| L'interférence ARN - ("RNA interference" - RNAi) - siRNA & miRNA |
| Tweet |
|
|
1. Introduction 2. Bref historique de la découverte de l'interférence ARN 3. Les siRNA ("small interfering RNA") 4. Les miRNA ("micro RNA") 5. Le complexe Drosha - Pasha (DGCR8) 6. La RNAse DICER 7. Le complexe RLC ("RISC-loading complex") et le complexe RISC ("RNA-Induced Silencing Complex") |
8. Rôles supposés des siRNA - miRNA dans l'inhibition de l'initiation de la traduction a. Cas général 9. Structure des protéines Argonaute 10. Le domaine PAZ (Piwi/Argonaute/Zwille) 11. Applications thérapeutiques de l'interférence ARN 12. Liens Internet et références bibliographiques |
| 1. Introduction
L'interférence ARN ("RNA interference") ou RNAi est une voie de régulation du taux d'ARN messagers traduits ou plus précisément de l'expression post-transcriptionnelle des gènes. Appellations historiques :
Andrew Fire et Craig Mello ont obtenu le Prix Nobel en 2006 pour l'explication des bases de ce mécanisme ("RNA interference - gene silencing by double-stranded RNA") chez le nématode Caenorhabditis elegans (travaux publiés en 1998). Le mécanisme de l'interférence ARN :
Deux types de petits ARN jouent un rôle central dans le mécanisme de l'interférence ARN: a. Les siRNA ("small interfering RNA") ou petits ARN interférents :
b. Les miRNA ("micro RNA") ou micro ARN :
D'autres types de petits ARN interférant jouent des rôles importants dans la régulation de l'expression des gènes. On peut citer :
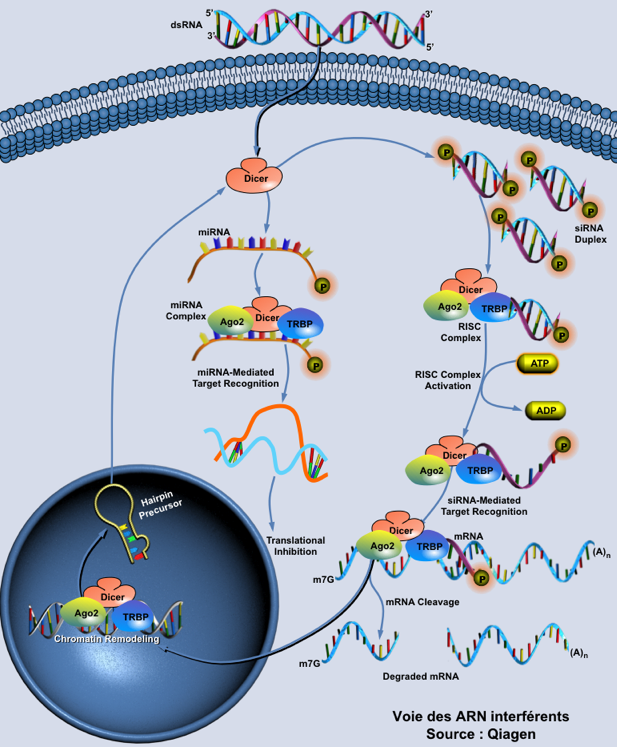
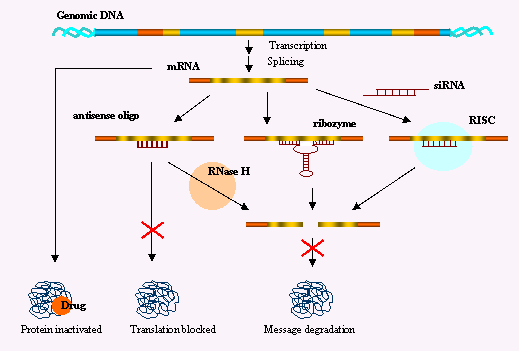
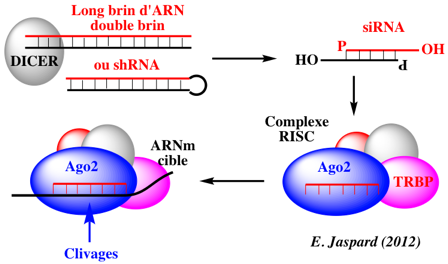
Voir l'algorithme pour la synthèse de shRNA ("short hairpin RNA") ou de a-miRNA ("artificial miRNA"). Enfin on peut mesurer l'importance de l'interférence ARN et de ses conséquences dans divers domaines (thérapie, amélioration des plantes, ...) en considérant le nombre de bases de données qui ont trait aux siRNA, miRNA, ... Schéma ci-dessous : ensemble des mécanismes qui aboutissent à l'interférence entre un ARN simple brin dit interférant (ARNi) avec un ARN messager (ARNm) spécifique. On aboutit à 2 cas de figure : soit la traduction de l'ARNm cible est inhibée, soit l'ARNm est dégradé. Dans les 2 cas, le taux de protéine synthétisée est diminué.
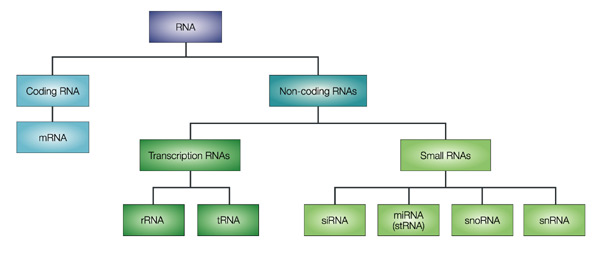
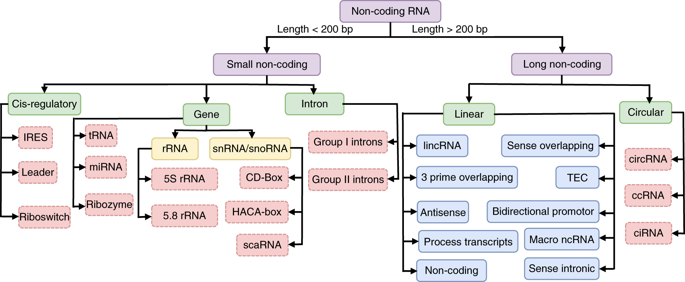
Source figure : Qiagen Les ARN et les petits ARN ("small RNAs" : snRNA, snoRNA, siRNA, miRNA, piRNA, ...) de 20 à 30 nucléotides. Ils participent à divers mécanismes génétiques, physiologiques et métaboliques.
Source : Buckingham S. (2003)
Source : Amin et al. (2019) |
|
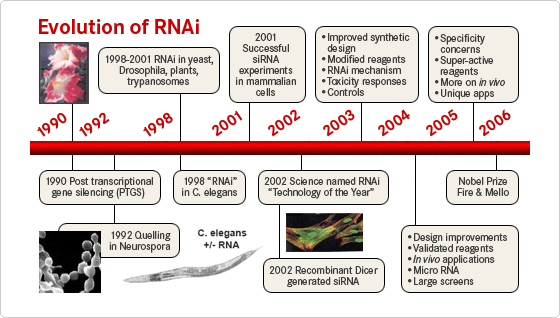
2. Bref historique de la découverte de l'interférence ARN En 1990, Napoli, Lemieux & Jorgensen ont été les premiers à décrire un phénomène d'interférence ARN, sans savoir qu'il s'agissait de ce mécanisme. L'objectif de leur étude était de déterminer si la chalcone synthase (CHS), une enzyme clé de la biosynthèse des flavonoïdes, est l'enzyme limitante dans la voie de biosynthèse de l'anthocyanine à l'origine de la coloration violette profonde du pétunia.
Ils ont donc surexprimé la CHS en introduisant dans des pétunias un transgène codant une CHS exogène :
Ces observations les ont menés à émettre l'hypothèse que le transgène était un co-suppresseur ("cosuppressing") du gène CHS endogène. Voir un développement et des éléments pour la conception bioinformatique de siRNA. Le terme "quelling" (étouffement) vient de Romano et Macino (1992) quand ils ont décrit un phénomène identique chez Neurospora crassa. Les techniques utilisant les ARN antisens pour inhiber la transcription d'un gène firent leur apparition. Un ARN antisens est un ARN complémentaire d'un ARN messager : l'association ARN antisens - ARN messager bloque donc la traduction de l'ARN messager. En 1993, Lee et al. ont montré qu'un petit ARN de Caenorhabditis elegans, LIN-4, régule le déroulement du développement en se fixant à une cible ARN messager, empêchant ainsi sa traduction. En 1994, Wassenegger et coll. montrèrent que l'introduction d'ARN double brin chez Arabidopsis thaliana méthyle l'ADN correspondant. En 1998, Fire, Mello et al. ont pensé à la présence de contaminants ARN double brin dans les expériences précédemment décrites. Ils ont donc fait l'hypothèse que l'élément clé n'est pas l'ARN simple-brin ("single-strand RNA" - ssRNA), mais l'ARN double-brin ("double-strand RNA" - dsRNA).
Source : Montgomery, Xu & Fire (1998)
Les articles de Fire, Mello et al. publiés en 1998 ont été les premiers à expliquer l'atténuation de l'expression de gènes endogènes. |
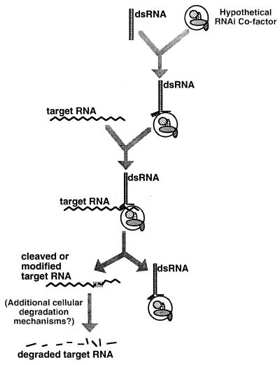
L'existence d'intermédiaires dans le mécanisme d'interférence ARN a été suggérée par Hamilton et Baulcombe en 1999. Bien que l'on pensait que le dsRNA devait se dérouler pour que le brin antisens puisse se fixer à l'ARNm, ce brin antisens pleine longueur n'a jamais été découvert. Hamilton et Baulcombe ont donc cherché des formes plus courtes du brin antisens issues du dsRNA. Ils ont émis l'hypothèse que l'ARN antisens sert de guide en se fixant à l'ARN messager cible, induisant ainsi sa dégradation. Quand Hamilton et Baulcombe ont détecté des brins antisens d'une longueur d'environ 25 nucléotides, ils ont suggéré que cette longueur était nécessaire pour la spécificité du mécanisme RNAi. L'année suivante, deux équipes ont constaté que des ARN de 21 à 23 nucléotides co-purifaient systématiquement par fractionnement, indiquant que le dsRNA est clivé en intermédiaires plus courts : les petits ARN interférant ("small interfering RNAs") ou siRNAs capables de se fixer à leur cible ARNm homologue, conduisant à la dégradation du transcrit. En 2000, on a montré que siRNA et miRNA ne sont pas une particularité du ver nématode (ARN lin-4 - nécessaire au passage du dernier stade larvaire au stade adulte, en identifiant un second petit ARN, let-7, hautement conservé (champignons, plantes, mouche, homme, ...), de même que sa longueur - environ 21 nucléotides (Pasquinelli et al., 2000). Il semblait improbable que le mécanisme RNAi existe chez les vertébrés puisque l'introduction d'ARN dans les cellules de mammifères déclenche une forte réaction antivirale non spécifique, la réponse interféron. En 2001, Elbashir et al. publient un article qui montre que le phénomène existe chez les mammifères. La mise en évidence de l'interférence ARN chez la drosophile a démontré l'existence d'un complexe protéique qui catalyse la séparation des deux brins de l'ARN interférant : le complexe appelé "RNA-Induced Silencing Complex" ou RISC (Hammond et al., 2000).
Source : Invitrogen
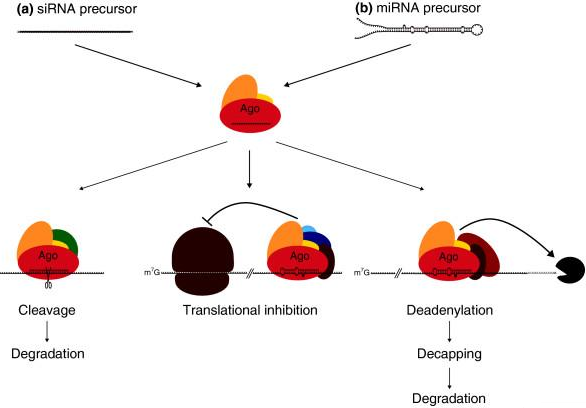
Figure ci-dessous : comparaison de divers de mécanisme d'extinction de gène ("gene silencing").
Source : NCBI |
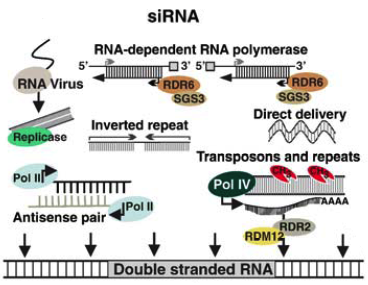
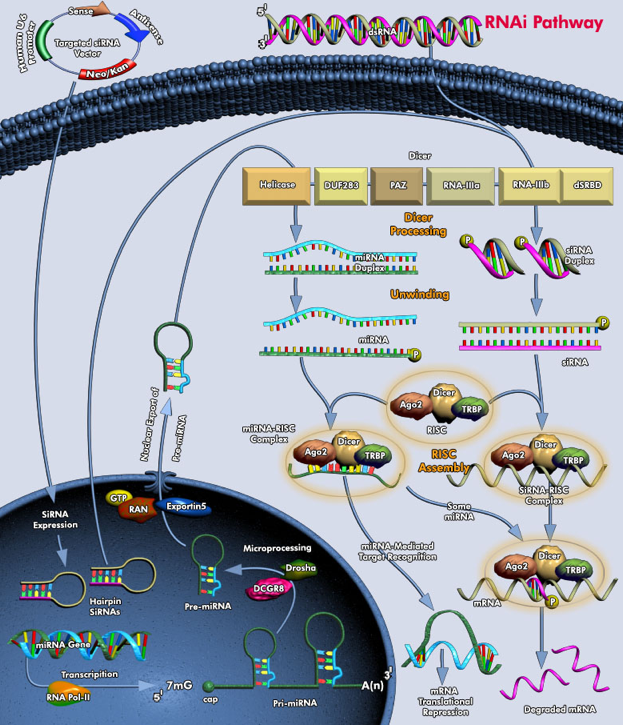
3. Les siRNA ("small interfering RNA") Précurseurs des siRNA a. Les long dsRNA ("long double-stranded RNA") : longs ARN double brin parfaitement complémentaires de plus de 200 nucléotides. Ils peuvent avoir pour origine des séquences répétées - inversées, être issus de la transcription de transgènes ou de transposons. A l'inverse, ils peuvent être synthétisés par des RNA polymérases RNA-dépendantes qui copient les ARN simple brin.
Source : Khraiwesh et al. (2012) b. Les shRNA ("short hairpin RNA") sont exprimés dans la cellule après transfection par des vecteurs viraux (plasmide, lentivirus, adénovirus, rétrovirus, ...).
Voir : Pan et al. (2012) "A dynamic perspective of RNAi library development" Trends Biotechnol. 30, 206 - 215 Ces précurseurs sont clivés tous les 21 à 25 nucléotides par une ribonucléase de type III appelée DICER ("éminceuse") : les courts fragments dsRNA obtenus sont appelés petits ARN interférents ("small interfering RNA" - siRNA). La ribonucléase DICER transfère les siRNA à un complexe multienzymatique : "RNA-Induced Silencing Complex" - RISC.
Ce mécanisme est donc lié au degré de la complémentarité des séquences [siRNA / cible ARNm] (voir la base de données "RNAi Atlas"). Par ailleurs, l'introduction de dsRNA de plus de 30 nucléotides dans des cellules mammifères active une réponse anti-virale de type interféron qui induit la dégradation non spécifique des ARNm et une réduction générale de la traduction. Rechercher de nouveaux siRNA ("design" / screening") nécessite donc que les ARN synthétisés contiennent moins de 30 nucléotides. Les travaux de Elbashir et al. ont montré que les siRNA avec un débordement en 3' constitué du dinucléotide UU sont les plus puissants. Il faut donc trouver une séquence de 21 nucléotides dans l'ARNm cible qui commence par un dinucléotide AA. On cherche le codon d'initiation de la transcription (AUG) : toutes séquences commençant par AA (et les 19 nucléotides suivants) constituent un site cible potentiel pour les siRNA. Modélisation - conception in silico ("design") de siRNA : voir "The RNAi web". |
|
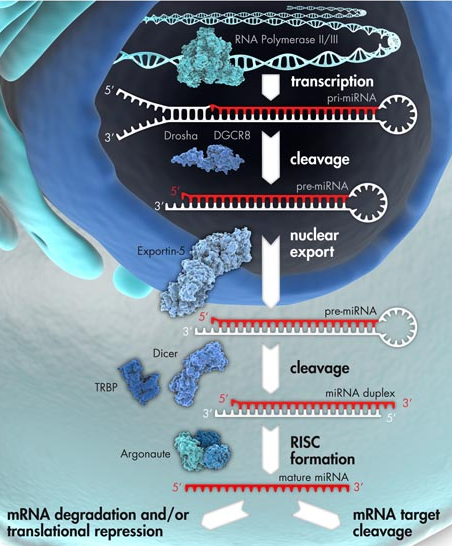
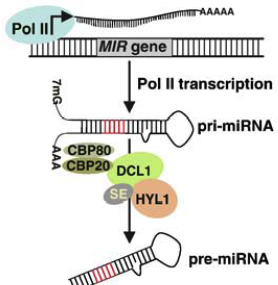
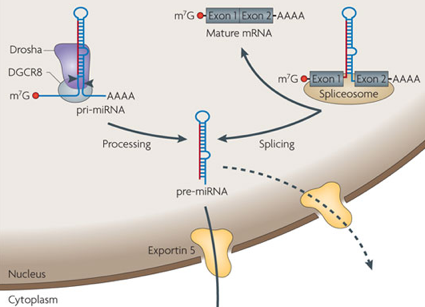
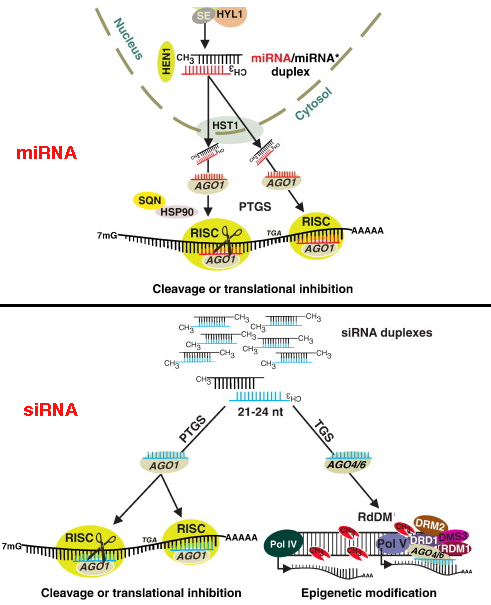
Les miRNA sont d'origine endogène : ils sont synthétisés dans le noyau sous forme de pri-miRNA ("primary-miRNA") à partir des gènes (poly-cistrons) miRNA transcrits par l'ARN polymérase II ou III.
Source : Qiagen Les pri-miRNA ont une structure en épingle à cheveux ("hairpin").
Figure ci-dessous : voies des ARN interférant : pri-miRNA, pre-miRNA, miRNA et siRNA.
Source : Qiagen Des protéines de type hélicase associées à DICER dissocient le dsRNA en deux ARN simple brin :
Les miRNA ont été identifiés dans tous les règnes (mammifères, drosophile, C. elegans, plantes …).
|
siRNA et miRNA chez les plantes Les siRNA et les miRNA des plantes interviennent dans de nombreux processus : transitions lors du développement, croissance des feuilles, polarité des organes, voie de signalisation par l'auxine, métabolisme des ARN, réponses des plantes aux stress biotiques et abiotiques, ...
La plupart des gènes miRNA :
Source : Khraiwesh et al. (2012) Les miRNA de plantes sont moins abondants que les siRNA. Dans la littérature , la notation est souvent : miRNA / miRNA* pour miRNA guide / miRNA passager.
Le décryptage du génome de Arabidopsis thaliana a montré qu'il contient :
Chez la tomate, on a dénombré (Bai et al, 2012) :
Chez les plantes, le clivage d'un ARNm ciblé par un miRNA peut entraîner la production de siRNA dits "secondaires" qui, à leur tour, peuvent induire une interférence ARN de gène en trans. Cette voie alternative dépend de la longueur du miRNA et de l'isoforme de la protéine AGO au sein du complexe RISC. Voir Manavella et al. (2012). |
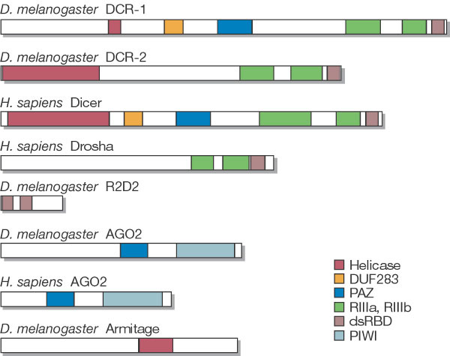
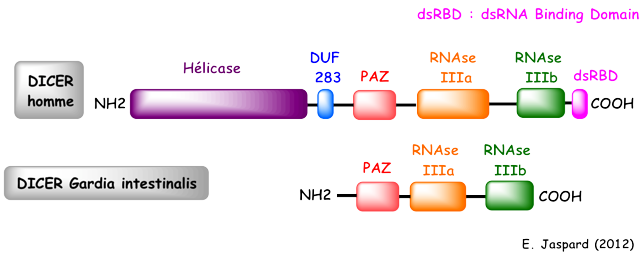
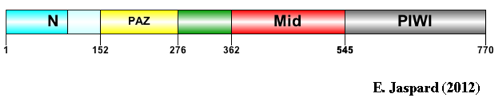
5. Le complexe Drosha - Pasha (DGCR8) Drosha (EC 3.1.26.3 / Uniprot : Q9NRR4 - homme, Q7KNF1 - Drosophile / PDB : 2KHX ) est une grosse ribonucléase de type III (1320 - 1380 acides aminés selon l'espèce) qui fixe les dsRNA. Les RNAses de type III ont un motif conservé de 9 acides aminés qui est une signature de leur site catalytique. Drosha en possède deux. Le domaine de fixation du dsRNA ("dsRNA-Binding Domain" - dsRBD) de Drosha est caractérisé par un repliement αβββα : 1 hélice α (Ser1263 à Thr1271), suivie de 3 feuillets β antiparallèles (Leu1283 à Gly1314), suivie de 1 hélice α (Ile1317 à Lys1331). Les différents domaines d'enzymes impliquées dans l'interférence ARN.
Source : Meister & Tuschl (2004) Drosha forme un complexe enzymatique (appelé "Microprocessor complex") avec DGCR8 ("DiGeorge syndrome critical region 8"). DGCR8 est une dsRBP ("dsRNA-Binding Protein") capable de fixer les fragments simple brin des pri-miRNA :
DGCR8 est connue sous le nom de Pasha chez la Drosophile et Caenorhabditis elegans. Drosha et Pasha sont localisées dans le noyau.
Source : Krol et al. (2010) |
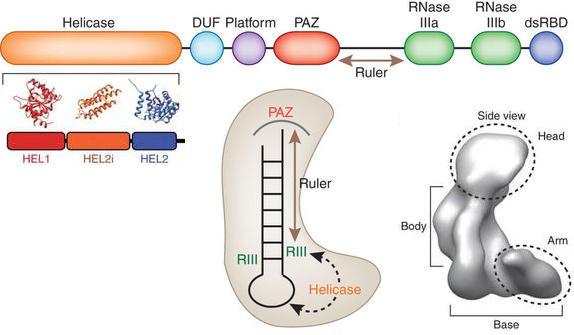
La protéine DICER (EC 3.1.26.3) est une endoribonucléase de la famille des RNAses de type III :
Le domaine "hélicase" à l'extrémité N-terminale a diverses propriétés potentielles :
Selon certains modèles, ce domaine se trouve dans l'espace à proximité de la boucle pre-miRNA et du coeur RNAse III.
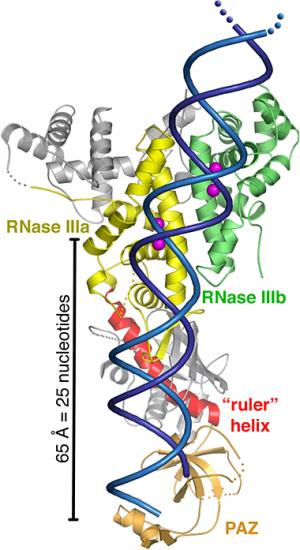
Sources : Sawh & Duchaine (2012) / Lau et al. (2012) Les domaines RNase IIIb et RNase IIIa clivent, respectivement, l'extrémité 5' et l'extrémité 3' des précurseurs RNA. L'une des principales difficultés pour déterminer la structure exacte de DICER est sa taille : plus de 200 kDa chez la plupart des espèces. Les modèles structuraux ne sont donc pas encore définitifs. La cryo-microscopie électronique devrait apporter de plus amples détails. La distance entre les domaines RNAses III et PAZ est déterminée par la longueur et l'angle de l'hélice appelée connecteur ("ruler helix") (figure ci-dessous).
Source : Macrae et al. (2006)
DICER2 possède une activité ATPase (domaine hélicase). L'hydrolyse de l'ATP est nécessaire pour que DICER2 clive les longs dsRNA.
|
7. Le complexe RLC ("RISC-loading complex") et le complexe RISC ("RNA-Induced Silencing Complex") Du fait de leur taille, l'essentiel des informations concernant ces complexes provient de leur reconstitution in vitro à partir des protéines individuelles purifiées. Les études structurales sont essentiellement effectuées par cryomicroscopie électronique ("negative-stain electron microscopy and single-particle analysis"). |
| Aperçu de la composition du complexe nucléasique RLC (appelé aussi "miRNA loading complex" ou miRLC) | |||
| Drosophile | Homme | ||
|
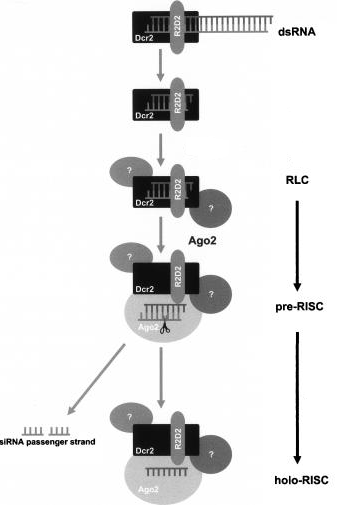
[DICER1 / R2D2] clivent les pré-miRNA en miRNA [DICER1 / R2D2] chargent les miRNA sur [EIF2C2 / Ago2] |
|
TRBP est très similaire de Loqs (Drosophile) et de PACT (mammifères).
|
|
Les protéines R2D2, R3D1, TRBP, Loqs, PACT sont des "dsRNA-binding proteins".
Elles servent d'intermédiaires entre le domaine PAZ de DICER et le domaine PAZ de la protéine Argonaute lors de la formation du complexe RISC. |
|||
Il est difficile de résumer tous les cas de figures (Drosophile, nématode, Homme, mammifères, plantes, ...) et ce d'autant que la composition des complexes varient en fonction de la nature des précurseurs et des différentes isoformes de certaines protéines. Par exemple, chez la Drosophile :
|
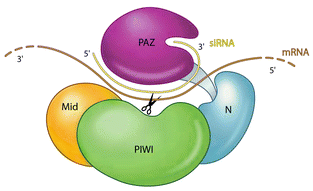
Formation du complexe RISC ("RNA-Induced Silencing Complex") Le complexe RISC est composé :
Le domaine PAZ de DICER du complexe RLC interagit avec le domaine PAZ d'une protéine Ago via R2D2 (ou TRBP) : le duplex siRNA ou miRNA est ainsi chargé sur Ago.
Source : Kim et al. (2007) Le brin passager est dégradé par Ago en (9 + 12) nucléotides.
Le complexe RISC est ensuite dirigé par le brin guide de l'ARN interférant vers l'ARNm cible dont il est complémentaire. Le devenir de l'ARNm cible n'est pas rigoureusement le même chez les plantes et les animaux. On peut schématiser ce devenir de la manière suivante : a. Si les siRNA ou miRNA ont une complémentarité parfaite avec la séquence de l'ARNm cible, ils induisent :
b. Si les miRNA ont une complémentarité imparfaite avec la séquence de l'ARNm cible, ils induisent une inhibition (répression) de la traduction de l'ARNm cible.
Source : Khraiwesh et al. (2012)
Source : Bratkovic et al. (2012) Chez la Drosophile, un nouvel activateur du complexe RISC a été identifié : C3PO ("Component 3 Promoter Of RISC").
|
8. Rôles supposés des siRNA et miRNA dans l'inhibition de l'initiation de la traduction Les membres de la superfamille de protéines Argonaute (Ago) sont impliqués dans l'interférence ARN tant au niveau transcriptionnel que post-transcriptionnel. Il existe différentes isoformes qui n'ont pas les mêmes spécificités : Ago2 (Drosophile) et AGO2 (Homme) vs. Ago1 (Drosophile) et AGO1, 3 et 4 (Homme). Les protéines Ago peuvent être divisées en 3 groupes :
Le complexe [Ago - siRNA] ou [Ago - miRNA] médient ensuite l'inhibition de l'initiation de la traduction de leurs ARNm cibles de plusieurs manières :
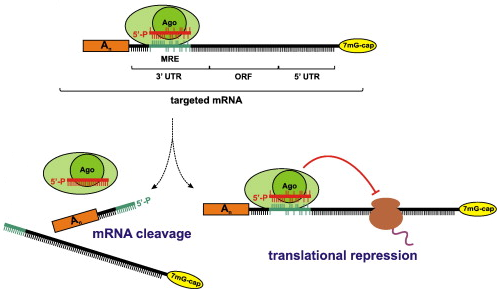
Source : Höck & Meister (2008) La complémentarité du siRNA ou du miRNA (porté par Ago) avec l'ARNm cible est parfaite, alors le complexe [Ago - siRNA] ou [Ago - miRNA] clive l'ARNm (figure de gauche ci-dessus). La complémentarité du miRNA avec l'extrémité 3' non traduite ("3' UTR") de l'ARNm cible est imparfaite, alors le complexe [Ago - miRNA] :
|
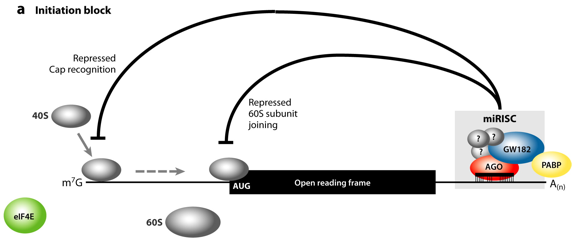
b. Mécanisme d'inhibition de l'initiation de la traduction par les miRNA eIF-4F est un complexe d'initiation de la traduction composé de 3 sous-unités (eIF-4E, eIF-4A et eIF-4G) et d'au moins 2 facteurs additionnels ("Poly(A)-Binding Protein" ou PABP, Mnk1 ou Mnk2). Parmi ses rôles, eIF-4F :
Le complexe [miRNA - RISC] inhibe l'initiation de la traduction de plusieurs manières :
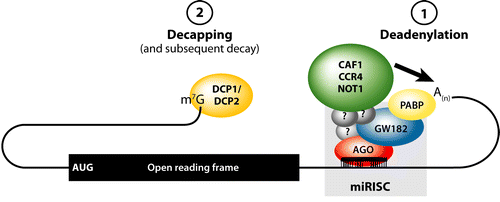
Source : Fabian et al. (2010) Le complexe [miRNA-RISC] interagit avec le complexe désadénylase CCR4-NOT1 pour la désadénylation de la queue poly(A) [notée A(n) dans la figure ci-dessus]. La désadénylation requière l'interaction de la protéine GW182 avec la protéine PABP qui fixe la queue poly(A). Après la désadénylation, la coiffe à l'extrémité 5' (m7G) est enlevé par le complexe DCP1-DCP2.
Source : Fabian et al. (2010) Légende de la figure ci-dessus : rectangle noir = phase de lecture ouverte ("open reading frame") ; CAF1 = "CCR4-Associated Factor 1" ; CCR4 = "Carbon Catabolite Repression 4 protein" ; NOT1 = "Negative On TATA-less". |
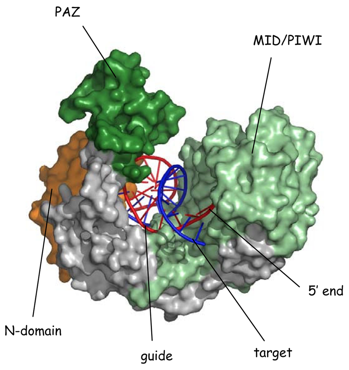
9. Structure des protéines Argonaute
Source : Parker J. (2010) Ago a la forme de 2 lobes qui contiennent chacun 2 domaines conservés au sein de la famille des protéines Argonaute :
L'extrémité C-terminale du domaine PIWI adopte un repliement de type RNAse H et possède le site catalytique (Asp-Asp-Asp/His - activité endonucléase). La catalyse est médiée par des ions Mg2+.
Source : Liu & Paroo (2010) |
|
Visualisation de Argonaute de Thermus thermophilus complexée à un fragment d'ADN à une résolution de 2,7 Å Code PDB : 3DLB Le chargement des structures peut prendre un peu de temps.
|
|
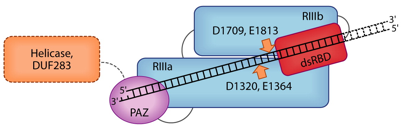
Ce domaine (PFAM : PF02170) est trouvé dans les 2 familles de protéines impliquées dans l'interférence post-transcriptionnelle : familles PIWI et DICER (incluant le facteur protéique Carpel). Le nom du domaine PAZ est ainsi appelé d'après les protéines "Piwi", "Argonaut" et "Zwille". Le domaine PAZ est composé de 2 sous-domaines :
Le domaine PAZ peut fixer les 2 nucléotides non appariés en 3' des siRNA et des miRNA : même si ce domaine n'est pas le site principal de fixation au sein de DICER ou du complexe RISC, il contribue à l'incorporation spécifique des siRNA et des miRNA dans la voie de l'interférence ARN. |
11. Applications thérapeutiques de l'interférence ARN Les pathologies sont très nombreuses, par exemple :
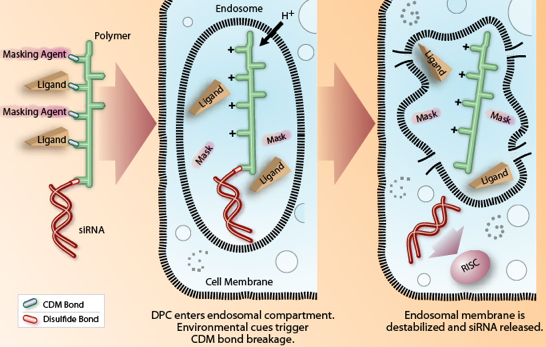
Cependant l'une des restrictions dans l'utilisation clinique des ARN interférents émane de leur temps de vie très court car ils sont rapidement dégradés par les nucléases du sérum ou des fluides extracellulaires. Les courts ARN double brins interférants sont donc instables et il génère des effets secondaires indésirables (stimulation du système immunitaire, sécrétion de cytokines inflammatoires, ...) s'ils sont administrés systématiquement, raison pour laquelle il n'y a pas ou peu de thérapie clinique à base d'interférence ARN. Il faut donc mettre au point des systèmes qui permettent :
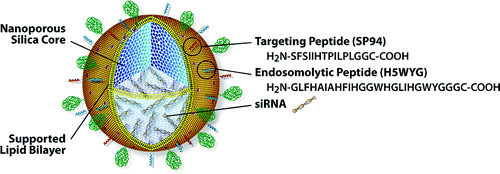
Source : Roche a. L'empaquetage Des "protocellules" - nanoparticules à base de silice mésoporeuse (exemples : "Mobil Crystalline Materials" - MCM-41 / "Santa Barbara Amorphous" - SBA-15) recouvertes d'une bicouches lipidique.
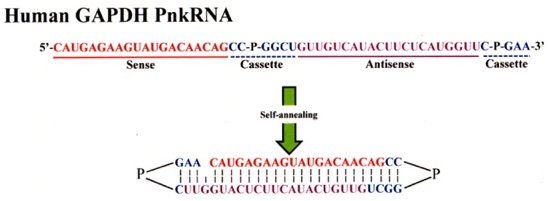
Source : Ashley et al. (2012) Elles sont 10 à 100 fois plus stables que les nanoparticules à base de bicouches lipidiques. Les protocellules chargées en siRNA se fixent spécifiquement aux cellules via un peptide de ciblage. b. La synthèse d'oligomères adoptant une structure résistante à la dégradation Les auteurs ont synthétisé un oligonucléotide de 51 nucléotides (correspondant à une partie de la séquence de l'ARNm de la glycéraldehyde 3-phosphate déshydrogénase - GAPDH - de l'homme) avec 2 prolines.
Source : Hamasaki et al. (2012) Cet oligomère avec 2 résidus proline s'autohybride et la structure obtenue, appelée agent RNAi PnkRNA™, correspond à :
Un autre oligonucléotide de 62 nucléotides (GAPDH humaine) et qui s'autohybride aussi a été synthétisé : agent RNAi nkRNA®. c. La synthèse assistée par ordinateur ("small interfering RNA design") La synthèse assistée par ordinateur s'appuie sur :
La conception de courts ARN interférant simple brin ("single-stranded short interfering RNA" - ss-siRNA) semble une voie prometteuse pour une formulation efficace dans l'optique d'une administration systématique : en effet, une étude de 2012 sur la souris a montré que le brin passager ne serait pas nécessaire pour une interférence ARN efficace (Lima et al., 2012). L'action du brin guide nécessitant un 5' phosphate, les auteurs de cette étude ont développé un ARN interférant simple brin (chargé sur AGO2) métaboliquement stable avec un 5'-(E)-vinylphosphonate. Autres exemples d'application :
|
| 12. Liens Internet et références bibliographiques |
![]()