
| Rôles des protéines LEA et des protéines de choc thermique (HSP) |
| Tweet |
|
|
1. Présentation générale 2. Aperçu de quelques protéines de stress 3. Aperçu de la réponse - tolérance des plantes aux stress biotiques et abiotiques 4. Quelques types de stress
|
5. Les HSP
6. Les chaperonnes moléculaires 7. Les protéines LEA
8. Liens Internet et références bibliographiques |
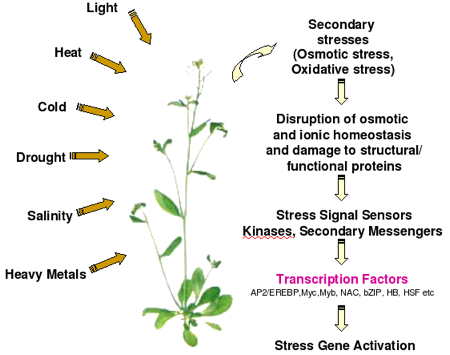
1. Présentation générale Tous les organsimes subissent des modifications/perturbations de leur conditions de vie, liées par exemple à des changements de leur environnement. Ces stress biotiques et abiotiques peuvent induire un dysfonctionnement cellulaire allant jusqu'à la mort des cellules. Différents stress (liste non exhaustive) :
Il existe des mécanismes cellulaires de défense contre cette atteinte à l'intégrité des cellules et /ou des protéines. Parmi les mécanismes de défense ou résistance ou tolérance aux stress des cellules, l'un des plus universels est une modification du profil d'expression des gènes codant des protéines spécialisées. En particulier, il existe une réponse appelée "réponse au choc thermique" (dénomination historique qui s'applique à tous les types de stress et pas seulement à la chaleur). Il en résulte une augmentation de la la synthèse de protéines particulières protectrices parmi lesquelles :
D'autres protéines de signalisation interviennent dans les mécanismes de régulation mis en place en réponse aux stress : par exemple la calmoduline ou les protéines 14.3.3. La résistance des plantes aux stress biotiques et abiotiques est un caractère multigénique qui dépend de la combinaison d'un grand ensemble de gènes, de protéines et de voies métaboliques qui agissent de concert. Les changements (tant qualitatifs que quantitatifs) au niveau des protéines en réponse aux stress, permettent une modulation des voies métaboliques et donc une meilleure protection de l'organisme. Ces éléments démontrent que les plantes ont développé des mécanismes cellulaires de réponse aux stress particulièrement flexibles qui leur permettent de s'adapter efficacement à ces stress. La tolérance des plantes aux stress peut être aussi acquise par une combinaision de diverses approches expérimentales :
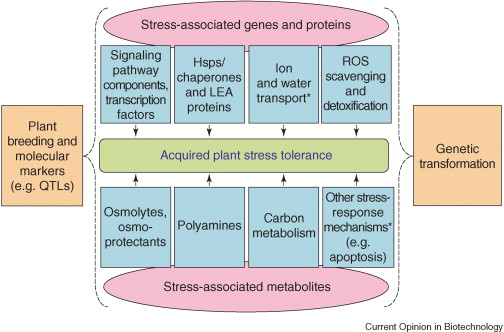
Source : Vinocur & Altman (2005) Légende de la figure ci-dessus :
|
| Les stress, de manière générale, et chez les plantes en particulier, sont des sujets de recherche actifs. | |
|
Publications scientifiques au NCBI Nombre d'articles PubMed (mai 2017) |
|
| Données d'expression des gènes (transcriptome) au TAIR | Microarray data from AtGenExpress : descriptions et résultats de toutes les expériences d'analyse des stress biotiques et abiotiques sur le transcriptome d'Arabidopsis par puces à ADN. |
| Bases de données |
|
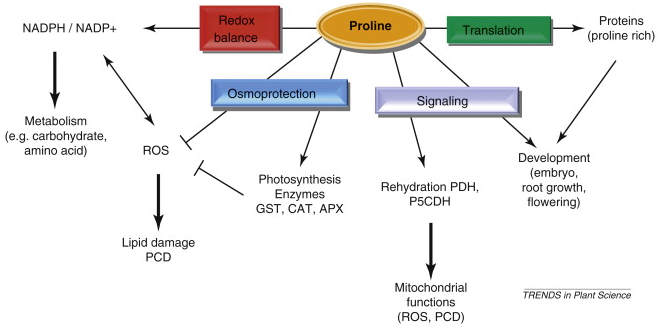
De nombreuses molécules autres que l'ADN (les gènes) et les protéines interviennent dans la régulation des différentes formes de réponses aux stress biotiques et abiotiques. En voici quelques exemples. a. Des osmoprotectants s'accumulent lors d'un stress lié à la sècheresse :
La proline a de trés nombreuses fonctions chez les plantes :
Légende de la figure ci-contre :
Source : Szabados & Savouré (2010)
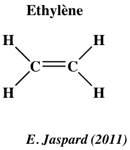
b. Une molécule extrêmement simple comme l'oxyde nitrique (NO) impliquée dans les réponses aux stress induits par la lumière, la gravité, les rayonnements UV-B, les conditions d'hypoxie, les blessures mécaniques, la température, les métaux lourds, les herbicides, la sècheresse, la salinité, ... c. L'éthylène (ou éthène) est l'alcène (hydrocarbure insaturé) le plus simple et c'est aussi un gaz.
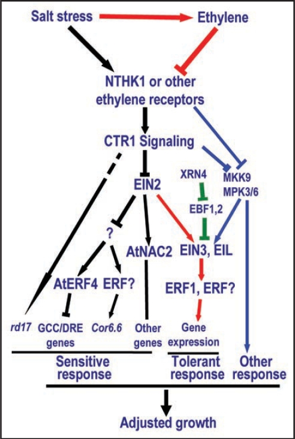
Cette simplicité structurale lui confère un trés grand nombre de fonctions dans les réponses aux stress mais également comme hormone végétale (implication dans la germination, les étapes de floraison, la maturation des fruits, la sénescence ...). L'acacia a développé une réponse particulièrement efficace au stress par blessure. L'éthylène (gaz) est relâché dans l'air et sert de moyen de communication entre les acacias pour signaler la présence d'herbivores (les koudous) : les acacias produisent alors des tanins mortels pour cette antilope. La voie de signalisation de l'éthylène module la réponse des plantes au stress salin à différents niveaux (récepteurs membranaires, composants cytoplasmiques et facteurs de transcription nucléaires). Les signaux sont perçus par la famille des récepteurs de l'éthylène, par exemple :
Source : Cao et al. (2008)
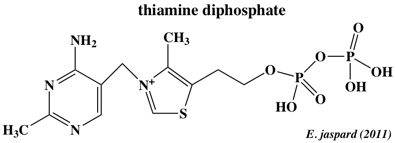
Ils contiennent un domaine N-terminal qui fixe l'éthylène (appelé aussi le domaine "senseur") et un domaine C-terminal conservé avec une activité Ser/Thr protéine kinase ou His protéine kinase. Ces signaux sont médiés par des modulateurs tels que : CTR1 ["(Mitogen-Activated Protein Kinase Kinase Kinase)-like protein Ctr1"] : Ser/Thr protéine kinase qui régulate négativement (répresseur) la voie de signalisation. EIN2 ("Ethylene INsensitive protein 2") : transporteur localisés dans la membrane du réticulum endoplasmique, régulateurs positifs de la réponse de l'éthylène. EIN3 / EIL : facteurs de transcription. Ce sont des activateurs (régulateurs positifs) de la transcription de ERF1 ("Ethylene Response Factor-1") et d'autres gènes liés à la réponse de l'éthylène. ERF1 appartient à la famille des facteurs de transcription APETALA2 / EREBP ("ethylene-responsive element binding proteins") spécifiques des plantes. Ils contiennent un domaine de fixation à l'ADN (boîte GCC présente dans de nombreux promoteurs des gènes inductibles par l'éthylène). d. La thiamine diphosphate (ou vitamine B1) a un rôle fondamental comme cofacteur enzymatique dans de nombreuses voies métaboliques (glycolyse, voie des pentose phosphates, pyruvate déshydrogénase, cycle de Krebs).
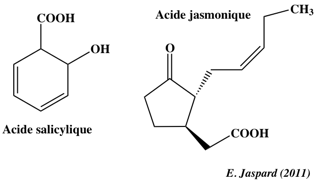
Cependant, il a été montré récemment qu'elle a d'autres fonctions et notamment qu'elle intervient dans la réponse des plantes aux stress biotiques et abiotiques. e. L'acide salicylique (acide 2-hydroxybenzoïque) intervient dans la défense des plantes contre les pathogènes. C'est aussi une phyto-hormone (transpiration, photosynthèse, transport d'ions).
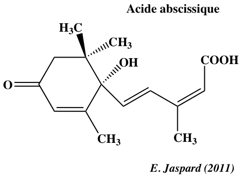
L'acide jasmonique intervient dans la réponse des plantes aux blessures (agressions par les herbivores, pathogènes, ...). L'acide jasmonique induit l'activation de facteurs de transcription de gènes codant des inhibiteurs de protéases qui se fixent aux enzymes digestives de certains prédateurs. f. L'acide abcissique ("abscisic acid" - ABA) a été identifié en 1963 par F. Addicott et ses collaborateurs, à partir des feuilles de cotonnier. C'est une hormone végétale synthétisée par les racines ou les feuilles (à l'intérieur des plastes).
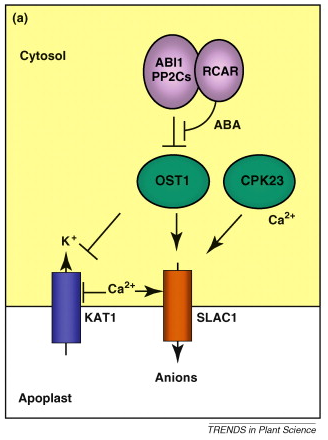
Structure : sesquiterpène - terpène dérivé de l'isoprène, composé en C15. Exemples de rôles de l'ABA L'ABA est un messager capital de nombreuses voies de transduction du signal en réponse à des stress biotiques et abiotiques (exemple : limiter le stress hydrique en période de sécheresse). Figure ci-contre, illustration du rôle de l'ABA dans l'ouverture de canaux ioniques.
Source : Raghavendra et al. (2010) Autres rôles physiologiques :
|
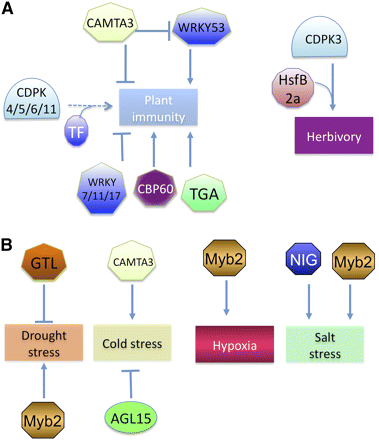
g. Le calcium et le complexe calcium/calmoduline (CaM) La figure ci-dessous illustre la régulation négative (indiquée par un segment horizontal) et positive (indiquée par des flèches) des stress biotiques (figure A) et abiotiques (figure B) par le groupe des facteurs de transcription qui fixent le Ca2+ ou le complexe Ca2+/calmoduline (Ca2+/CaM). Ligne pointillée : voie non encore complètement élucidée.
Source : Reddy et al. (2011) Cette figure montre :
Légende de la figure ci-dessus TF ("transcription factor") : facteur de transcription. CAMTA ("CAlModulin-binding Transcription Activator") : famille de facteurs de transcription activés par la calmoduline. WRKY : famille de facteurs de transcription ainsi nommés parce qu'ils possèdent un domaine de fixation à l'ADN (du côté N-terminal) qui contient, une ou deux fois, la séquence en acides aminés (quasi invariante) WRKY. CDPK : protéines kinases calcium dépendantes spécifiques des plantes. HsfB ("heat shock transcription factor") : famille de facteurs de transcription trimériques avec un domaine de fixation à l'ADN qui reconnaît la séquence répétée (nGAAn) et un domaine impliqué dans l'oligomérisation. Ils sont impliqués dans la réponse dite "de choc thermique" et la synthèse d'HSP ("heat shock proteins"). Il existe 3 classes (A, B et C). NIG ("NaCl - Inducible Gene 1") : facteur de transcription qui contient un motif "EF-hand". Ce facteur de transcription se fixe à la séquence CANNTG ("canonical E-box element") trouvée dans le promoteur d'un grand nombre de gènes impliqués dans la réponse au stress salin. MYB2 : famille de facteurs de transcription impliqués dans la réponse au stress salin et hydrique. CBP60 ("calmodulin-binding protein 60") : Il existe plusieurs isoformes (CBP60a à CBP60g). CBP60g est un facteur de transcription qui intervient dans la production d'acide salicylique en réponse aux MAMPs ("microbe-associated molecular patterns" - exemple : flg22, un fragment de la flagelline bactérienne). TGA ("TGACG motif-binding factor") : un membre de la famille de facteurs de transcription bZIP ("basic leucine zipper"), impliqué dans la résistance systémique acquise, dépendante de l'acide salicylique, des plantes, en réponse aux pathogènes biotrophiques. GTL ("GT element binding proteins") : famille de facteurs de transcription qui fixent des régions de l'ADN dites "GT element" (séquence GGTTAA). Les "GT elements" sont des éléments agissant en cis, trés dégénérées, avec des séquences riches en A/T. AGL15 ("AGAMOUS-like 15") : membre de la famille de facteurs de transcription qui contiennent un domaine N-terminal "MADS". AGL15 est préferentiellement exprimé dans les cellules embryonnaires de plantes. L’acronyme "MADS" vient de « minichromosome maintenance1 » (MCM1, Saccharomyces cerevisiae), « Agamous » (Arabidopsis thaliana), « Deficiens » (Antirrhinum majus) et « serum response factor » (SRF, Homo sapiens). L'analyse des facteurs de transcription et de leur sites de fixation à l'ADN bénéficie de méthodes récentes telles que l'immunoprécipitation de la chromatine ("Chromatin immunoprecipitation") couplée à l'analyse du transcriptome par une méthode massivement parallèle telle que "RNA-seq". |
2. Aperçu de quelques protéines de stress
La protéine chaperonne GRP78 (ou Bip) est impliquée dans la réponse au stress lié à l'accumulation de protéines mal repliées dans le réticulum endoplasmique ("Unfolded Protein Response" - UPR). |
| Stress | protéines sur-exprimées ou voies induites | protéines sous-exprimées ou voies réprimées |
| froid |
|
|
| châleur |
|
Riz : diminution de la RuBisCO |
| sécheresse |
|
protéines de signalisation (protéine "homoeobox / leucine zipper", facteur de transcription AP2/EREBP, ...) |
| salinité |
|
|
| ozone | nécrose des feuilles : protéines d'élimination des espèces radicalaires activées de l'oxygène (ROS) | chlorose des feuilles :
|
| hypoxie / anoxie |
|
biosynthèse des protéines (eIF-4A, eEF-2, EF-Tu mitochondrial) |
| Sous-illumination ou luminosité excessive |
|
--------------- |
| Excès de minéraux |
|
manganèse : assimilation du CO2 ("RuBisCO-binding protein", RuBisCO activase, PRK), sous-unité β6 du protéasome, "pathogenesis-related proteins" |
| Déficience en minéraux |
|
--------------- |
| Source : Kosovà et al. (2011) | ||
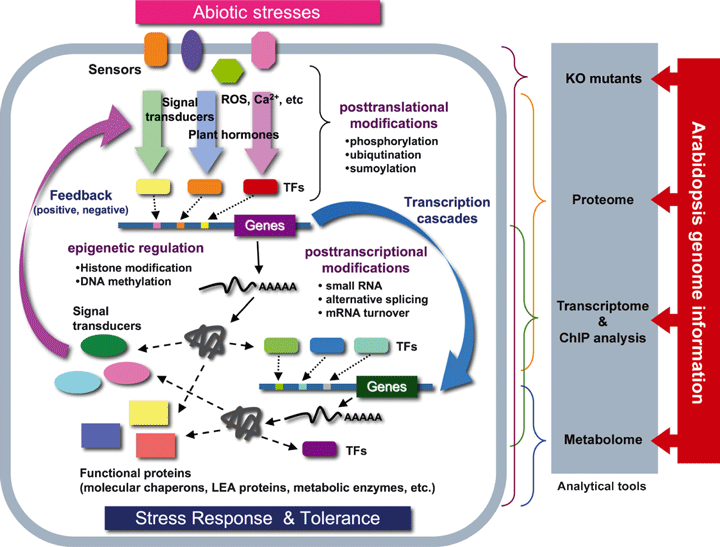
3. Aperçu de la réponse - tolérance des plantes aux stress biotiques et abiotiques Le décryptage de plus en plus complet et correctement annoté du génome d'Arabidopsis a considérablement contribué à l'élucidation du modèle général des réponses aux stress abiotiques chez les plantes (figure ci-dessous). Par ailleurs, les disciplines telles la génomique, la transcriptomique, la protéomique et la métabolomique permettent d'étudier au cours de ces réponses :
Ces informations permettent de décrire la régulation de la transduction des signaux lors des réponses aux stress, le rôle des protéines de stress dans les mécanismes de défense qui sont mis en place et les interactions entre ces protéines. Les produits des gènes inductibles par les stress peuvent être classés en 2 groupes :
Par ailleurs, certains transducteurs des signaux, également inductibles par les stress, ont été identifiés. Modele général de la réponse aux stress abiotiques chez les plantes Les cellules des plantes reçoivent des signaux via maints "senseurs" (peu ou pas encore connus) et ces signaux sont transduits via de nombreuses voies de signalisation dans lesquelles interviennent des seconds messagers, des hormones, des transducteurs du signal et des régulateurs de la transcription. Les gènes inductibles par les stress sont directement régulés par de nombreux "signaux de stress". Certains gènes inductibles par les stress sont régulés par des facteurs de transcription ("TFs" dans la figure ci-dessous) dont la transcription est elle-même induite par les stimuli liés aux stress (cascade de transcription).
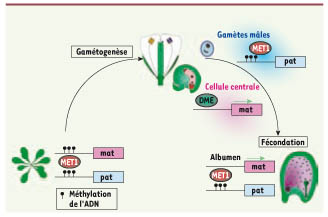
Source : Hirayama & Shinozaki (2010) Certains gènes inductibles par les stress codent des protéines dont la fonction est directement impliquée dans la tolérance aux stress. Certains gènes inductibles par les stress codent des protéines régulatrices (comme les transducteurs du signal) qui, trés probablement, sont impliquées dans des boucles de contrôle en amont ou des boucles de rétro-contrôle ("feedback positive / negative") de la réponse aux stress. L’épigénétique (figure ci-dessus) est un système de régulation fondamental, au-delà de l’information contenue dans la séquence d'ADN, puisqu'elle étudie les modifications, transmisssibles entre générations et réversibles, de l'expression des gènes sans qu'il y ait altération des séquences (modification et maintien de l’expression d’un gène sans modification de l’ADN). Un mécanisme possible de la régulation épigénétique est la méthylation de la cytosine en 5-methylcytosine (catalysée par la méthyltransférase de maintenance MET1 chez Arabidopsis) et la déméthylation de l’ADN dans les régions du promoteur (exemple : WRKY70) ou dans les régions de régulations d’un gène. Un exemple de régulation épigénétique pour la reproduction des plantes est l'empreinte génomique parentale qui affecte les gènes exprimés dans les tissus extra-embryonnaires responsables de l’acheminement des nutriments d’origine maternelle vers l’embryon (figure ci-dessous) .
Source : Jullien & Berger (2008) DME : la glycosylase DEMETER qui déméthyle l’ADN. |
Les facteurs de transcription impliqués dans la réponse aux stress Chez Arabidopsis, on dénombre plus de 2000 facteurs de transcription (> 7% du protéome). Ils font partie des 58 familles suivantes :
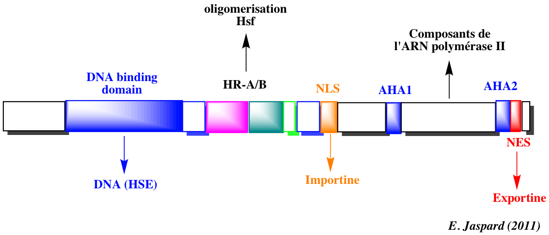
Source : Stress responsive TranscrIption Factor Database Etude de de la cinétique des effets de différents stress (ABA, dessication, froid, salinité) sur l'expression de génes d'Arabidopsis thaliana (analyse d'article). Quelques exemples de familles de facteurs de transcription - Famille AP2 (APETALA2) / EREBP ("ethylene-responsive element binding proteins") : ils sont spécifiques des plantes. Ils contiennent un domaine de fixation à l'ADN appelé AP2. La sous-famille de facteurs de transcription DREB ("dehydration responsive element binding proteins") fait partie de cette famille. - Famille bZIP ("basic leucine zipper") : leur nom vient du fait qu'ils contiennent un domaine de fixation à l'ADN mixte constitué d' une séquence en acides aminés basiques et d'un motif riche en leucine à intervalles réguliers ("leucine zipper"). Les facteurs de transcription GBF1, GBF2 et GBF3 ("Arabidopsis bZlP family of G-box binding factors") intéragissent avec le motif palindrome "G-box" (CCACGTGG) trouvés dans de nombreux promoteurs de plantes. - Famille HSF : ("heat shock transcription factor") : ils sont trimériques avec un domaine de fixation à l'ADN qui reconnaît la séquence répétée (nGAAn) et un domaine impliqué dans l'oligomérisation. Ils sont impliqués dans la réponse dite "de choc thermique" et la synthèse d'HSP ("heat shock proteins"). Il existe 3 classes (A, B et C). Ces facteurs de transcription sont organisés en modules fonctionnels comme le facteur de transcription AtHsfA2 de Arabidopsis :
- Famille WRKY : ils sont ainsi nommés parce qu'ils possèdent un domaine de fixation à l'ADN (du côté N-terminal) qui contient, une ou deux fois, la séquence en acides aminés (quasi invariante) WRKY. Ils sont classés en fonction du nombre de motif WRKY et de leur motif "zinc-finger-like" : Cx[4,5]Cx[22,23]HxH ou Cx7Cx23HxC. |
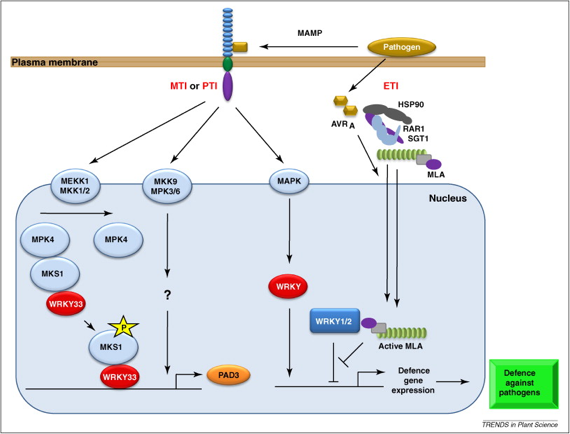
Schéma des voies de signalisation impliquant les facteurs de transcription de la famille WRKY en réponse aux attaques de certains pathogènes.
Source : Rushton et al. (2010) MAMP : "microbe-associated molecular patterns" |
|
Voie de gauche : Arabidopsis
WRKY33 augmente le niveau d'expression de PAD3, nécessaire à la synthèse de la camaléxine (antimicrobien). |
Voie de droite : Hordeum vulgare
La dé-répression des réponses de défense de base est sans doute cause d'une amplification du niveau d'expression des gènes liés aux mécanismes de défense à ces pathogènes. |
|
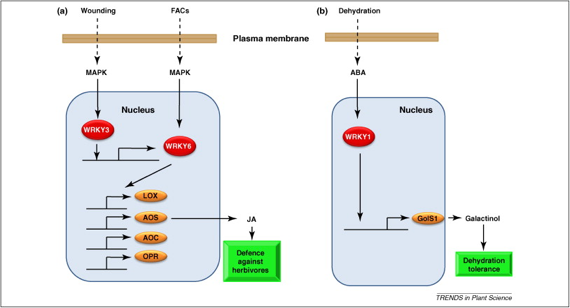
Schéma des voies de signalisation impliquant les facteurs de transcription de la famille WRKY en réponse aux attaques de certains herbivores et à la déshydratation
Source : Rushton et al. (2010) |
|
Voie de gauche : Nicotiana attenuata 2 facteurs de transcription (WRKY3 et 6) coordonnent la réponse en régulant l'expression des gènes de la voie de biosynthèse de l'acide jasmonique (gènes LOX, AOS, AOC et OPR).
|
Voie de droite : Boea hygrometrica ABA : acide abcissique |
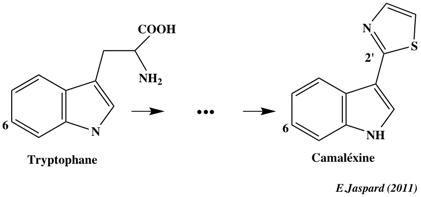
La camaléxine (3-thiazol-2′-yl-indole) est une phytoaléxine dérivée du tryptophane (via l'indole-3-acétaldoxime).
C'est un métabolite secondaire impliqué dans la défense contre certains pathogènes. Exemples de bases de données de facteurs de transcription spécifiques des plantes :
Chez les plantes, la plupart des gènes miRNA :
Les miRNAs sont inclus dans les pri-miRNA et clivés en 2 étapes :
On pense que les deux clivage ont lieu dans le noyau et sont catalysés par l'enzyme Dicer-like 1 (DCL1 qui génère des fragments de 21 nucléotides) chez Arabidopsis. Le duplex miRNA:miRNA* est déroulé et le miRNA (mais pas le miRNA*) est préférentiellement incorporé dans le complexe RISC. il joue le rôle d'ARN guide pour diriger la répression post-transcriptionnelle des cibles mRNA. Le miRNA* est dégradé. Arabidopsis possède 4 orthologues de Dicer (DCL1 à DCL4). Les miRNAs des plantes interviennent dans de nombreux processus : transitions lors du développement, croissance des feuilles, polarité des organes, voie de signalisation par l'auxine, métabolisme des ARN, réponses des plantes aux stress biotiques et abiotiques, ... |
|
a. Stress hydrique : déshydratation, déficit en eau, dessication, sècheresse ("dehydration, water deficit, dessication, drought") - Voir la revue : Timperio et al. Le contenu en eau dépend du type de tissu et du type de cellule. L'eau est nécessaire pour :
Le stress hydrique conduit invariablement à un ralentissement de la vitesse de la photosynthèse. La tolérance à ce stress varie cependant selon les espèces végétales. Parmi les facteurs responsables de cette diminution de la photosynthèse, la fermeture des stomates (liée à l'ABA) peut-être considérée comme une réponse directe à la réduction du potentiel hydrique des feuilles par la sècheresse. La réduction de la conductance des stomates limite les apports en CO2, ce qui diminue la concentration intercellulaire en CO2. L'assimilation nette en CO2 est de ce fait contrainte ce qui diminue la croissance de la plante et sa productivité (Chaves et al., 2002). La sècheresse pertube :
Les réponses cellulaires incluent :
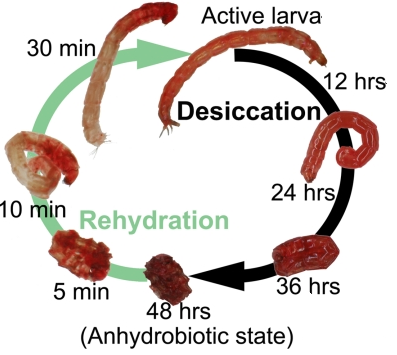
L'une des signatures de la tolérance à la dessication est l'accumulation d'HSP et de protéines LEA. Elles s'accumulent dans le cas de stress hydrique, salin, et de température élevée dans le but de protéger les semences, par exemple. Des invertebrés comme les nématodes (Aphelenchus avenae), les rotifères, les tardigrades, les chironomides (Polypedilum vanderplanki ) et les arthropodes (Artemia franciscana) sont capables de survivre à une déshydratation réversible jusqu'à 2% d'eau dans leurs tissus. On parle d'anhydrobiose. Figure ci-dessous : cycle d'anhydrobiose de la larve de Polypedilum vanderplanki (chironomide).
Source : Cornette et al. (2010) Le 6 mai 2011, des tardigrades ont été envoyés dans l'esapce (avec d'autres extremophiles) lors du dernier voyage de la navette spatiale "Endeavour". |
|
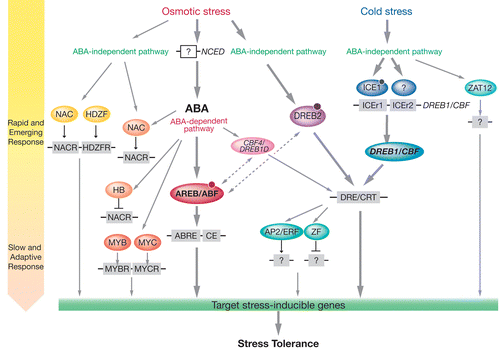
Froid et stress hydrique ont un lien : quand la température chute, les molécules d'eau se déplacent des cellules vers l'espace intercellulaire par osmose, ce qui induit un stress hydrique de la cellule. La déshydratation induite par le gel déstabilise les membranes plasmiques ce qui entraîne la formation de phases hexagonales inversées au niveau des structures membranaires. Un gel rapide peut entraîner des dégâts majeurs dûs à la formation de glace. La déshydratation entraîne l'agrégation des macromolécules biologiques, notamment des protéines, donc leur inactivation irréversible.
Source : Yamaguchi-Shinozaki & Shinozaki (2006)
Pour affronter les températures inférieures à 0°C, les plantes modifient leurs compositions enzymatiques et membranaires, modifiant en conséquence leur métabolisme pour synthétiser des cryoprotectants comme les polyols et les sucres (par exemples, le tréhalose ou le sucrose , le myo-inositol) afin d'abaisser le point de congélation de leurs tissus et lutter contre la dessiccation.
Ces osmolytes, outre leur rôle de remplacement de molécules d'eau, protègent également les membranes en maintenant leur fluidité. Les protéines LEA sont, dans leur grande majorité, extrêmement hydrophiles : leur accumulation en cas de stress hydrique sert de "tampon d'hydratation" en séquestrant les ions et en stabilisant les protéines et les membranes en interagisaant directement avec elles. Elle sont pour la plupart nativement non structurées. |
|
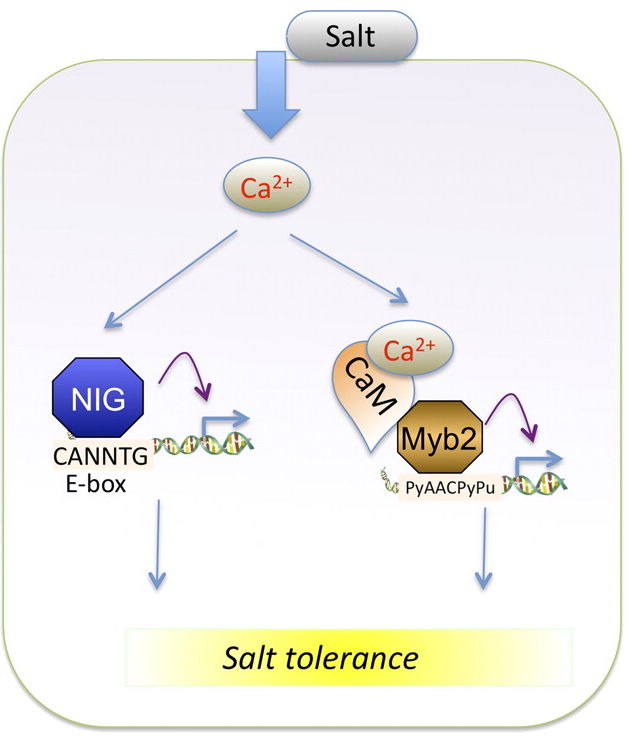
Ce stress est l'un des plus sévères pour les plante. L'aridité croissante de nombre de régions du monde y contribuant et s'y ajoutant, il affecte environ 23% des terres cultivées. Des concentrations élevées en sel dans le milieu environnant pénalisent les organismes car le potentiel hydrique est diminué. Or l'eau est absorbée par les cellules par osmose, elle même gouvernée par le gradient du potentiel hydrique. Une cellule peut absorber de l'eau quand elle a un potentiel hydrique inférieur à celui du milieu environnant. Une concentration en ion correspondant à [NaCl] 100 mM (équivalent à une concentration osmotique de 200 mOsmol/L) rend difficile, voire impossible, l'absorption de l'eau par une cellule (ou même de retenir cette eau) sans la mise en place de système d'acclimatation. Pour maintenir constante la concentration en eau intracellulaire, la concentration cytoplasmique de composés osmotiquement actifs doit être supérieure à celle du milieu environnant par l'absorption d'ions du milieu environnant (phénomène de "salt-in"). Exemple de micro-organismes hautement spécialisés (concentration cytoplasmique : 2 à 3 M KCl !). Cependant, de fortes concentrations de certains ions ont des effets toxiques sur le métabolisme cellulaire. En conséquence, les réponses au stress salin et celles au stress hydrique et au froid ont de nombreux points communs. L'une des signatures de la tolérance au stress salin est, une fois encore, l'accumulation d'HSP et de protéines LEA. Schéma de la régulation de l'expression de gènes impliqués dans la réponse au stress salin via des facteurs de transcription fixant le Ca2+ ou le complexe Ca2+/calmoduline (CaM).
Source : Reddy et al. (2011)
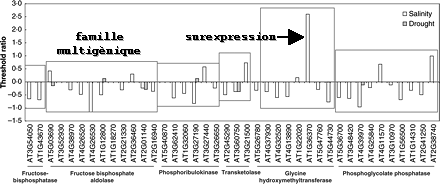
La calmoduline (CaM) se fixe à MYB2 en fonction de la concentration en Ca2+. La fixation de la CaM induit un changement de conformation de MYB2, ce qui augmente sa capacité à se fixer à sa séquence cible sur l'ADN. La conséquence est une activation de la transcription des gènes cibles de MYB2. Comparé aux effets de la sécheresse, le stress salin modifie la sur-expression ou la sous-expression d'un plus grand nombre de gènes avec une intensité plus élevée (figure ci-dessous).
Source : Chaves et al. (2009) Celà reflète sans doute les effets combinés de la salinité (déshydratation et choc osmotique). Au sein d'une famille multigénique, on observe que le même stress peut avoir des effets distincts sur différents gènes, tant sur le plan quantitatif que qualitatif. Par exemple, les deux gènes codant la fructose-bisphosphatase sont sous-transcrits de manière semblable par le stress salin. En revanche, les gènes codant des enzymes du cycle de Calvin et de la photorespiration (fructose-bisphosphate aldolase, phosphoribulokinase, transcétolase, ribose-5-phosphate isomérase, phosphoglycolate phosphatase, glycine hydroxyméthyltransférase) sont modifiés différemment. Celà souligne la trés grande aptitude des plantes à réagir et à répondre de manière différente à des conditions environnementales et de croissance distinctes. Celà souligne aussi l'importance capitale d'étudier l'incidence de la compartimentation des voies métaboliques sur la régulation de l'expression des gènes et sur la teneur en protéines codées par ces gènes et spécifiquement adressées. En toute rigueur, de tels résultats ne peuvent être pris en compte que s'ils sont confirmés par l'analyse quantitative et qualitative des protéines exprimées par ces gènes : c'est l'un des buts de la protéomique (gels 2D et spectromètrie de masse). |
|
Le basculement d'un métabolisme général aérobie (respiration) à un métabolisme général anaérobie (fermentation) permet un apport d'énergie à la cellule dans des conditions de concentration d'oxygène faible (hypoxie). L'éthylène joue un rôle central dans la signalisation de la réponse à l'hypoxie de même que les protéines G (transducteurs de ce signal). Il semble qu'il y ait un lien entre les espèces radicalaires activées de l'oxygène (ROS - "reactive oxygen species") et les HSP, suggérant qu'au cours de l'évolution, les plantes ont acquis des mécanismes sophistiqués pour contrôler la toxicité des ROS et qu'elles savent aussi les utiliser comme signaux pour l'induction des HSP. Un radical libre porte sur sa couche électronique externe un ou plusieurs électrons célibataires (non apparié à un électron de spin opposé). Cette configuration confère à l'entité radicalaire une trés haute réactivité chimique : les radicaux tendent à capturer un électron pour compléter leur couche électronique. Des études par double-hybrides (Elmayan & Simon-Plas, 2007) ont mis en évidence un rôle trés important des protéines 14.3.3 en réponse aux ROS. Ces protéines de signalisation interagissent avec plus de 300 cibles à ce jour identifiées : elles jouent en fait un rôle majeur dans les mécanismes mis en place en réponse à un trés grand nombre de stress : |
| Voie de signalisation | Protéines cibles des protéines 14.3.3 | Mode d'interaction |
| Acide gibberellique | Activateur de la transcription (bZIP) "Repression of shoot growth" | Changement de localisation sub-cellulaire |
| Acide abcissique | Régulateurs de la transcription "Abcissic acid insensitive" ABI3, ABI5 | Association de protéines cibles |
| Hormones stéroïdes | "leucine-rich repeat (LRR) receptor kinase" BRI1 Répresseur de la transcription BZR1 |
Changement de localisation sub-cellulaire |
| lumière bleue | Phototropines 1(protéines kinases autophosphorylantes) | Inconnu |
| lumière rouge | CO | Inconnu |
| "Hypersensitive response" | NADPH oxydase | Changement d'activité (activation / inhibition) |
| MAPKKKα ("mitogen-activated protein kinase kinase kinase") | Stabilisation de la forme active |
α. Energie des rayonnements ionisants Les radicaux libres proviennent essentiellement de l'interaction des rayonnements ionisants avec les électrons des molécules d'eau, et ce en raison de la teneur extrèmement élevée en eau des organismes vivants. 1er cas : 2ème cas : Finalement : 10-10 secondes à 10-9 secondes aprés l'interaction avec le rayonnement ionisant, l'eau est devenue une solution plus ou moins concentrée en radicaux OH° et H° et en molécules d'hydrogène issues de la réaction : H° + H° ---> H2 β. Le rôle du transfert linéique d'énergie (T.L.E.) Les réactions qui succèdent à la formation de radicaux libres surviennent dans un délai de 10-7 secondes à 10-3 secondes aprés l'interaction du rayonnement ionisant et constituent l'étape diffusionnelle. Ces réactions dépendent du transfert linéique d'énergie du rayonnement. 1er cas : le T.L.E. est élevé, donc le nombre
d'ionisation est grand, donc la probabilité de rencontre entre
les radicaux libres formés est élevée : 2ème cas : le T.L.E. est faible et la réaction de recombinaison est la plus probable : H° + OH° ---> H2O γ. L'influence de la présence d'oxygène Les réactions qui succèdent à la formation de radicaux libres dépendent également de la présence de l'oxygène: 1er cas : en absence d'oxygène, la réaction
s'arrète par dimérisation : H° + H° ---> H2 2ème cas : en présence d'oxygène, la réaction
suivante à lieu : H° + O2 ---> OOH° Le radical hydroperoxyde, OOH°, est un oxydant
avide d'électrons. Il est à l'origine de la formation
d'eau oxygénée : L'électron arraché de la molécule d'eau ionisée lors de l'interaction avec le rayonnement ionisant peut réagir avec l'oxygène et former le radical superoxyde, O2°-: e- + O2 ---> O2°- δ. L'influence de la présence de molécules organiques Les réactions qui succèdent à la formation de radicaux libres dépendent enfin de la présence de molécules organiques. Un grand nombre de réactions (voir ci-après) ont lieu et aboutissent à la formation :
Les peroxydes et les tétroxydes sont des oxydants puissants qui altèrent les lipides des membranes des cellules. Ces réactions sont :
OOH° + RH ---> HOOH + R° ε. L'inactivation des radicaux libres Dans toutes cellules normales il existe des réactions qui produisent des radicaux libres et des peroxydes similaires à ceux formés en cas de radioexposition. Les cellules possèdent donc des enzymes dont la fonction est d'inactiver ces composés extrèmement toxiques: - la superoxyde
dismutase :
2 O2°- + 2 H+ ---> H2O2 + O2 Remarque : la réaction catalysée par la superoxyde dismutase est équilibrée en terme de radical (symbole °) puisque le transfert électronique met en jeu un atome de cuivre lié à l'enzyme qui est successivement oxydé puis réduit. Voir un cours sur les effets biologiques des rayonnements ionisants. |
|
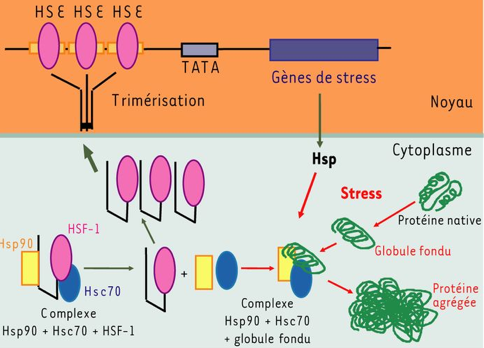
a. Généralités La réponse de choc thermique a été découverte chez des drosophiles exposées à une température sublétale. En 1962, F.M. Ritossa remarqua que la châleur (accidentellement élevée) et le découplant dinitrophénol induisaient une augmentation de l'expression d'une protéine chez la Drosophile (Ritossa F.M. , 1962 & 1964). Les gènes eucaryotes codant pour les HSP possèdent un promoteur particulier (HSE - "heat shock element"), contenant au moins trois motifs nGAAn en répétition inversée.
Source : HSPIR database Les différents agents ou conditions induisant la réponse aux stress agissent sur le facteur de transcription HSF-1 ("heat shock transcription factor-1"), présent et inactif dans le cytoplasme (figure ci-dessous).
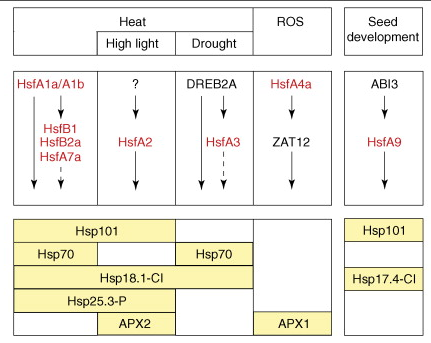
Source : Arrigo (2005) Après une étape de phosphorylation et de trimérisation, HSF-1 migre dans le noyau, se lie aux sites HSE sur l'ADN et active la transcription des gènes codant pour les HSP. Rôle de différents facteurs de transcription de la famille HSF dans diverses voies de transduction du signal. Le modèle représente une synthèse des résultats obtenus avec des mutants d'Arabidopsis portant sur les facteurs de transcription HsfA1a, HsfA1b, HsfA2, HsfA4a, HsfA9, DREB2A, APX1 et ABI3 Les différents signaux/stimuli (en haut de la figure ci-dessous) induisent des cascades transcriptionnelles distinctes qui impliquent des facteurs de transcription de la famille HSF (en rouge), qui agissent directement ou indirectement sur le niveau d'expression de leurs gènes cibles (rectangles jaunes en bas).
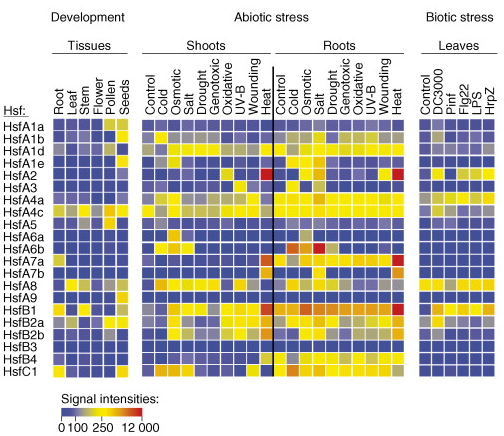
Source : Koskull-Döring et al. (2007) APX : ascorbate peroxidase Expression des différents facteurs de transcription de la famille HSF dans diverses conditions de stress Représentation sous forme de "heat-map" (figure ci-dessous) des intensités des signaux relatifs et normalisés traduisant les profils d'expression des transcrits des gènes qui codent les facteurs de transcription d'Arabidopsis.
Source : Koskull-Döring et al. (2007) Ces données de puces à ADN sont extraites de AtGenExpress. Le code couleur des intensités est présenté en bas de la figure selon la représentation du logiciel "GeneSpring 7.2". |
| Conditions expérimentales des stress testés (figure ci-dessus) | ||
|
Echantillons "stade de développement" :
|
Echantillons "stress abiotiques" : semences de 18 jours poussées en conditions hydroponiques.
|
Echantillons "stress biotiques" : feuilles analysées au bout de 24h :
|
AtGenExpress est un consortium qui regroupe les données de transcriptomique de Arabidopsis obtenues par puces à ADN à différents stades de développement et pour de trés nombreuses conditions de stres biotiques et abiotiques. Cette base de données implémentée au TAIR renvoit vers de nombreuses autres bases de données. b. Caractéristiques et fonction(s) des "small HSP" (sHSP) Les sHSP ont été mises en évidence chez quasiment tous les organismes, mais elles semblent plus particulièrement liées aux plantes, chez lesquelles elles représentent la classe majeure d'HSP (Vierling, 1991). Par ailleurs, les sHSP fonctionnent selon un mécanisme qui est indépendant de l'hydrolyse de l'ATP, contrairement aux autres membres de cette famille, notamment les HSP 70 (Waters et al., 1996). Cependant, la (ou les) fonction(s) biologique(s) exacte(s) des sHSP n'est (ne sont) pas encore clairement établie(s). C'est chez le pois que l'on a mis pour la première fois en évidence une de leurs propriétés fonctionnelles : les deux sHSP cytosoliques empêchent l'agrégation de protéines dénaturées artificiellement (Lee et al., 1995). En conséquence, puisque :
il a été proposé que les sHSP mitochondriales, et notamment l'HSP22, jouent un rôle clé dans le maintien du métabolisme aérobie en conditions de stress (Lund et al., 1998). c. L'HSP22 de mitochondries de feuilles de pois (Pisum sativum) α. La séquence nucléotidique du précurseur et de la forme mature de l'HSP22 La séquence qui code le précurseur de l'HSP22 de mitochondries de feuilles de pois présente les caractéristiques suivantes :
β. La séquence polypeptidique du précurseur et de la forme mature de l'HSP22 L'HSP22 précurseur est une protéine de 202 acides aminés d'une masse molaire apparente de 22,9 kDa, dont les caractéristiques sont les suivantes :
La protéine sur-exprimée (la forme mature de l'HSP22) contient 170 acides aminés pour une masse molaire prédite de 19,5 kDa. L'extrémité N-terminale de la protéine purifiée à partir des mitochondries est l'Asn 33 (Lenne & Douce, 1994). γ. Alignement de séquences d'HSP22 de mitochondries L'extrémité N-terminale de la protéine recombinante purifiée a été séquencée. Du fait de la construction dans le vecteur et donc la présence du codon d'initiation de la traduction (ATG), il y a une méthionine additionnelle avant l'Asn 33 (premier acide aminé de la forme mature de l'HSP22). |
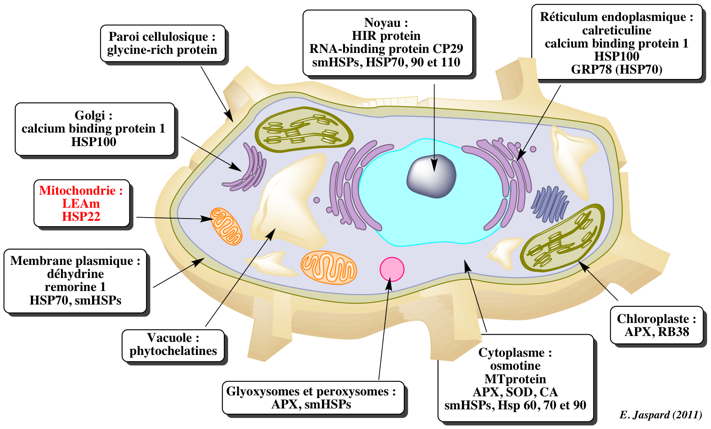
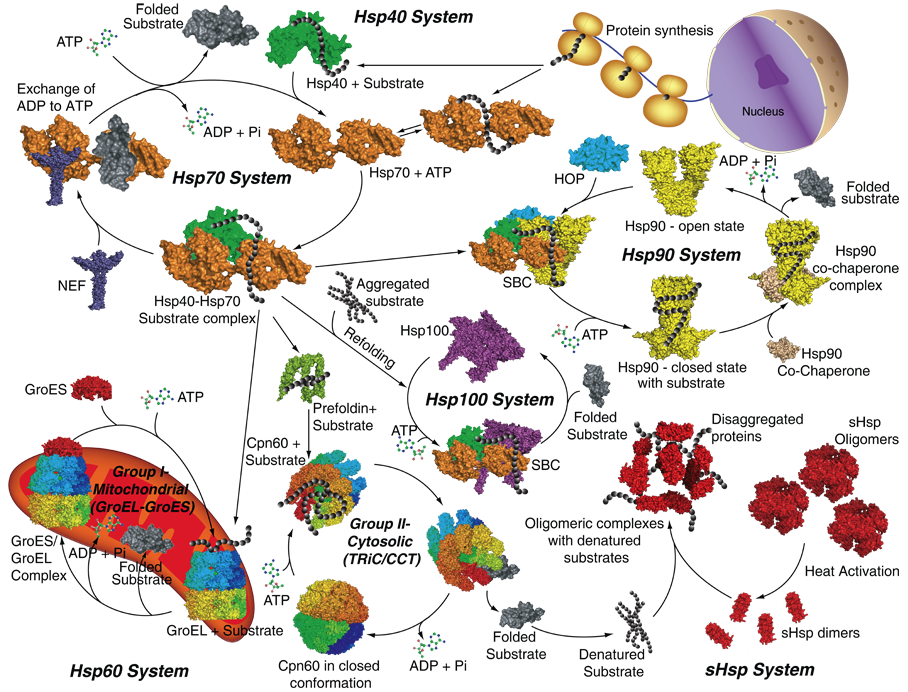
6. Les chaperonnes moléculaires Dans la cellule, les conditions physiologiques ne permettent pas à toutes les chaînes polypeptidiques de se replier sans assistance. Il existe un groupe de protéines dont le rôle est d'aider certaines autres protéines à se replier, et c'est pour cette raison qu'elles ont été dénommées "chaperonnes moléculaires" (Ellis, 1987). Les chaperonnes existent dans toutes les cellules (eucaryotes et procaryotes) quelles que soient les conditions, mais on ne les a mises en évidence qu'à la suite d'une modification temporaire de la température optimale pour l'existence de la levure. Il y a donc une synthèse accrue des chaperonnes moléculaires après un choc thermique, mais également après d'autres types de stress comme le stress oxydatif, le déficit en eau ou l'élévation de salinité. Le froid induit la synthèse de protéines appelées " Cold Induced Protein ", en nombre plus restreint et de masse molaire plus élevée. Cette profonde altération du profil protéique d'un organisme en réponse à un stress (c'est-à-dire le type de chaperonne(s) moléculaire(s) nouvellement synthétisée(s) ou sur-exprimée(s)), est spécifique non seulement de l'organisme considéré mais également, au sein d'une cellule, des compartiments qui la composent. La famille des chaperonnes moléculaires se divise en deux sous-familles de protéines, distinctes tant du point de vue de leur structure quaternaire que de leur mécanisme d'action (Gething & Sambrook, 1992 ; Ellis, 1994 ; Clarke, 1996 ; Braig, 1998 ; Ellis & Hartl, 1999) :
Voir un tableau qui résume les dénominations, localisations et caractéristiques des chaperonnes. |
|
Les protéines LEA ont été mises en évidence dans la phase tardive de l'embryogénèse de semences (d'où leur nom) du coton (Gossypium hirsutum) (Dure, Galau et al.). Leur synthèse et leur accumulation est corrélée à divers types de stress, notamment le stress hydrique (déshydratation, dessication), le stress par le froid et le stress salin. Bien qu'initialement mises en évidence chez les plantes, on en découvre cependant de plus en plus dans d'autres types d'organismes, notamment les organismes anhydrobiotes. Par exemple, des invertebrés comme les nématodes (Aphelenchus avenae), les rotifères, les tardigrades, les chironomides (Polypedilum vanderplanki ) et les arthropodes (Artemia franciscana) qui sont capables de survivre à une déshydratation réversible jusqu'à 2% d'eau dans leurs tissus. Les protéines LEA forment une famille réparties en 12 classes qui correspondent à 8 PFAM. Ces classes se distinguent par l'utilisation des acides aminées et les propriétés physico-chimiques très différentes des protéines qui en découlent. A l'exception de la classe 7, les protéines LEA n'ont pas de structure tri-dimensionnelle établie à l'état natif dans des conditions normales d'hydratation. Les protéines LEA sont donc pour la plupart des protéines dites intrinsèquement désordonnées ("intrinsically disordered proteins") ou nativement non structurées ("natively unstructured proteins"). Cette absence de structure tridimensionnelle connue et résolue pose un problème :
|
| Evolution de la classification des protéines LEA | |||||||
| PFAM (name) | Dure et al. (1989) | Bray (1993) | Tunnacliffe & Wise (2007) | Battaglia et al. (2008) | Bies-Esthève et al. (2008) | Hundertmark & Hincha (2008) | LEAPdb Hunault & Jaspard (2010) |
| PF00257 (dehydrin) | D11 | Group 2 | Group 2 | Group 2 | Group 2 | dehydrin | classes 1 to 4 |
| PF04927 (SMPb) | D34 | Group 6 | Group 6 | Group 5A | Group 5 | SMP | class 11 |
| PF03760 (LEA_1) | --- | Group 4 | Group 4 | Group 4A | Group 4 | LEA_1 | class 10 |
| D113 | Group 4B | --- | |||||
| PF03168 (LEA_2) | D95 | --- | --- | Group 5C | Group 7 | LEA_2 | classes 7 & 8 |
| PF03242 (LEA_3) | D73 | --- | Lea5 | Group 5B | Group 6 | LEA_3 | class 9 |
| PF02987 (LEA_4) | D7 | Group 3 | Group 3 | Group 3A | Group 6 | LEA_4 | class 6 |
| D29 | Group 5 | Group 3B | --- | ||||
| PF00477 (LEA_5) | D19 | Group 1 | Group 1 | Group 1 | Group 1 | LEA_5 | class 5 |
| D132 | |||||||
| PF10714 (LEA_6) | --- | --- | --- | Group 6 | Group 8 | PvLEA18 | class 12 |
| PF10714 (ABA-WDS) | --- | --- | --- | Group 7 | --- | --- | --- |
|
|||||||
Les protéines LEA et la tolérance à la déshydratation / dessication Leurs fonctions sont encore trés peu connues et encore moins leur mécanisme d'action. Malgré tout, leurs rôles les mieux documentés est l'acquisition d'une tolérance à la dessication par protection des membranes à l'état sec, la séquestration d'ions et la protection de certaines protéines contre la dénaturation. Voir un article qui décrit l'acquisition de la tolérance à la déshydratation de plants de riz transgéniques (transgert du gène HVA1 du blé). Dans une cellule à l'état normal, les protéines LEA n'ont pas de structure tridimensionnelle définie. En revanche, au cours d'un stress comme la déshydratation ("dehydration"), la structure de certaines LEA est enrichie en structures secondaires en particulier en hélices amphiphiles (figure ci-dessous).
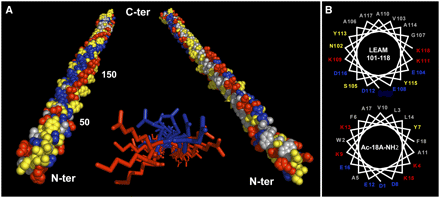
Source : Tolleter et al. (2007) Enrichissement en structures secondaires de certaines protéines LEA lors de la dessication Figure ci-contre, représentations schématiques des structures de la protéine LEA (AaVLEA1) d'Aphelenchus avenae (nématode), obtenues par dynamique moléculaire (champs de force Gromos 96) , pour différents pourcentages en eau (points rouges).
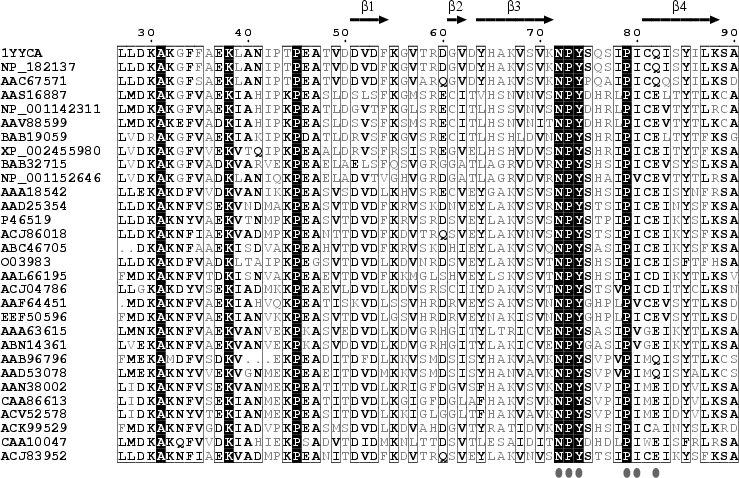
Source : Li & He (2009) Puisqu'aucune structure 3D de LEA n'a été obtenue à ce jour, la structure hypothètique de la protéine AaVLEA1 a d'abord été déterminée par modélisation par homologie ("protein threading - homology modeling") en utilisant comme modèle la structure 3D déterminée pour l'apolipoprotéine AI humaine (PDB : 1AV1). Voir un alignement des séquences de LEA de la classe 7 (E. Jaspard). Un motif conservé au sein de cette classe (comme pour les autres classes) en est la signature. |
| 8. Liens Internet et références bibliographiques |
|
|
|
Szabados & Savouré (2010) "Proline: a multifunctional amino acid" TIBS 15, 89 - 97 Timperio et al. (2008) "Proteomics applied on plant abiotic stresses: role of heat shock proteins (HSP)" J. Proteomics 71, 391 - 411 Vinocur & Altman (2005) "Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations" Curr. Opin. Biotechnol. 16, 123 - 132 Jullien & Berger (2008) "L'empreinte parentale, un phénomène épigénétique essentiel pour la reproduction des plantes" Med. Sci. (Paris) 24, 753 - 757 Hirayama & Shinozaki (2010) "Research on plant abiotic stress responses in the post-genome era: past, present and future" The Plant Journal 61, 1041 - 1052 |
|
|
Chaves et al. (2002) "How plants cope with water stress in the field? Photosynthesis and growth" Ann. Bot. Water Stress 89, 907 - 916 Chaves et al. (2009) "Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell" Ann. Bot. 103 : 551 - 560 Kosovà et al. (2011) "Plant proteome changes under abiotic stress - contribution of proteomics studies to understanding plant stress response" J. Proteomics 74, 1301 - 1322 |
|
|
Goyer A (2010) "Thiamine in plants: Aspects of its metabolism and functions" Phytochemistry 71, 1615 - 1624 Arrigo A.-P. (2005) "Repliement des protéines" M/S : médecine sciences 21, 619 - 625 Cornette et al. (2010) "Identification of Anhydrobiosis-related Genes from an Expressed Sequence Tag Database in the Cryptobiotic Midge Polypedilum vanderplanki (Diptera; Chironomidae)" J. Biol. Chem. 285, 35889 - 35899 Elmayan & Simon-Plas (2007) "Regulation of plant NADPH oxidase" Plant Signal Behav. 2, 505 - 507 Denison et al. (2011) "14-3-3 proteins in plant physiology" Semin. Cell. Dev. Biol. 22, 720 727 Khraiwesh et al. (2012) "Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants" Biochim. Biophys. Acta 1819, 137 - 148 |
|
|
Ritossa F.M. (1962) "A new puffing pattern induced by temperature shock and DNP in drosophila" Cell Mol. Life Sci. 18, 571 - 573 Ritossa F.M. (1964) "Experimental activation of specific loci in polytene chromosomes of Drosophila" Exp. Cell Res. 35, 601 - 607 Koskull-Döring et al. (2007) "The diversity of plant heat stress transcription factors" Trends Plant Sci. 12, 452 - 457 Rushton et al. (2010) "WRKY transcription factors" Trends Plant Sci. 15, 247 - 258 Raghavendra et al. (2010) "ABA perception and signalling" TIBS 15, 395 - 401 Reddy et al. (2011) "Coping with Stresses: Roles of Calcium- and Calcium/Calmodulin-Regulated Gene Expression" The Plant Cell 23, 2010 - 2032 GENEVESTIGATOR : Arabidopsis Microarray Database and Analysis Toolbox MapManWeb : Outil global de visualisation WEB |
|
|
Protéines LEA Dure LIII, Greenway SC, Galau GA (1981) "Developmental biochemistry of cottonseed embryogenesis and germination: changing messenger ribonucleic acid populations as shown by in vitro and in vivo protein synthesis" Biochemistry 20: 4162-4168 Galau GA, Dure LIII (1981) "Developmental biochemistry of cottonseed embryogenesis and germination: changing messenger ribonucleic acid populations as shown by reciprocal heterologous complementary deoxyribonucleic acid-messenger ribonucleic acid hybridization" Biochemistry 20: 4169-4178 Galau GA, Hugues DW, Dure LIII (1986) "Abscisic acid induction of cloned cotton late embryogenesis-abundant (Lea) mRNAs" Plant Mol. Biol. 7: 155-170 Dure LIII, Crouch M, Harada J, Ho T-HD, Mundy J, Quatrano R, Thomas T, Sung ZR (1989) "Common amino acid sequence domains among the LEAP of higher plant" Plant Mol. Biol. 12: 475-486 Galau GA, Wang HY-C, Hugues DW (1993) "Cotton Lea5 and LEA4 encode atypical late embryogenesis-abundant proteins" Plant Physiol. 101: 695-696 Li D, He X (2009) "Desiccation induced structural alterations in a 66-amino acid fragment of an anhydrobiotic nematode late embryogenesis abundant (LEA) protein" Biomacromolecules 10: 1469-1477. Hunault & Jaspard (2010) "LEAPdb: a database for the late embryogenesis abundant proteins" BMC Genomics 11, 22 Jaspard, Macherel & Hunault (2012) "Computational and statistical analyses of amino acid usage and physico-chemical properties of the twelve late embryogenesis abundant protein classes" PLoS One 7, e36968 |
|
![]()

{kind=link}