| Puces à ADN : analyse de l'article de Seki et al. (2002) |
| Tweet |
|
|
1. Caractéristiques et apports de l'étude 2. Obtention d'ADNc pleine longeur ("full-length cDNA") 3. Caractéristiques de la puce |
4. Vérification des données issues de la puce à ADN par Northern blot 5. Résultats généraux 6. Liens Internet et références bibliographiques |
|
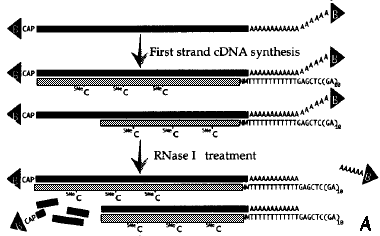
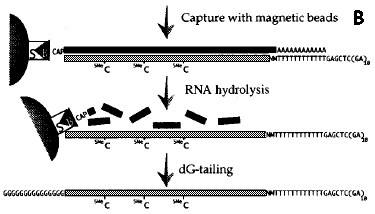
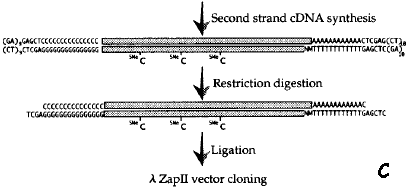
1. Caractéristiques et apports de l'étude a. Travail collossal en amont (voir Seki et al. (2001) : construction de plusieurs banques d'ADNc pleine longueur d'Arabidopsis thaliana => fabrication d'une puce contenant 7000 ADNc pleine longueur. Ces ADNc pleine longueur ont été isolés à partir d'une série de banques d'ADNc obtenues (voir article p284 - "Results and discussion") :
Remarques :
b. Dans cet article :
|
| Résumé des conditions d'études avec les puces à ADN | |
| Sondes fixées | Cibles hybridées (ARN messagers extraits) |
Plusieurs banques d'ADNc pleine longueur construites à partir d'Arabidopsis thaliana
=> obtention de 7000 ADNc pleine longueur pour la fabrication des puces à ADN. Issu d'un travail préliminaire => voir Seki et al. (2001). |
ARN messagers d'Arabidopsis thaliana (3 semaines de culture)
Voir article p 283 - "Materials and methods". |
c. Travail exigeant : seuls les gènes ayant un rapport d'expression supérieur à 5 [(log2(r)] sont analysés. d. Travail trés précieux pour la communauté scientifique :
|
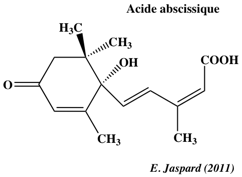
Principaux résultats de cette étude 1. Plusieurs mécanismes de la régulation de la transcription sont impliqués dans la voie de transduction du signal de l'ABA. 2. Il y a davantage de points communs entre les gènes impliqués dans la voie de signalisation de l'ABA et ceux liées à la sécheresse et à une forte salinité qu'entre la voie de signalisation de l'ABA et celle liée au froid. L'acide abcissique ("abscisic acid" - ABA) a été identifié en 1963 par F. Addicott et ses collaborateur, à partir des feuilles de cotonnier. C'est une hormone végétale synthétisée par les racines ou les feuilles (à l'intérieur des plastes). Structure : sesquiterpène - terpène dérivé de l'isoprène, composé en C15.
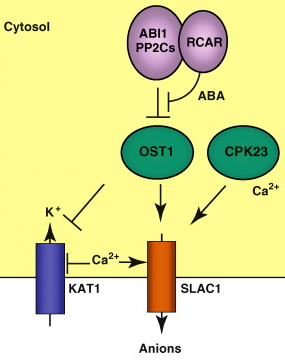
Exemples de rôles physiologiques de l'ABA Figure ci-dessous : Illustration du rôle de l'ABA dans l'ouverture de canaux ioniques.
Source : Raghavendra et al. (2010)
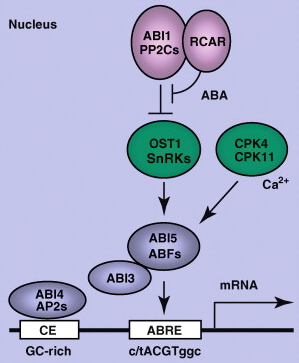
Autres rôles de l'ABA L'ABA est un messager capital de nombreuses voies de transduction du signal en réponse à des stress biotiques et abiotiques (exemple : limiter le stress hydrique en période de sécheresse). Figure ci-dessous : Rôle de l'ABA dans la régulation de la transcription de gènes (activation de facteurs de transcription de type ABI).
Source : Raghavendra et al. (2010) |
|
Intérets des ADNc pleine longueur La comparaison des séquences d'ADNc pleine longueur d'Arabidopsis thaliana (ou de n'importe quel organisme) avec les séquences génomiques (mais aussi les données d'EST) permet :
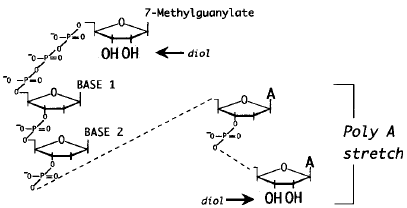
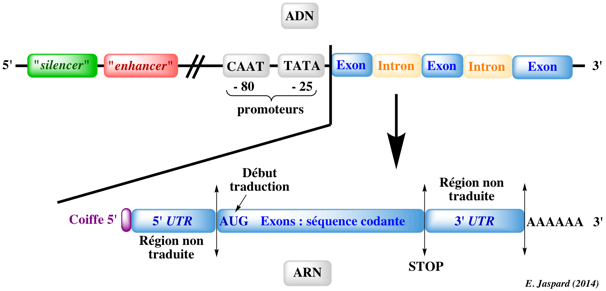
Autre intéret d'ADNc pleine longueur : il y a peu d'hybridation croisée ("cross-hybridization") avec des pseudogènes. Figure ci-dessous : schéma simplifié d'un gène Eucaryote et d'un transcrit issu de ce gène.
Les régions non traduites ("UnTranslated Regions") en 5' (5'-UTR) sont davantage conservées entre différentes espèces et ne changent pas beaucoup au sein d'une famille de gènes. A l'inverse, les régions 3'-UTR sont caractérisées par une plus faible conservation entre différentes espèces. |
| Eléments agissant en "cis" : séquences consensus de l'ADN située en amont des sites d'initiation de la transcription. Exemple : l'opéron lac. | ||
| procaryotes | région -10 ou boîte Pribnow (10 nucléotides avant le site d'initiation de la transcription) région -35 séquence "opérateur" de l'opéron lactose |
TATAAT TTGACA ------- |
| eucaryotes | région -10 ou boîte Goldberg-Hogness ou "TATA box" | TATAAT |
| Eléments agissant en "trans" (action d'une molécule sur une autre) : ce sont les facteurs de transcription (protéines) qui se fixent sur ces séquences d'ADN (les éléments agissant en "cis") . | ||
|
Applications des ADNc pleine longueur
|
|
3. Caractéristiques de la puce Les 7000 ADNc pleine longueur d'Arabidopsis thaliana déposés sur la puce ont été isolés à partir d'une série de banques d'ANc obtenues (voir article p284 - "Results and discussion") :
|
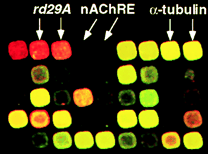
| contrôles positifs | ADNc issus des gènes rd et erd | gènes d'Arabidopsis thaliana inductibles par la dessication |
| contrôle interne | séquence d'ADN amplifiée par PCR | fragment d'ADN du phage lambda |
| contrôles négatifs |
fragment d'ADN du gène
d'un récepteur nicotinique de la souris (nAChRE) fragment d'ADN du gène d'un homologue du recepteur de glucocorticoïde de souris |
ADN sans aucune homologie avec une quelconque séquence d'Arabidopsis thaliana qui permet d'évaluer toute hybridation non spécifique. |
L'intensité de chaque spot reflète le niveau d'expression de chaque gène.
Source : Seki et al. (2001)
Ces témoins permettent le calcul des rapports d'intensité de fluorescence (IF) puis la valeur [(log2(r)] entre la condition normale (non stressée) et la condition de stress. Par exemple, le rapport dans le cas de la dessication est calculé de la manière suivante : [IF de chaque spot d'ADNc en condition de dessication] / [IF de chaque spot d'ADNc en condition normale] --------------------------------------------------------------------------------------------------------------------- IF du spot d'ADN du phage lambda en condition de dessication] / [IF du spot d'ADN du phage lambda en condition normale] |
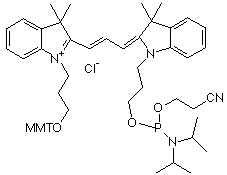
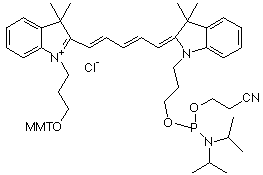
Le marquage des cibles consiste en l'incorporation de nucléotides portant :
Source : Amersham Biosciences Limited Ces 2 molécules sont les plus classiquement utilisées. MMT : groupe 4-monomethoxytrityle. |
| cyanine | nom | longueur d'onde émission fluorescence | couleur |
| Cy3 | indodicarbocyanine 3-1-O-(2-cyanoethyl)- (N,N-diisopropyl)-phosphoramidite | 563 - 570 nm | vert |
| Cy5 | indodicarbocyanine 5-1-O-(2-cyanoethyl)- (N,N-diisopropyl)-phosphoramidite | 662 - 670 nm | rouge |
|
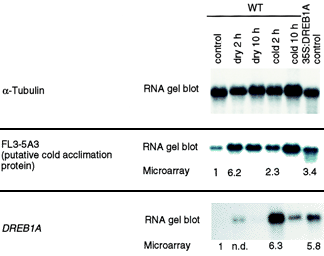
4. Vérification des données issues de la puce à ADN par Northern blot Les gènes dont le rapport d'augmentation d'expression (par exemple : dessication / non stressé) est supérieur à 2 fois celui du contrôle positif sont réputés inductibles par le stress considéré. La seule manière de confirmer ce genre de résultat est de faire un Northern blot à partir des ARNm correspondants aux ADNc candidats. Parfois, les résultats ne concordent pas et les "faux-positifs" issus de la puce à ADN peuvent être dûs aux biais suivants :
Figure ci-dessous, comparaison des données issues de la puce à ADN et des résultats du Northern blot.
Source : Seki et al. (2001) Conditions testées :
Les rapports d'augmentation d'expression sont indiqués en dessous de chaque autoradiographie. Voir N° accession GenBank : AB044404 - cold acclimation protein WCOR413 |
|
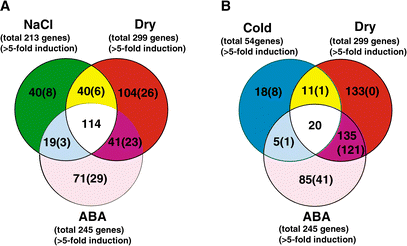
Diagramme de John Venn (1834-1923) : schémas géométriques utilisés pour représenter des relations logico-mathématiques.
Exemple : 245 gènes induits par l'ABA = 41 gènes + 71 gènes + 19 gènes + 114 gènes |
| stress | nombre de gènes dont l'expression augmente | nombre de gènes dont l'expression diminue |
| Nomenclature des ADNc de RIKEN (RAFL : "Riken Arabidopsis Full-Length"): RAFL XX - YY - LettreZZ | ||
| ABA |
245 gènes 22 codent pour des facteurs de transcription : 1 de la famille DREB / 2 - famille ERF / 5 - famille doigt de zinc / 1 - famille WRKY / 3 - famille MYC/MYB / 1 - famille bHLH / 4 - famille NAC / 2 - famille homéodomaine / 2 - famille bZIP / 1 - autre famille |
34 |
| dessication | 54 | 77 |
| froid | 299 | 70 (ou 79 ? - article p. 290) |
| forte salinité | 213 | 79 (ou 70 ? - article p. 290) |
Les facteurs de transcription impliqués dans la réponse aux stress Chez Arabidopsis, on en dénombre plus de 2000 (> 7% du protéome). Ils font partie des 58 familles suivantes : AP2/EREBP, ABI3/VP1, ARF, bHLH, bZIP, HB, HSF, MYB, NAC, WRKY
Source : Stress responsive TranscrIption Factor Database Exemples de bases de données de facteurs de transcription spécifiques des plantes : |
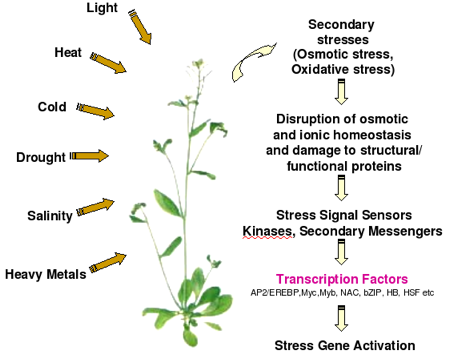
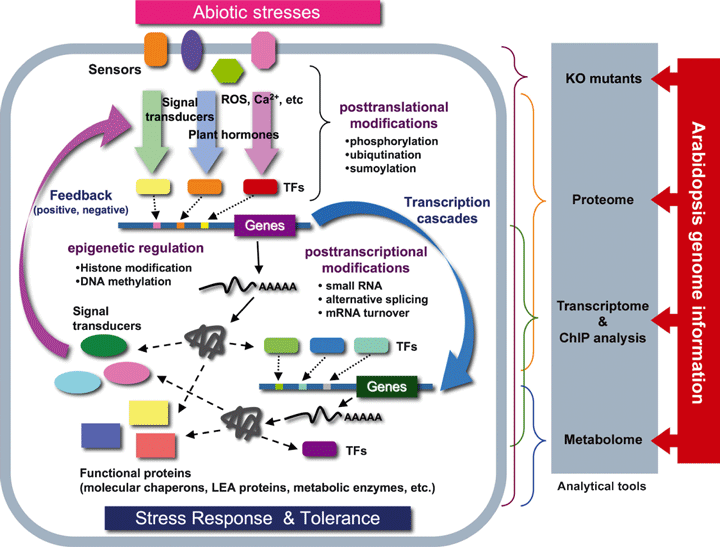
Modèle général de la réponse aux stress abiotiques chez les plantes Les cellules des plantes reçoivent des signaux via maints "senseurs" (peu ou pas encore connus) et ces signaux sont transduits via de nombreuses voies de signalisation dans lesquelles interviennent des hormones, des transducteurs du signal, des régulateurs de la transcription, ... Les gènes inductibles par les stress sont directement régulés par de nombreux signaux de stress.
Source : Hirayama & Shinozaki (2010) Certains gènes inductibles par les stress sont régulés par des facteurs de transcription ("TFs" dans la figure ci-dessus) dont l'expression est elle-même induite par les stimuli liés aux stress (cascade de transcription). Certains gènes inductibles par les stress codent des protéines dont la fonction est directement impliquée dans la tolérance aux stress. Certains gènes inductibles par les stress codent des protéines régulatrices (comme les transducteurs du signal) qui, trés probablement, sont impliquées dans des boucles de contrôle en amont ou des boucles de rétro-contrôle ("feedback positive / negative") de la réponse aux stress. L’épigénétique (figure ci-dessus) est un système de régulation fondamental, au-delà de l’information contenue dans la séquence d'ADN, puisqu'elle étudie les modifications, transmisssibles entre générations et réversibles, de l'expression des gènes sans qu'il y ait altération des séquences (modification et maintien de l’expression d’un gène sans modification de l’ADN). |
Quelques exemples de familles de facteurs de transcription Famille AP2 (APETALA2) / EREBP ("ethylene-responsive element binding proteins") : ils sont spécifiques des plantes. Ils contiennent un domaine de fixation à l'ADN appelé AP2. La sous-famille de facteurs de transcription DREB ("dehydration responsive element binding proteins") fait partie de cette famille. Famille bZIP ("basic leucine zipper") : leur nom vient du fait qu'ils contiennent un domaine de fixation à l'ADN mixte constitué d' une séquence en acides aminés basiques et d'un motif riche en leucine à intervalles réguliers ("leucine zipper"). Les facteurs de transcription GBF1, GBF2 et GBF3 ("Arabidopsis bZlP family of G-box binding factors") intéragissent avec le motif palindrome "G-box" (CCACGTGG) trouvés dans de nombreux promoteurs de plantes. Famille HSF : ("heat shock transcription factor") : ils sont trimériques avec un domaine de fixation à l'ADN qui reconnaît la séquence répétée (nGAAn) et un domaine impliqué dans l'oligomérisation. Ils sont impliqués dans la réponse dite "de choc thermique" et la synthèse d'HSP ("heat shock proteins"). Il existe 3 classes (A, B et C). Ces facteurs de transcription sont organisés en modules fonctionnels comme le facteur de transcription AtHsfA2 de Arabidopsis. Famille WRKY : ils sont ainsi nommés parce qu'ils possèdent un domaine de fixation à l'ADN (du côté N-terminal) qui contient, une ou deux fois, la séquence en acides aminés (quasi invariante) WRKY. Ils sont classés en fonction du nombre de motif WRKY et de leur motif "zinc-finger-like" : Cx[4,5]Cx[22,23]HxH ou Cx7Cx23HxC. |
| 6. Liens Internet et références bibliographiques |
|
Plant Promoter Database Database of Orthologous Promoters The Database of Arabidopsis Transcription Factors AthaMap : genome-wide map of potential transcription factor and small RNA binding sites in Arabidopsis thaliana Arabidopsis Gene Regulatory Information Server Plant TFDB : Plant Transcription Factor Database DATF: Database of Arabidopsis Transcription Factors STIFDB : Stress responsive TranscrIption Factor Database |
|
|
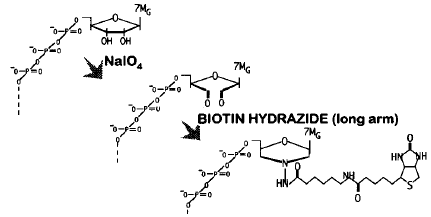
Seki et al. (2002) "Monitoring the expression pattern of around 7,000 Arabidopsis genes under ABA treatments using a full-length cDNA microarray" Funct. Integr. Genomics 2, 282 - 291 Carninci et al. (1996) "High-efficiency full-length cDNA cloning by biotinylated CAP trapper" Genomics 37, 327 - 36 |
|
|
Seki et al. (2001) "Monitoring the Expression Pattern of 1300 Arabidopsis Genes under Drought and Cold Stresses by Using a Full-Length cDNA Microarray" Plant Cell 13, 61 - 72 Raghavendra et al. (2010) "ABA perception and signalling" TIBS 15, 395 401 |
|
![]()