| Les déshydrogénases à NAD+ ou NADP+ |
| Tweet |
|
|
1. Les familles de déshydrogénases 2. Les déshydrogénases à NAD+ ou NADP+
3. Structure du domaine liant le NAD(P)+ : le pli Rossmann |
4. Motif β1-αA-β2 de la lactate-DH lié au NAD+ 5. Mécanisme de transfert de H+ de l'éthanol à la nicotinamide (alcool-DH) 6. Représentation de la poche qui accomode la nicotinamide et stéréoisomérie du groupement carboxamide 7. Séquence des acides aminés du motif β1-αA-β2 de l'alcool, de la lactate et de la G-3-P DH 8. La glutamate déshydrogénase EC 1.4.1.4 |
|
1. Les familles de déshydrogénases La base de données PFAM ("Protein families database of alignments and HMMs") contient plus de 16.700 familles de protéines (Avril 2018). En choisissant l'option "Keyword search" et en tapant "dehydrogenase", on obtient plusieurs centaines de résultats. Remarque : le groupe des déhydrogénases à NAD(P)+ (E. C. 1.4.1.X ) est un sous-ensemble du groupe des oxydoréductases. Les oxydoréductases ne contiennent pas le même nombre de membres, par exemple :
|
||||||||||||||||||
| 2. Les déshydrogénases à NAD(P)+ |
| Exemples de déshydrogénases (DH) à NAD+ ou NADP+ | Rôle ou voie métabolique |
Toutes les déshydrogénases avec NAD+ ou NADP+ comme accepteurs d'électrons (EC : Enzyme Commission) |
|
||
|
||
|
||
|
||
|
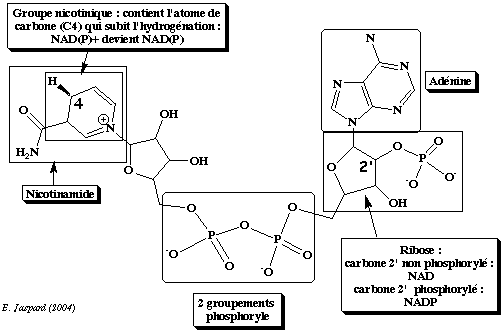
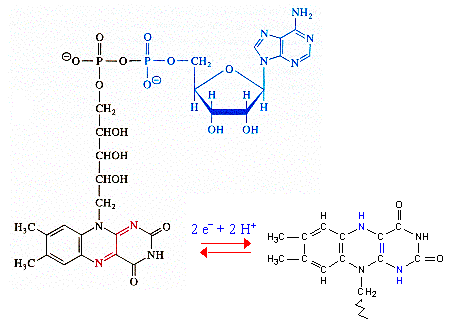
Les co-enzymes nicotinamide adénine dinucléotide NAD+ et NADP+ ne se distinguent que par un groupement phosphate sur le carbone 2' du ribose lié à l'adénine (figure ci-dessous).
Ils sont donc extrêmement proches du point de vue structural. Cependant :
Cette discrimination entre les deux formes du co-enzyme est un exemple admirable du pouvoir de reconnaissance des molécules par les enzymes en général et par les déshydrogénases en particulier. |
|
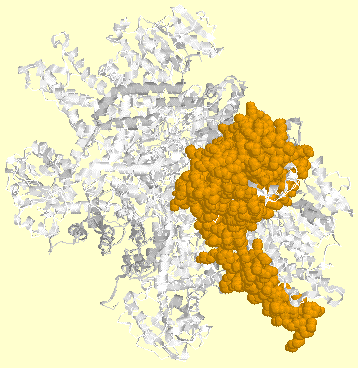
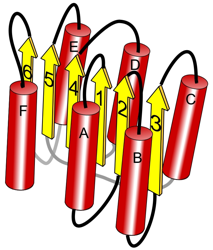
2a. Structure quaternaire des DH Les DH sont souvent des enzymes multimériques constituées de 2, 4 ou 6 sous-unités identiques. Ci-dessous la structure quaternaire (assemblage des sous-unités) de la glutamate DH. L'une des 6 sous-unités est en orange.
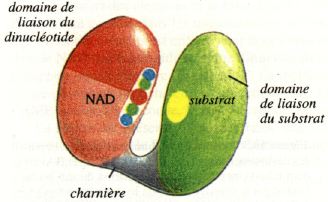
2b. Structure tertiaire des DH : les 2 domaines de l'alcool-DH NAD-dépendante Les séquences en acides aminés des DH n'ont que peu d'homologie. Les DH sont structurées en deux domaines :
Source : "Introduction à la structure des protéines" Branden & Tooze (1996) - ed. De Boeck Université
Ces 2 domaines sont reliées par une région dite "charnière" (hinge-bending") qui assure la souplesse conformationnelle nécessaire à l'activité catalytique. Le rapprochement des 2 domaines exclue les molécules de solvant lors de la catalyse. Le centre actif (site de fixation et site catalytique) est constitué par les acides aminés du sillon inter-domaine. |
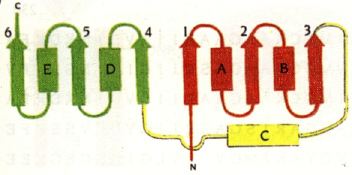
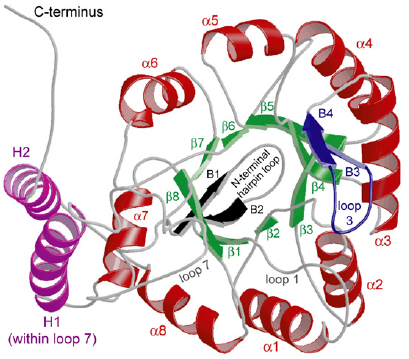
3. Structure du domaine liant le NAD(P)+ : le pli Rossmann Voir un rappel sur les structures secondaires. Le pli Rossmann ("Rossmann fold") est une structure super-secondaire (assemblage de plusieurs types de structures secondaires) : il est composé de 3 feuillets β liés à 2 hélices α de manière alternée (β-α-β-α-β). Un pli Rossmann peut fixer un nucléotide : le domaine de fixation d'un dinucléotides (tel que NAD+ ou NADP+) contient donc 2 plis Rossmann appariés, chacun d'eux fixant l'un des nucléotides du co-facteur. Le nom a été donné en hommage à Michael Rossmann.
Source : Branden & Tooze (1996)
|
|
Visualisation de la lactate DH de Squalus acanthias, non complexée au calcium, à une résolution de 3 Å Code PDB : 3LDH Seuls les acides aminés 20 à 83 sont représentés.
|
Les DH à flavine mononucléotide (FMN), appelées flavoenzymes, ne contiennent qu'un pli Rossmann. Figure ci-dessous :
Source : "Principes de Biochimie" Horton et al. (1994)
Le centre réactionnel est un noyau aromatique tricyclique, l'isoalloxazine (la partie en rouge). La FMN est une sorte de "convertisseur" d'un flux bi-électronique en un flux mono-électronique, puisqu'elle cède ses électrons à des centres [Fe - S], accepteurs mono-électroniques : +H+, +H- -H+ , -e- -H+ , -e- FMN -----------> FMNH2 ------------> FMNH. ------------> FMN |
Base de données :
Il existe des DH qui fixent le NAD(P)+ mais qui n'ont pas de pli Rossmann dans leur domaine de fixation du nucléotide. La structure est un tonneau (α/β)8 : 8 hélices et 8 feuillets qui s'alternent ("TIM-barrel fold").
Source : Richter et al. (2010) |
|
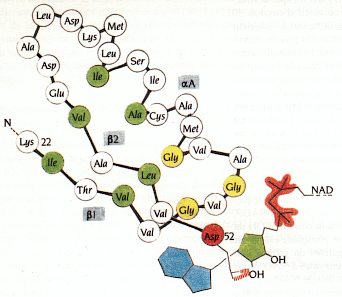
4. Motif β1-αA-β2 de la lactate-DH lié au NAD La présence de chaînes latérales hydrophobes à certaines positions (en vert sur la figure ci-dessous) est nécessaire pour l'empilement des brins β contre les hélices α.
Source : Branden & Tooze (1996) Le motif qui contient 31 résidus (de la lysine 22 à l'aspartate 52 en rouge) permet à cet aspartate de former une liaison hydrogène avec le 2'OH de l'adénosine. |
|
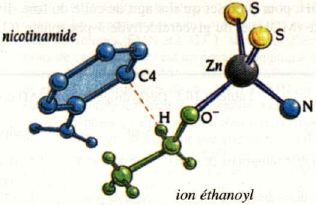
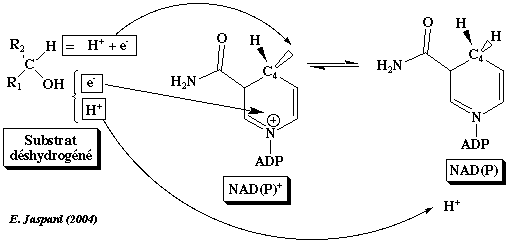
5. Mécanisme de transfert de H de l'éthanol à la nicotinamide (alcool-DH) La position spatiale du noyau nicotinamide du NAD+ et du substrat (l'éthanol dans cet exemple) explique la très haute stéréospécificité du transfert de l'ion hydrure sur le carbone 4 de la nicotinamide. Dans l'alcool DH, un atome de zinc du site catalytique établit une liaison avec la fonction alcool du substrat. Cette liaison s'ajoute à celles qui stabilisent de manière optimale le substrat dans le site catalytique et, en polarisant la liaison C-OH de la fonction alcool du substrat, elle facilite le transfert de l'atome d'hydrogène.
Source : Branden & Tooze (1996) Stéréospécificité du transfert de l'atome d'hydrogène La réaction est : NAD(P)+ + H2 <=> NAD(P)H Lors de la déshydrogénation (H2 = 2 H+ + 2 e-) du substrat, les transferts suivants s'opèrent :
|
|
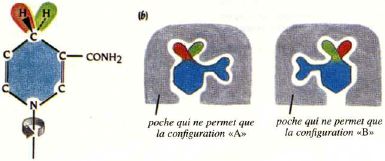
6. Représentation de la poche qui accomode la nicotinamide et stéréoisomérie du groupement carboxamide Les déshydrogénases sont divisées en 2 classes : A et B, en fonction de la stéréospécificité du transfert de l'atome d'hydrogène sur l'atome C4 de la nicotinamide. Le cycle est asymétrique à cause du substituant sur le C3 : le groupement carboxamide. En conséquence, les 2 atomes d'hydrogène sur le C4 dans la forme réduite (au-dessus et au dessous du cycle nicotinamide) ne sont pas équivalents.
Source : Branden & Tooze (1996)
L'explication structurale est que pour chaque classe de déshydrogénases, le groupement carboxamide est fixé dans une "poche" de sorte qu'une seule des faces du cycle nicotinamide est accessible lors du transfert de l'atome d'hydrogène. |
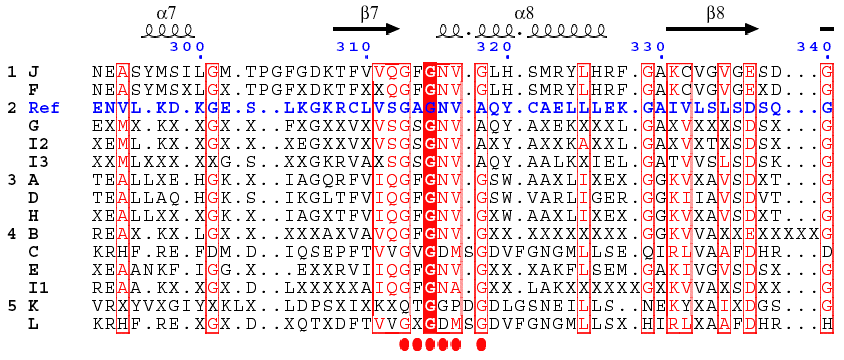
| 7. Séquence des acides aminés du motif β1-αA-β2 L'aspartate en rouge est conservé comme l'indique l'alignement ci-dessous : alcool, lactate et glycéraldéhyde-3 phosphate déshydrogénases.
Source : Branden & Tooze (1996)
Celà permet aux déshydrogénases de discriminer les deux formes du coenzyme. Les déshydrogénases à NADP+ (exemple : la thréonine déshydrogénase) ont pour leur part :
|
|
8. La glutamate déshydrogénase EC 1.4.1.4 A βαβ fold is found in the coenzyme-binding sub-domain (β7-α8-β8) of the three isoforms of glutamate dehydrogenases (EC 1.4.1.2, EC 1.4.1.3 and EC 1.4.1.4).
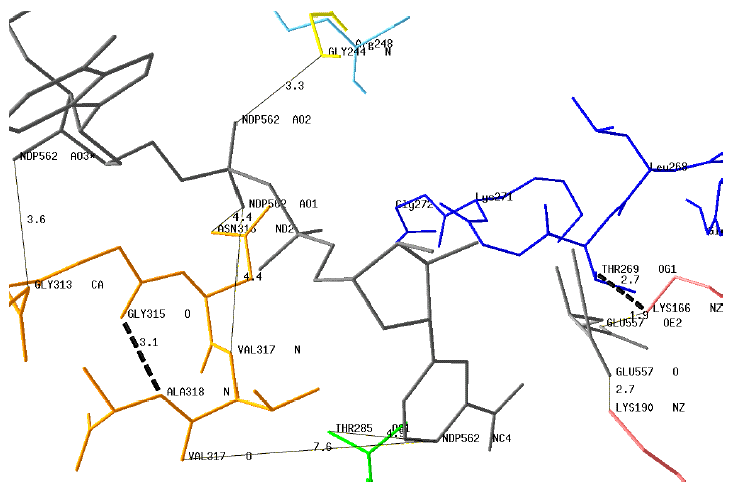
Source : Jaspard E. (2006) This Rossmann fold begins with the motif G313AGNVA318 in the case of plant GDH4 (Ref). By comparison, the motifs described in the literature are GXGXXG, GXGXXA and even GXGXXS as for example in the case of very large GDH from Streptomyces clavuligerus. Modeling of the coenzyme-binding motifs and key residues of GDH4 from Chlorella sorokiniana with NADPH (NDP562) and glutamate Some interactions (plain lines) between the motif G313AGNVA318 or key residues and the coenzyme are indicated.
Source : Jaspard E. (2006) The distances between the protonated carbon atom of the nicotinamide moiety (NDP562NC4) are too long for direct interactions with the motif G313AGNVA318. However, this motif is stabilized by the H-bond Gly315O - Ala318N (dotted line).
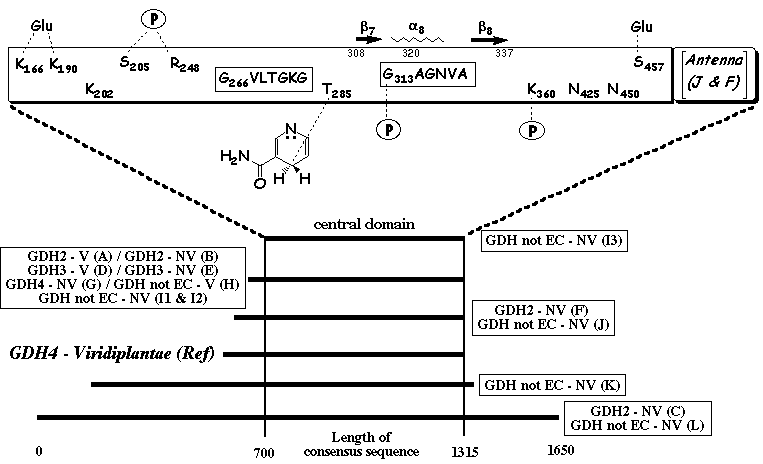
Global organization of GDH subunits All GDHs share a common pattern called the central domain flanked by an N-terminal region of various lengths and, for large GDHs, by a shorter C-terminal one. The upper scheme shows the structural features specific of the central domain from plant GDH4. The length indicated includes the gaps generated by the alignment.
Source : Jaspard E. (2006) |
![]()