| Les structures des protéines - Introduction |
| Tweet |
|
|
1. Introduction - Protéines "fibreuses" et protéines "globulaires" 2. La liaison peptidique |
3. Les angles de torsion 4. Les diagrammes de Ramachandran 5. Liens Internet et références bibliographiques |
1. Introduction - Les 3 grands types de protéines α. Les protéines fibreuses : elles sont insolubles dans l'eau et constituent la charpente de la cellule. Les fibres protéiques sont constituées de la répétition d'éléments simples qui s'associent en "fils". Ce sont des protéines allongées dont les éléments de structure secondaire sont les structures dominantes. Exemple : la kératine (cheveux, ongles); le collagène (peau, os, ...). β. Les protéines membranaires :
γ. Les protéines globulaires : elles ont une structure tridimensionnelle assimilable à une sphère (un "globule"). On les trouve essentiellement dans le cytosol et les fluides cellulaires.
Voir un cours sur la détermination et l'analyse de la structure des protéines. |
|
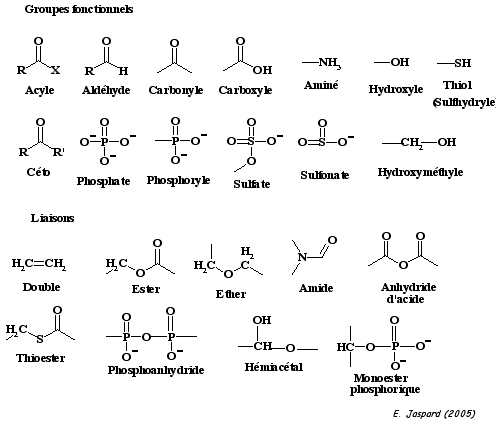
Les protéines sont des bioplolymères formés par la condensation d'éléments de base : les acides aminés. Tous les acides aminés comportent donc au moins un groupe aminé et un groupe carboxylique. Les réactions biochimiques modifient des liaisons chimiques ou des régions particulières d'une macromolécule biologique. Ces sites "réactifs" ou groupes fonctionnels sont de différents types. Figure ci-dessous : groupes fonctionnels et les liaisons dans les macromolécules biologiques.
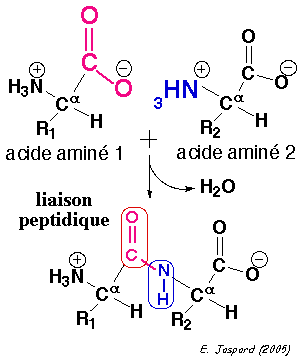
La liaison qui unit 2 acides aminés consécutifs s'appelle la liaison peptidique. Les 2 acides aminés sont alors appelés résidus d'acide aminé.
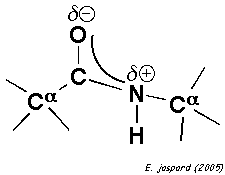
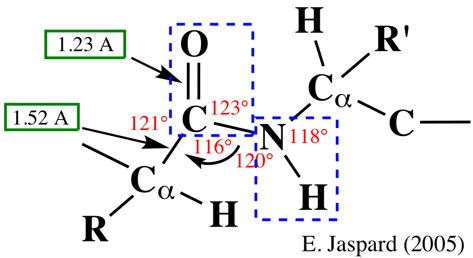
Comme l'oxygène est plus électronégatif que l'azote, les électrons délocalisés de la liaison peptidique sont plus proches de l'oxygène : la liaison peptidique est donc polaire. L'oxygène carbonyle porte une charge partielle négative et l'azote une charge partielle positive : tous deux peuvent former une liaison hydrogène. La liaison du carbone carbonyle avec l'azote dans la liaison peptidique (1,33 Å, non indiquée dans la figure) est plus courte que la liaison simple C-N mais plus longue qu'une liaison double C=N classique.
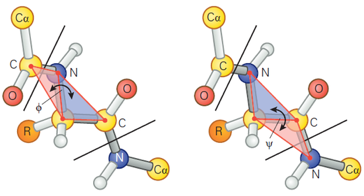
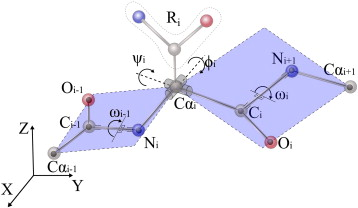
Le caractère partiellement double de la liaison peptidique empêche la rotation autour de la liaison C-N. En conséquence, le groupe peptidique est confiné dans un plan. Il existe cependant une liberté de rotation autour des liaisons Cα-C et N-Cα. |
|
La structure secondaire de la liaison peptidique est déterminée par 3 angles de torsion :
Source : Kuriyan et al. (2013) La liaison peptidique à le caractère d'une liaison double partielle (40%). Elle est presque exclusivement dans la conformation trans dans les protéines car cette conformation est plus favorable stériquement. En conséquence, l'angle ω est fixe et l'aire entre 2 carbones a consécutifs peut être considérée comme une unité peptidique planaire et rigide (figure ci-dessus).
Parfois l'angle dihédral est utilisé à la place de l'angle de torsion. Il est défini comme l'angle de torsion +180° : Φdihédral = Φtorsion + 180°. |
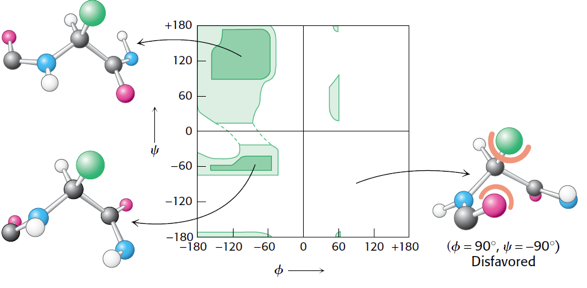
4. Les diagrammes de Ramachandran Dans la structure des protéines, toutes les valeurs des angles Ψ et Φ ne sont pas possibles car certaines rapprochent trop les atomes des chaînes latérales les uns des autres : il en résulte une répulsion entre ces atomes due à l'interaction de van der Waals et cette situation est énergétiquement défavorables. Comme l'angle ω est fixe, la structure secondaire de la liaison peptidique peut, en pricipe, être décrite par les 2 autres angles Φ et Ψ.
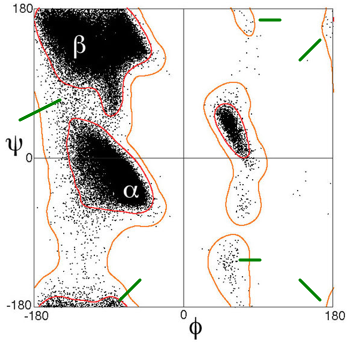
Sources : figure de gauche, Diez et al. (2013) - figure de droite, Kuriyan et al. (2013) Méthode du diagramme de Ramachandran Un tel diagramme permet de représenter sous forme d'aires, les différentes combinaison d'angles tolérés de la liaison peptidique.
Figure adaptée de : "Alpha helix (Wikipedia)" Dans la figure ci-dessus, on distingue différentes aires :
Visualiser un diagramme de Ramachandran à l'EBI : superoxide dismutase de Bos taurus (PDB 1E9O). Voir PDBsum / EBI pour d'autres diagrammes de Ramachandran. |
Un diagramme de Ramachandran peut être construit de 2 manières :
|
| liaison | limite normale | limite intérieure |
| C-C | 2.8 | 2.7 |
| C-O | 2.8 | 2.7 |
| C-N | 2.9 | 2.8 |
| C-H | 2.4 | 2.2 |
| O-O | 2.8 | 2.7 |
| N-N | 2.7 | 2.6 |
| H-H | 2.0 | 1.9 |
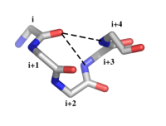
| Courts motifs (2 à 6 acides aminés) stabilisés par des liaisons hydrogène, trouvés dans les protéines | ||
| alpha-beta-motif | asx-motif | Exemple de définition : motif alpha-beta (figure ci-dessous)
Motif fréquent dans les hélices α C- et N-terminales. Les motifs α-β de pas gauche sont rares. Caractéristiques structurales :
|
| asx-turn-iL | asx-turn-iR | |
| asx-turn-iiL | asx-turn-iiR | |
| beta-bulge | ----- | |
| beta-bulge-loop-5 | beta-bulge-loop-6 | |
| beta-turn-iL | beta-turn-iR | |
| beta-turn-iiL | beta-turn-iiR | |
| gamma-turn-classic | gamma-turn-inverse | |
| nest-LR | nest-RL | |
| niche-3R | niche-3L | |
| niche-4L | niche-4L | |
| schellmann-loop-6 | schellmann-loop-7 | |
| st-staple | st-motif | |
| st-turn-iL | st-turn-iR | |
| st-turn-iiL | st-turn-iiR | |
| PDBeMotif: interface Web pour la recherche de motifs selon divers critères dans les protéines de la PDB. | ||
| 5. Liens Internet et références bibliographiques |
|
"Introduction à la structure des protéines" - C. Branden & J. Tooze (1996) - ed. De Boeck Université Kuriyan et al. (2013) "The molecules of life : physical and chemical principles" - ed. New York : Garland Science, Taylor & Francis Group |
|
Secondary structure and backbone conformation Motivated proteins : A Web Facility for Studying Small Hydrogen-Bonded Motifs Proteopedia : The free, collaborative 3D-encyclopedia of proteins & other molecules The Ramachandran Principle Phi (φ) and Psi (ψ) Angles in Proteins Base de données de structures cristallographiques "Protein Data Bank" |
|
|
Anfinsen et al. (1961) "The kinetics of formation of native ribonuclease during oxidation of the reduced polypeptide chain" Proc. Natl. Acad. Sci. USA 47, 1309 - 1314 Ramachandran et al. (1963) "Stereochemistry of polypeptide chain configurations" J. Mol. Biol. 7, 95 - 99 Ramachandran & Sassiekharan (1968) "Conformation of polypeptides and proteins" Adv. Prot.Chem. 28, p 283 - 437 Diez et al. (2013) "Biokinematic protein simulation by an adaptive dihedral angle approach" Mech. Mach. Theory 69, 105 - 114 |
|
![]()