| Régulation de la néoglucogénèse |
| Tweet |
|
|
1. Introduction 2. Rappel sur la réaction catalysée par la pyruvate kinase 3. Réaction catalysée par la pyruvate carboxylase 4. Réaction catalysée par la PEP carboxykinase |
5. Réaction catalysée par la fructose-1,6-bisphosphatase 6. Réaction catalysée par la glucose-6-phosphatase 7. Régulation de la néoglucogénèse 8. Liens Internet et références bibliographiques |
|
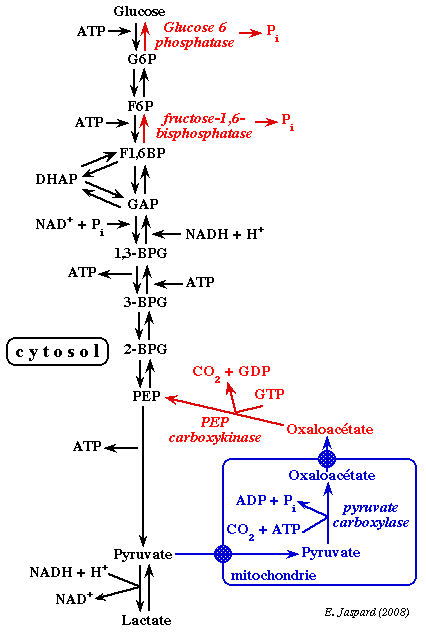
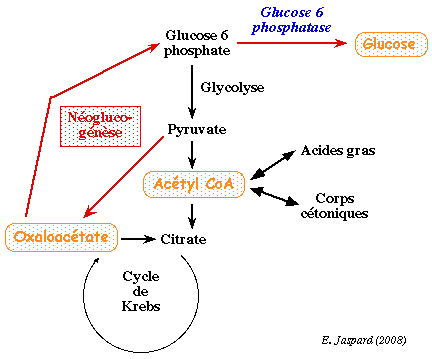
1. Introduction La réserve de glycogène du foie qui approvisionne les autres tissus en glucose s'épuise à la suite d'un jeûne par exemple. Le glucose indispensable est alors fourni par la néoglucogénèse (ou gluconéogénèse), figure ci-dessous.
La néoglucogénèse est la synthèse de novo du glucose à partir de substrats carbonés non-glucidiques (précurseurs de la voie) : les acides aminés dits néoglucogènes, le lactate, le glycérol et le pyruvate. Chez l'homme, elle a lieu essentiellement dans le foie et, dans une moindre mesure et dans certaines conditions, dans les reins et l'intestin. Les réactions de la néoglucogénèse sont les mêmes chez les animaux, les végétaux, les champignons et les microorganismes. La néoglucogénèse utilise une grande partie des enzymes de la glycolyse : celles qui catalysent des réactions au voisinage de l'équilibre. Ces enzymes sont cytosoliques. |
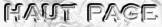
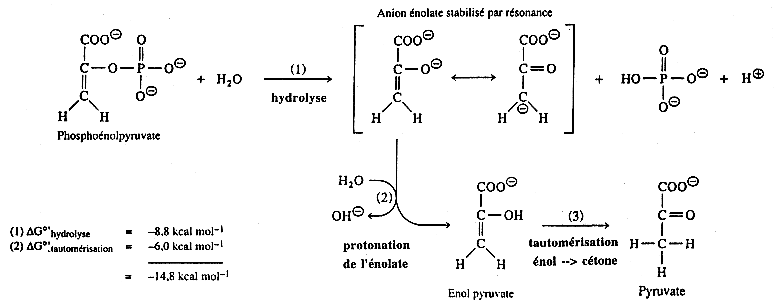
Cependant, trois réactions de la glycolyse sont caractérisées par des variations d'énergie libre de Gibbs si négatives qu'elles sont irréversibles dans les conditions physiologiques (ΔG'). Ces réactions sont celles catalysées par l'hexokinase (ou la glucokinase), par la phosphofructokinase et par la pyruvate kinase. |
| Enzyme | ΔG' (kcal.mol-1) |
| hexokinase ou glucokinase | - 8,0 |
| phosphofructokinase | - 5,3 |
| pyruvate kinase | - 4,0 |
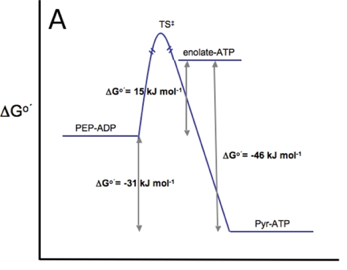
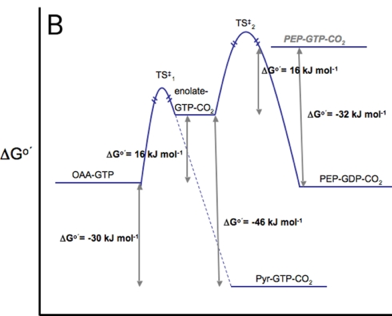
Ci-dessous : comparaison des barrières énergétiques d'activation franchies lors des réactions catalysées respectivement par la pyruvate kinase (figure A) et la PEP carboxykinase (figure B).
Source :Carlson & Holyoak (2009) En conséquence, ces réactions doivent être "contournées" lors de la néoglucogénèse. |
| glycolyse Voir la structure de ces enzymes |
néoglucogénèse | ||
hexokinase |
glucose + ATP ---> glucose-6-phosphate + ADP | glucose-6-phosphatase |
glucose-6-phosphate + H2O ---> glucose + Pi |
|
phosphofructokinase |
fructose-6-phosphate + ATP ---> fructose-1,6-bisphosphate + ADP | fructose-1,6-bisphosphatase |
fructose-1,6-bisphosphate + H2O ---> fructose-6-phosphate + Pi |
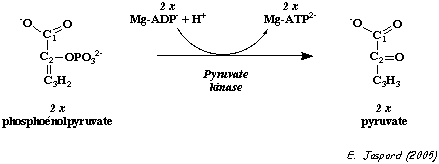
| pyruvate kinase E.C. 2.7.1.40 |
phosphoénolpyruvate + ADP ---> pyruvate + ATP |
pyruvate carboxylase PEP carboxykinase |
pyruvate + HCO3- + ATP ---> oxaloacétate + ADP + Pi oxaloacétate + GTP ---> phosphoénolpyruvate + GDP + CO2 |
| Les modalités de la néoglucogénèse dépendent de la localisation cellulaire des enzymes et de la nature des précurseurs de la voie. | |||
| glucose-6-phosphatase | fructose-1,6-bisphosphatase | pyruvate carboxylase | PEP carboxykinase |
| réticulum endoplasmique | cytosol | mitochondrie | cytosol ou mitochondrie |
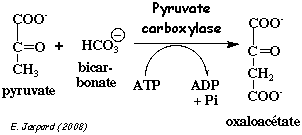
3. Réaction catalysée par la pyruvate carboxylase La pyruvate carboxylase est un homotétramère de 520.000 Da. La pyruvate carboxylase catalyse la formation de l'oxaloacétate à partir du pyruvate (figure ci-dessous).
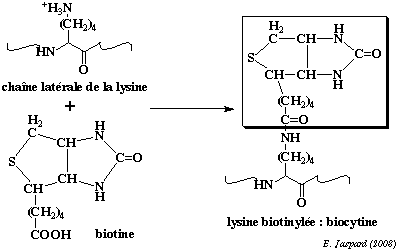
Cette réaction a lieu dans la mitochondrie. Le pyruvate du cytosol y est importé par un transporteur (symport avec un proton). Chaque protomère de pyruvate carboxylase contient une lysine à laquelle est liée une molécule de biotine. La biotine et la chaîne latérale de la lysine forment un bras flexible qui permet au groupe fonctionnel de la biotine d'aller et venir entre les deux sites actifs de la pyruvate carboxylase.
Sur l'un
des sites actifs de la pyruvate
carboxylase la biotine est carboxylée :
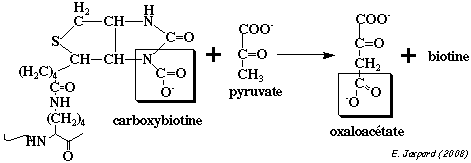
Au niveau de l'autre
site actif de la pyruvate
carboxylase, le CO2 activé est transferré de la biotine au pyruvate :
|
|
L'acétyl-CoA est un activateur allostérique de la pyruvate carboxylase :
|
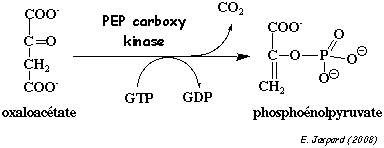
4. Réaction catalysée par la PEP carboxykinase (EC 4.1.1.32) La PEP carboxykinase (PEPCK) catalyse la transformation de l'oxaloacétate en PEP.
C'est une enzyme du foie et des reins. C'est un monomère d'environ 67.000 Da qui requiert un cation divalent (en particulier le manganèse) comme activateur de la réaction. Il existe deux types de PEPCK :
La PEPCK est inhibée par l'AMP. La PEPCK est localisée :
La réaction catalysée par la PEPCK est considérée comme l'un des points de contrôle de la néoglucogénèse. Non pas tant en ce qui concerne l'activité de la PEPCK elle-même, mais davantage en ce qui concerne la régulation de la biosynthèse de la PEPCK :
La synthèse de la PEPCK est induite par le cortisol. Le cortisol est synthétisé par les corticosurrénales à partir du cholestérol des LDL. C'est une hormone glucocorticoïde, qui active les facteurs de transcription des gènes codant les enzymes propres à la néoglucogénèse. La synthèse de la PEPCK est réprimée par l'insuline. |
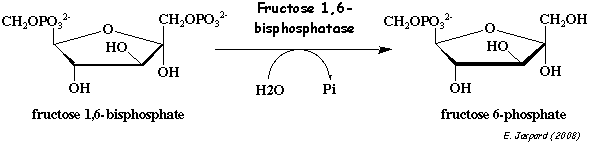
5. Réaction catalysée par la fructose-1,6-bisphosphatase (F-1,6BPase) La F-1,6BPase des vertébrés est un homotétramère d'une masse molaire de 148.000 Da. Chaque monomère est constitué de 2 domaines : le domaine fructose 1,6-bisphosphate qui contient le site actif et le domaine de fixation de l'AMP. Les sites de fixation des métaux se situent entre ces deux domaines.
La F-1,6BPase de foie est l'enzyme clé de la régulation de la néoglucogénèse. Elle catalyse l'hydrolyse du fructose 1,6-bisphosphate en fructose 6-phosphate et requiert des cations divalents (magnesium, manganese, zinc ou cobalt) comme co-facteur.
|
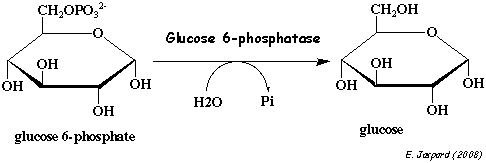
6. Réaction catalysée par la glucose-6-phosphatase (EC 3.1.3.9) La glucose-6-phosphatase (G6Pase) catalyse la dernière réaction de la néoglucogénèse : l'hydrolyse du glucose 6-phosphate en glucose.
N'étant pas phosphorylé, le glucose est plus libre de rejoindre le sang par des transporteurs. Ainsi la G6Pase joue un rôle important dans l'homéostasie de la concentration du glucose sanguin. La G6Pase est localisée dans la membrane du réticulum endoplasmique (RE) des cellules du foie :
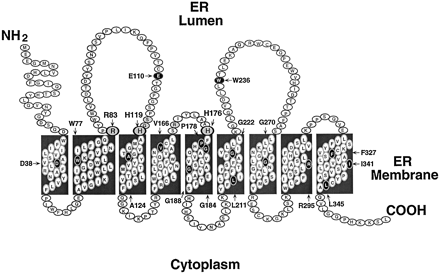
Du fait de sa forte association au réticulum endoplasmique, la structure exacte de la glucose-6-Pase n'est pas encore résolue. Ci-dessous, la structure supposée à 9 hélices trans-membranaires de la G6Pase humaine.
Source : Pan et al. (1998) Une sous-unité de cette enzyme semble jouer le rôle de translocase pour que le substrat atteigne ce site. Ci-dessous, un schéma du fonctionnement de la G6Pase selon l'hypothèse dite de l'unité catalytique - translocation du substrat ("substrate translocase-catalytic unit").
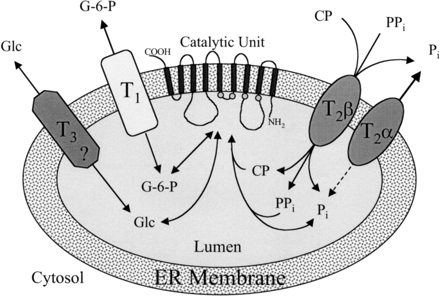
Source : Foster & Nordlie (2002) L'unité catalytique est la G6Pase localisée dans la membrane du réticulum endoplasmique. Elle est peu spécifique du substrat. Ce sont (au moins) 3 translocases additionnelles qui confèreraient au système sa spécificité en filtrant sélectivement l'accès des substrats et des produits. Ces transporteurs et/ou protéines auxiliaires avec les spécificités indiquées sont : T1 ("putative glucose-6-P transporter"), T2α, T2β et T3. Les cercles dans la boucle interne de l'unité catalytique indique les acides aminés qui englobent le motif phosphatase de la G6Pase. Un autre modèle a été proposé : le transport du substrat et une flexibilité conformationnelle combinés en une seule et même protéine (la G6Pase). |
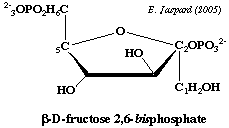
| 7. Régulation de la néoglucogénèse De manière générale, les effecteurs négatifs (inhibiteurs) de la glycolyse sont des effecteurs positifs (activateurs) de la néoglucogénèse. Le point de contrôle majeur de la dégradation ou au contraire de la synthèse du glucose est la régulation de l'activité de la phosphofructokinase-1 (PFK-1) et de la F-1,6BPase. Cette régulation est assurée essentiellement par le F2,6BP qui active la PFK-1 et est un inhibiteur allostérique de la F-1,6BPase.
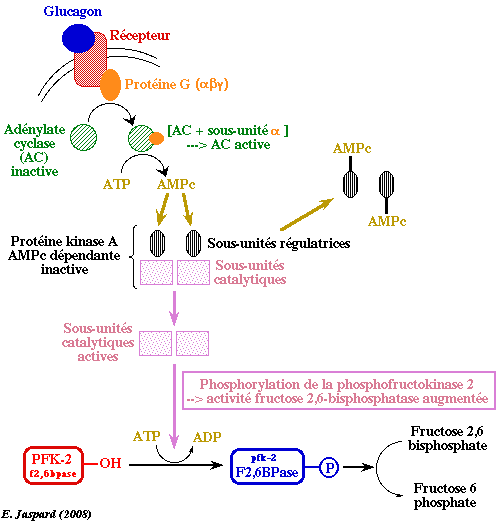
La concentration en F2,6BP est sous le contrôle du glucagon. Le glucagon est une hormone peptidique de 29 acides aminés sécrétée par le pancréas : elle stimule la lipolyse et la conversion des acides gras libres en cétones et inhibe la synthèse et favorise la dégradation des protéines. Les seules cellules qui possèdent de nombreux récepteurs du glucagon sont celles du foie. Le glucagon agit donc de façon extrêmement sélective.
Le glucagon se lie à un récepteur de glucagon qui active une protéine G appelée GS, constituée de 3 sous-unités (αβγ). La sous-unité α se fixe à l'adénylate cyclase qui est activée et qui convertit l'ATP en AMP cyclique (AMPc). Celui-ci active à son tour la protéine kinase A phosphoryle la [PFK2/F26BPase]. Cette phosphorylation :
Il en résulte une baisse de la concentration du F2,6BP.
Or le F2,6BP est :
La diminution d'insuline augmente les effets du glucagon :
Les changements du métabolisme induits par le glucagon favorisent la néoglucogénèse en défaveur de la glycolyse et entraînent une hyperglycémie. |
| 8. Liens Internet et références bibliographiques |
|
Un site avec une multitude de cours trés bien faits et de nombreuses illustrations : "Biochemistry of Metabolism" Pan et al. (1998) "Transmembrane topology of glucose-6-phosphatase" J. Biol. Chem. 273, 6144 - 6148 Foster & Nordlie (2002) "The biochemistry and molecular biology of the glucose-6-phosphatase system" Exp. Biol. Med. 227, 601 - 608 Dzugaj A. (2006) "Localization and regulation of muscle fructose-1,6-bisphosphatase, the key enzyme of glyconeogenesis" Adv. Enzyme Regul. 46, 51 - 71 Carlson & Holyoak (2009) "Structural Insights into the Mechanism of Phosphoenolpyruvate Carboxykinase Catalysis" J. Biol. Chem. 284, 27037 - 27041 |
![]()