| L'ADP-ribosylation |
| Tweet |
|
|
C'est une modification post-traductionnelle immédiate et transitoire des protéines du noyau. Elle est la réponse de la cellule aux interruptions du squelette sucre-phosphate de l'ADN créées par des agents génotoxiques :
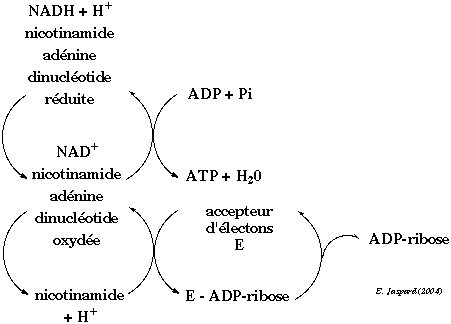
Les protéines qui subissent ce type de modification post-traductionnelle (histones, lamine B et enzymes du métabolisme de l'ADN) sont pour la plupart associées à la chromatine. Exemple de processus physiologiques : fonctionnement des adypocytes, fonctionnement de la topoisomèrase II, différentiation cellulaire, ... Une fois poly-ADP-ribosylées, les protéines perdent leur affinité pour l'ADN. Le groupement ADP-ribose provient de la nicotinamide adénine dinucléotide (NAD) sous sa forme oxydée. La forme oxydée et réduite du NAD sont elles-mêmes en interconversion permanente et en relation avec la synthèse d'ATP. 3 types d'enzymes sont impliquées dans le métabolisme de l'ADP-ribose :
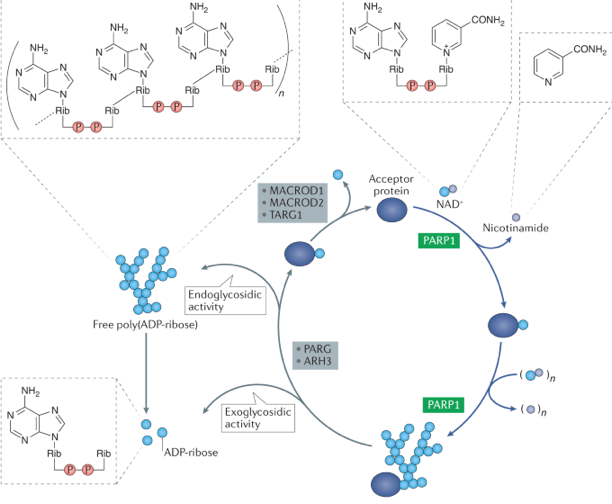
La poly-ADP-ribose polymérase 1 (PARP1, E.C. 2.4.2.30) catalyse la synthèse du PAR à partir du NAD+ : PARP1 clive le NAD+ en ADP-ribose et nicotinamide et attache de manière covalente l'ADP-ribose à une protéine accepteur. Des molécules de NAD+ supplémentaires peuvent être clivées et constituer des formes linéaires et ramifiées du PAR.
Source : Curtin & Szabo (2020)
|
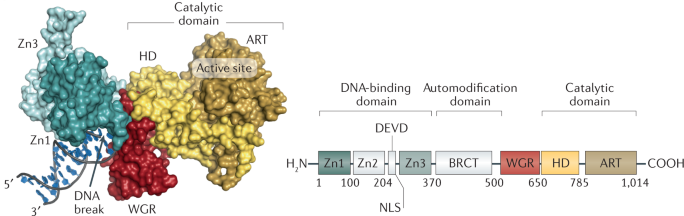
Domaines structuraux de PARP1 La poly-ADP-ribose polymérase contient 3 domaines structuraux : le domaine de fixation à l'ADN, le domaine d'auto-modification et le domaine de fixation du NAD+ (domaine catalytique).
Source : Curtin & Szabo (2020) A gauche : la surface de la liaison de PARP1 à l'ADN endommagé. A droite : organisation des domaines de PARP1 de l'homme.
|
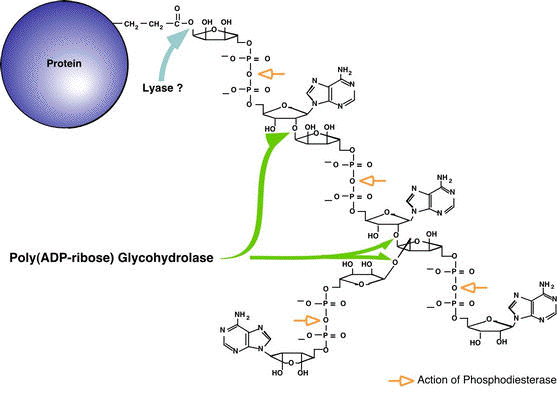
Structure schématique du poly-(ADP-ribose) liée à une protéine et sites de clivage
Source : "ADP-ribose polymer metabolism" - Amé, Jacobson & Jacobson Deux enzymes sont impliquées dans la dégradation du poly-(ADP-ribose) :
|
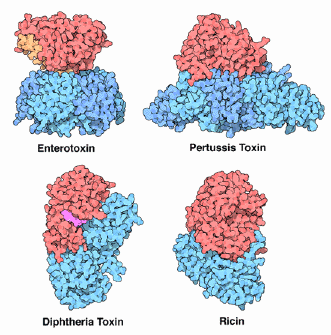
Mode d'action des toxines de type A-B Les bactéries pathogènes utilisent leurs toxines pour modifier ou détruire les cellules hôtes. Les toxines bactériennes à activité ADP-ribosyle forment une grande famille de toxines mortelles (voir tableau ci-dessous). Ces toxines sont produites par un large éventail de bactéries pathogènes et constituent l'agent cytotoxique qui cause de graves maladies infectieuses (coqueluche, choléra, diphtérie, ...). |
| Caractéristiques des toxines à activité ADP-ribosyle de plusieurs souches bactériennes virulentes | |||
| Toxines bactériennes à activité ADP-ribosyle | Bactérie | Cible | Effets pathologiques |
| toxine de la coqueluche | Bordetella pertussis | Cystéine des sous-unités α de la sous-famille Gαi (Gαi, Gαo et Gαt) sauf Gαz | Le couplage [protéine Gαi -récepteur] est inhibé et la transduction du signal est bloquée |
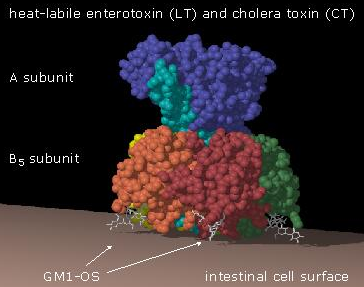
toxine du choléra enterotoxine labile à la chaleur |
Vibrio cholerae Escherichia coli |
Arginines des sous-unités α de la sous-famille Gαs (Gαs et Gαolf) | L'activité GTPase de la sous-unité stimulante Gαs est inhibée : la sous-unité Gαs est activée en permanence |
toxine diphtérique exotoxine A |
Corynebacterium diphtheirae Pseudomonas aeruginosa |
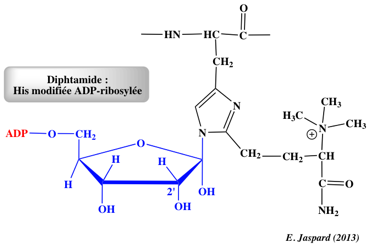
Le facteur 2 d'élongation de la traduction chez les Eukaryotes (eEF2) contient une histidine modifiée appelée diphthamide qui est la cible de la toxine
|
La synthèse des protéines est bloquée |
|
Sources : PDB et "The cholera toxin family : action and inhibition" Codes PDB : toxine du choléra : 1XTC; enterotoxine : 1LTB; toxine de la coqueluche : 1PRT; toxine diphtérique : 1MDT |
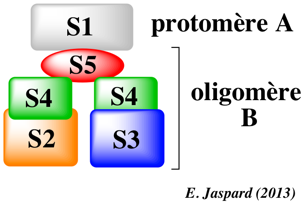
La toxine de la coqueluche (PTX) est une toxine de type A-B : ces toxines sont constituées de deux parties (A et B) qui jouent des rôles différents dans l'action de la toxine. PTX modifie les réponses cellulaires via au moins 2 voies de signalisation différentes :
L'ADP-ribosylation de la sous-unité α de la protéines Gi/o verrouille cette sous-unité dans un état inactif (forme GDP fixé) : elle ne peut plus inhiber l'adénylate cyclase. Cela entraîne l'accumulation d'AMPc ce qui induit divers effets pathologiques dans les cellules hôtes. La sous-unité S1 (protomère A) contient 2 cystéine en positions 41 et 201, qui forment un pont disulfure dans l'holotoxine native. La réduction de ce pont disulfure induit une forte stimulation de l'activité catalytique de la sous-unité S1, concomitante avec le relarguage de la sous-unité S1 de l'oligomère B. L'extrémité C-terminale de la sous-unité S1 de PTX (composée de 235 acides aminés) est importante dans l'interaction de cette sous-unité S1 avec l'oligomère B (notamment les acides aminés 220 à 235 hydrophobes). Les acides aminés 195 à 204 sont requis pour l'ADP-ribosylation de la sous-unité α de la protéines Gi/o. Les acides aminés 205 à 219 constituent la région catalytique de la sous-unité S1 et sont le site de fixation de l'oligomère B. Les séquences en acides aminés des sous-unités S2 et S3 ont environ 70% d'homologie. Cependant, la sous-unité S2 se fixe aux glycanes non-sialylés et la sous-unité S3 se fixe aux glycanes sialylés. |
|
Erener et al. (2012) "Poly(ADP-ribose)polymerase-1 (PARP1) controls adipogenic gene expression and adipocyte function" Mol. Endocrinol. 26, 79 - 86 Meyer-Ficca et al. (2011) "Poly(ADP-ribose) polymerases PARP1 and PARP2 modulate topoisomerase II beta (TOP2B) function during chromatin condensation in mouse spermiogenesis" Biol. Reprod. 26, 900 - 909 Curtin & Szabo (2020) "Poly(ADP-ribose) polymerase inhibition: past, present and future" Nat. Rev. Drug Discov. 19, 711 - 736 |
![]()