| Les différents types de photosynthèse : plantes en C3, en C4 et CAM |
| Tweet |
|
|
1. Introduction 2. Le mécanisme des plantes en C3 3. Le mécanisme des plantes en C4 |
4. Le mécanisme des plantes CAM (Crassulacean Acid Metabolism) 5. La phosphoenolpyruvate carboxylase (PEPCase) des plantes en C4 et des plantes CAM 6. Liens Internet et références bibliographiques |
|
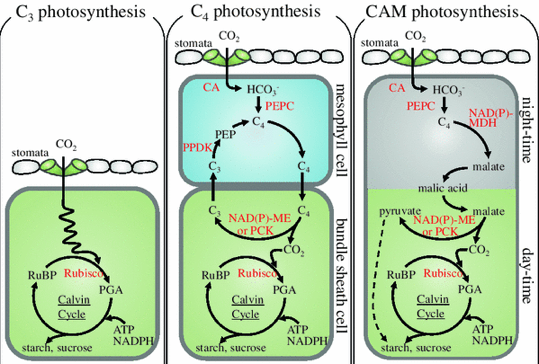
1. Introduction Il existe différents modes de fixation du CO2 chez les plantes, au cours de la photosynthèse. Ces mécanismes diffèrent par l'efficacité de cette étape de carboxylation. Le type de photosynthèse d'une plante est déterminé par le nombre d'atomes de carbone de la 1ère molécule organique formée lors de la fixation du CO2.
Source : Yamori et al. (2014) Les plantes en C3 convertissent le CO2 en un composé à 3 carbones (l'acide phosphoglycérique - PGA) avec la ribulose-1,5-bisphosphate carboxylase / oxygénase (RuBisCO). Les plantes en C4 et les plantes CAM convertissent le CO2 en un intermédiaire à 4 carbones (l'oxaloacetate - OAA) avec la phosphoenolpyruvate carboxylase (PEPC). Les plantes CAM diffèrent des plantes en C4 car les plantes CAM fixent le CO2 la nuit pour le stocker en tant qu'intermédiaire à 4 carbones (l'acide malique). Il existe 3 sous-types de plantes en C4, basés sur l'enzyme de décarboxylation de l'acide à 4 carbones : l'enzyme NADP-malique (NADP-ME), l'enzyme NAD-malique (NAD-ME) et la phosphoenolpyruvate carboxykinase (PCK). Il existe 2 sous-types de CAM, basés sur l'enzyme de décarboxylation de l'acide à 4 carbones : l'enzyme NAD(P) -ME et la PCK. CA : anhydrase carbonique / RuBP : ribulose-1,5-bisphosphate / PEP : phosphoénolpyruvate / PPDK : pyruvate phosphate dikinase / NAD(P)-MDH : NAD(P)-malate déshydrogénase
|
|
2. Le mécanisme des plantes en C3 La première étape du cycle de Calvin - Benson - Bassham (cycle RPP) est la carboxylation du ribulose 1,5 bisphosphate pour former un composé à 3 atomes de carbone, le 3-phosphoglycérate. Cette réaction est catalysée par la ribulose 1,5-bisphosphate carboxylase - oxygénase ou RuBisCO. La plupart des plantes fonctionnent selon ce mécanisme. Comme son nom l'indique, la RuBisCO catalyse en fait 2 réactions :
La photorespiration peut être préjudiciable à la plante du fait d'une diminution du taux net de photosynthèse. |
|
3. Le mécanisme des plantes en C4 Chez certaines plantes (maïs, canne à sucre, sorgho, ...) il existe une autre voie d'assimilation du carbone qui se superpose au cycle RPP. Dans cette voie, le CO2 forme un acide à 4 carbones (et non à 3 carbones) et 2 types de cellules sont impliqués. Les plantes qui possèdent cette voie n'ont pratiquement pas d'activité photorespiratoire. Cette voie consiste en :
Les plantes qui ont ce mécanisme possèdent une autre enzyme capable de fixer le CO2 : la phosphoénolpyruvate carboxylase ou PEPcase, uniquement présente dans les cellules du mésophylle (figure ci-dessous). |
|
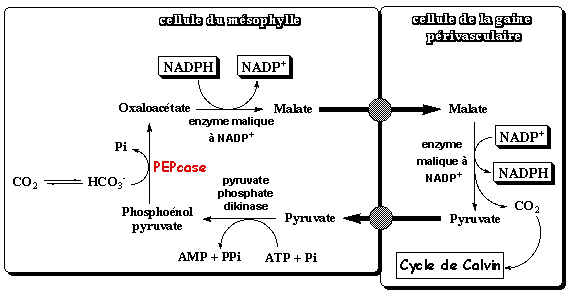
Le CO2 atmosphérique est hydraté HCO3-. Le phosphoénolpyruvate (PEP) et HCO3- se combinent par une réaction de carboxylation catalysée par la PEPcase (dépourvue d'activité oxygénase) et forment l'oxaloacétate (acide en C4).
Adapté de : "Photosynthèse et fixation du gaz carbonique" - Université Lausanne Selon l'espèce de plante en C4, l'oxaloacétate est réduit en malate ou transaminé en aspartate. Les acides en C4 passent dans les les cellules de la gaine périvasculaire voisines par des "tunnels" inter-cellulaires appelés plasmodesmes. La paroi des cellules de la gaine périvasculaire est imperméable aux gaz. En conséquence, la décarboxylation des acides en C4 accroit notablement la concentration en CO2. Cet enrichissement en CO2 rend la photorespiration nulle ou très faible chez les plantes en C4 et ce d'autant plus que les les cellules du mésophylle sont dépourvues en RuBisCO. Suivant les plantes il existe 3 types de métabolisme de type C4, qui font intervenir des intermédiaires et des compartiments cellulaires supplémentaires. Le nom de ces métabolisme est donné en fonction de l'enzyme de décarboxylation :
|
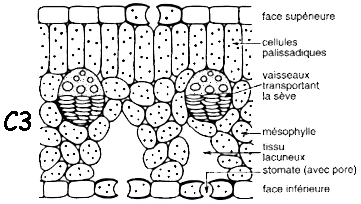
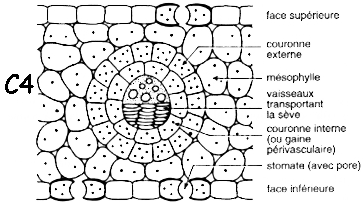
| Anatomie des plantes en C3 | Anatomie des plantes en C4 (dite en couronne - "Krantz anatomy") |
| mésophylle lacuneux avec des chloroplastes trés riches en grana | mésophylle avec peu ou pas de lacune qui entoure complètement la gaine périvasculaire |
| cellules de la gaine périvasculaire dépourvues de capacité photosynthétique | cellules de la gaine périvasculaire volumineuses contenant des chloroplastes trés pauvres en grana |
|
|
|
| Source figures : "Physiologie végétale" (1995) - Laval-Martin & Mazliak | |
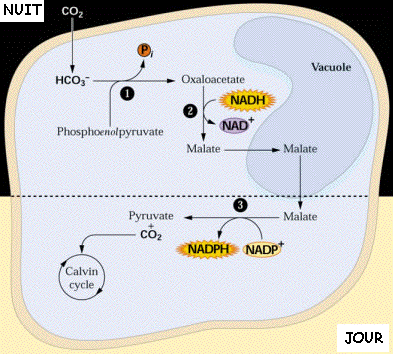
4. Le mécanisme des plantes CAM (Crassulacean Acid Metabolism) Ces plantes grasses (désert, milieux riches en sel) possèdent la RuBisCO et la PEPcase comme les plantes de type C4. Leur mode de fonctionnement leur permet de réduire les pertes d'eau. Cependant, la fixation du CO2 n'est pas séparée dans l'espace (mésophylle / gaine périvasculaire) mais dans le temps : nuit/jour. Durant la nuit :
Source : "Photosynthèse et fixation du gaz carbonique" - Université Lausanne Pendant la journée :
|
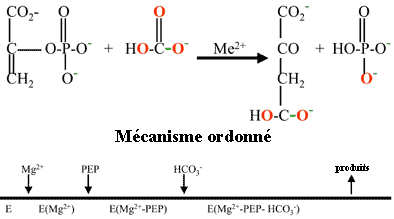
5. La phosphoenolpyruvate carboxylase (PEPCase) des plantes en C4 et des plantes CAM La réaction catalysée par la PEPcase (EC 4.1.1.31) suit un mécanisme ordonné : l'enzyme fixe d'abord Mg2+, puis le PEP et HCO3- en dernier.
|
|
Le trés faible KM de la PEPcase pour HCO3- (environ 2 µM) facilite à la fois :
Source : B. Buchanan - "Plant Physiology Online" |
|
|
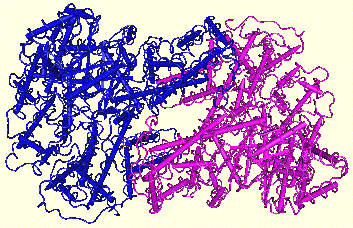
Ci-contre,
structure de la phosphoenolpyruvate
carboxylase de Zea mays (Matsumura & Kai - 2001).
Code accès : PDB 1JQO La PEPCase est un homotétramère (110 kDa pour chaque sous-unité). |
|
| 6. Liens Internet et références bibliographiques |
| "Physiologie végétale" (1995) volume I - "Nutrition et métabolisme", Danielle Laval-Martin et Paul Mazliak, Collection "Méthodes", Hermann / ISBN : 2 7056 6253 7 | |
|
B. Buchanan - "Modulation of Phosphoenolpyruvate Carboxylase in C4 and CAM Plants" - "Plant Physiology Online" Yamori et al. (2014) "Temperature response of photosynthesis in C3, C4, and CAM plants: temperature acclimation and temperature adaptation" Photosynth Res. 119, 101 - 117 |
|
![]()