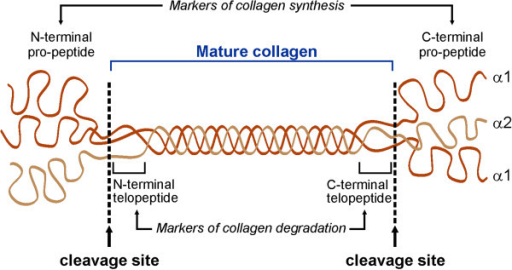
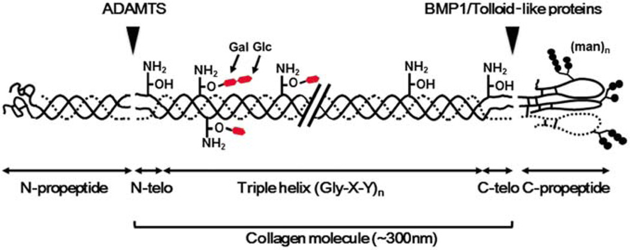
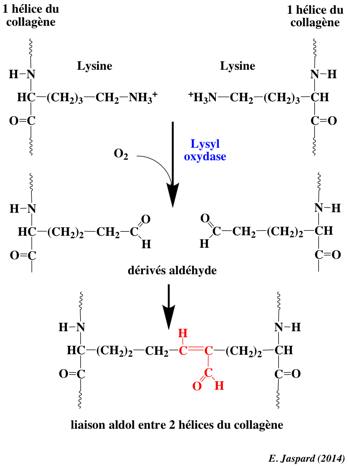
6. Les séquences télopeptides (lysine et hydroxylysine) Le collagène de type I est un hétérotrimère (longueur ≈ 300 nm / épaisseur ≈ 1,5 nm) composée de deux chaînes α1 et d'une chaîne α2. Il existe une forme mineure homotrimérique α1. Chaque chaîne polypeptidique se compose de trois domaines :
Les télopeptides N- et C-terminaux jouent un rôle capital dans la formation des liaisons covalentes entre les 3 chaînes du tropocollagène pour la formation de fibrilles de collagène solides.
Source : Yamauchi & Sricholpech (2012) Les télopeptides n'adoptent donc pas la conformation triple hélice et contiennent l'acide aminé inhabituel hydroxylysine. Une liaison covalente aldol est établie entre 2 résidus lysine ou hydroxylysine de l'extrémité C-terminale d'une molécule de collagène avec 2 résidus identiques de l'extrémité N-terminale d'une molécule adjacente. Ces réticulations compactent les molécules de collagène et génèrent des fibrilles fortes.
Des résidus spécifiques de lysine et d'hydroxylysine dans les 2 télopeptides sont désaminés par oxydation par la lysyl oxidase ("Protein-lysine 6-oxidase" - LOX) pour former respectivement des dérivés aldéhyde (acide α-aminoadipique-δ-semialdéhyde) de ces résidus spécifiques, qui initient une série de réactions de condensation pour former des liaisons covalentes intra et inter-moléculaires. Remarque : les pyridinolines sont relarguées de la matrice osseuse au cours de sa résorption par les ostéoclastes. Ces molécules ("aminoterminal cross-linked telopeptides of type I collagen" - NTX1" et "carboxyterminal cross-linked telopeptides of type I collagen" - CTX1) servent de marqueurs de dégradation du collagène. La réaction catalysée par LOX est : R-CH2NH2 + H2O + O2 ---> R-CH=O + NH3 + H2O2 Dans le collagène de type I, il y a 5 résidus lysine ou hydroxylysine des télopeptides qui fournissent le groupe aminé de cette réaction :
Exemples de séquences de télopeptides du collagène de type I de bos taurus :
Séquence du télopeptide C-terminal de la chaîne α1 de l'homme : -S1AGFDFSFLPQPPQEK16AHDGGRYYRA |
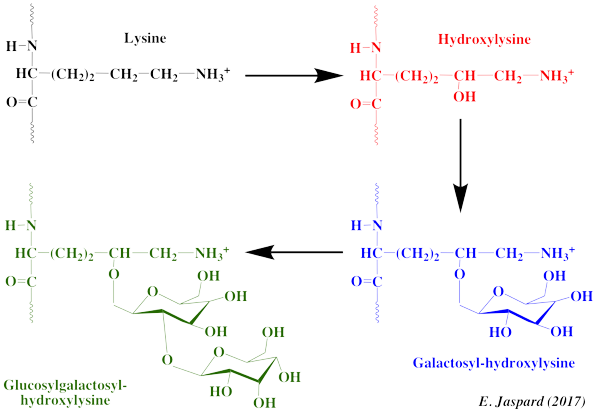
7. Modification des lysines par des oses Dans le collagène de type I de l'homme, il y a :
La lysyl hydroxylase ("Procollagen-lysine,2-oxoglutarate 5-dioxygenase 2" - EC 1.14.11.4) catalyse la formation d'hydroxylysine dans des séquences -X-K-G- en présence de Fe2+, de 2-oxoglutarate, d'O2 et d'ascorbate. Certains résidus hydroxylysine du domaine central hélicoïdal sont encore modifiés :
Structure du complexe 2-O-α-D-glucopyranosyl-O-β-D-galactopyranosyl-hydroxylysine :
Les deux réactions nécessitent la présence du groupe ε-aminé libre de l'hydroxylysine, de l'UDP-galactose et de l'UDP-glucose (donneurs de glucides) et le de cation bivalent Mn2+ (cofacteur). |
|
Figure ci-dessous : séquence des 139 premiers résidus d'une chaîne de collagène de peau de rat.
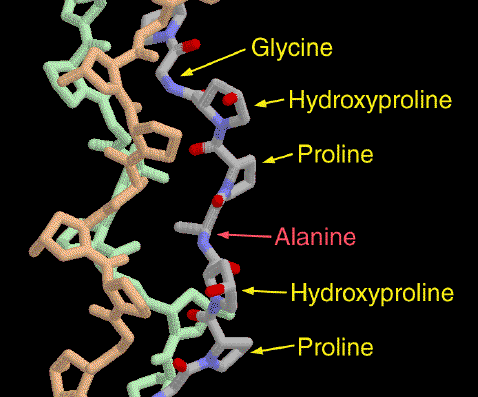
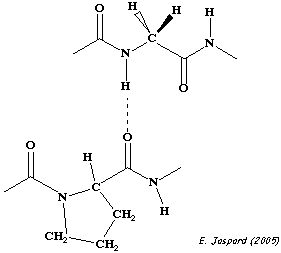
Source : "Principes de Biochimie" Horton et al. (1994) Ed. DeBoeck Universités Le collagène est caractérisé par le motif général : Gly - X - Y - Gly. En conséquence, chacune des chaînes portent une glycine tous les 3 résidus (30% des résidus). En effet, sa petite chaîne latérale (H) peut s'insinuer dans les espaces interhélicaux. Plus spécifiquement, on trouve les 2 motifs : Gly - Pro - X et Gly - X - Hyp. Pro et Hyp représentent 25% des résidus. Figure ci-dessous : liaison hydrogène intercaténaire entre l'hydrogène amide d'une glycine de la chaîne centrale de l'hélice triple du collagène et l'oxygène carbonyle des Pro d'une chaîne adjacente.
Figure ci-dessous : formation de la 4-hydroxyproline (Hyp). Il s'agit de l'hydroxylation de résidus proline par la prolyl-hydroxylase.
L'un des 2 atomes de l'oxygène (préalablement activé) s'attache à la proline, l'autre est incorporé au succinate lors de la décarboxylation de l'α-cétoglutarate. L'ascorbate (forme ionisée de la vitamine C) sert d'agent réducteur pour cette modification post-traductionnelle. Les étapes de maturation sont fondamentales pour la formation du procollagène et un déficit en ascorbate est léthal (scorbut des anciens marins - pas d'activité prolyl-hydroxylase). |
|
Une partie substantielle du volume des tissus s'appelle l'espace extracellulaire, qui est largement rempli par un réseau complexe de macromolécules constituant la matrice extracellulaire (MEC). La MEC se compose de 3 grandes classes de macromolécules :
Dans la plupart des tissus conjonctifs, les constituants de la MEC sont sécrétés principalement par les fibroblastes. Cependant, dans certains types spécialisés de tissus conjonctifs tels que le cartilage et l'os, les constituants de la MEC sont sécrétés par des chondroblastes et des ostéoblastes, respectivement. La collagénase 1 ("Matrix MetalloProteinase-1" - MMP-1 / EC 3.4.24.7)est synthétisée par une variété de cellules épithéliales et mésenchymateuses humaines (exemples : les kératinocytes, les fibroblastes et les macrophages). Elle hydrolyse le collagène de types I, II et III et la gélatine de types I, II, III, VI, VIII et X. Elle hydrolyse le collagène de type III plus rapidement. La collagénase 2 hydrolyse le collagène de types I et III. Elle hydrolyse le type I plus rapidement. Elle est synthétisée uniquement dans les granules spécifiques de cellules de neutrophiles polymorphonucléaires. La gélatinase A (MMP-2 / EC 3.4.24.24) : hydrolyse du collagène de types IV, V, VII et X et de la gélatine de type I. La gélatinase B (EC 3.4.24.35) : hydrolyse du collagène de types IV et V et de la gélatine de types I et V. Elle est synthétisée produite par des éosinophiles, des macrophages, des kératinocytes et les granules de neutrophiles. Les stromelysines [stromélysine1 (MMP-3), stromélysine2 (MMP-10) et stromélysine3 (MMP-11)] dégradent les protéoglycanes, la membrane basale, la laminine et la fibronectine en plus des collagènes. La matrilysine (MMP-7) et la metalloelastase (MMP-12). |
La bactérie anaérobie hautement pathogène, Clostridium perfringens, sécrète une enzyme qui hydrolyse la liaison peptidique suivante : X - Gly - Pro - Y ---> X-COO- + 3+HN-Gly - Pro - Y a. Comment la sécrétion de cette enzyme contribue-t-elle au caractère invasif de cette bactérie dans les tissus humains ? a. La protéine dont la liaison peptidique est hydrolysée est le collagène. b. L'enzyme sécrétée est donc une collagénase (EC 3.4.24.3). Les bactéries étant dépourvues de collagène, elles ne sont donc pas affectées par la collagénase. La collagénase amplifie le phénomène en favorisant la propagation des germes bactériens dans les tissus. Clostridium perfringens est une bactérie Gram+. Elle produit une toxine alpha (une lécithinase) impliquée dans des cas de gangrène (exemple : gangrène gazeuse) en anaérobiose dans les tissus. |
| 10. Liens Internet et références bibliographiques |
|
Online Mendelian Inheritance in Man "Collagen : The Fibrous Proteins of the Matrix" Molecular Cell Biology (4th ed.) Bone Biology and Mechanics Lab - Indiana University |
|
|
Bateman et al. (2009) "Genetic diseases of connective tissues: cellular and extracellular effects of ECM mutations" Nature Rev. Genet. 10, 173 - 183 Beuvelot et al. & Libouban (2009) "In vitro kinetic study of growth and mineralization of osteoblast-like cells (Saos-2) on titanium surface coated with a RGD functionalized bisphosphonate" J. Biomed. Mater. Res. B. Appl. Biomater. 90, 873 - 881 Ricard-Blum S. (2011) "The collagen family" Cold Spring Harb. Perspect. Biol. 3, a004978 Bertassoni et al. (2012) "The dentin organic matrix – limitations of restorative dentistry hidden on the nanometer scale" Acta Biomater. 8, 2419 - 2433 |
|
|
Yamauchi & Sricholpech (2012) "Lysine post-translational modifications of collagen" Essays Biochem. 52, 113 - 133 Fang & Banaszak Holl (2013) "Variation in type I collagen fibril nanomorphology: the significance and origin" BoneKEy Reports 2, 394 Mouw et al. (2014) "Extracellular matrix assembly: a multiscale deconstruction" Nat. Rev. Mol. Cell Biol. 15, 771 - 785 |
|
Vadon-Le Goff et al. (2015) "BMP-1/tolloid-like proteinases synchronize matrix assembly with growth factor activation to promote morphogenesis and tissue remodeling" Matrix Biol. 44-46, 14 - 23 Andriotis et al. (2015) "Structure–mechanics relationships of collagen fibrils in the osteogenesis imperfecta mouse model" J. R. Soc. Interface 12, 20150701 Bekhouche & Colige (2015) "The procollagen N-proteinases ADAMTS2, 3 and 14 in pathophysiology" Matrix Biol. 44 - 46, 46 - 53 Sherman et al. (2015) "The materials science of collagen" J. Mech. Behav. Biomed. Mater. 52, 22 - 50 Libouban, H. & Chappard, D. (2017) "Altered bone microarchitecture and gene expression profile due to calcium deficiency in a mouse model of myeloma" Micron. 96, 77 - 85 |
![]()