Les sirtuines |
| Tweet |
|
|
1. Introduction 2. Structure et mécanisme catalytique des sirtuines 3. La sirtuine 1 et le resvératrol |
4. Les sirtuines mitochondriales 5. La sirtuine 5 6. Liens Internet et références bibliographiques |
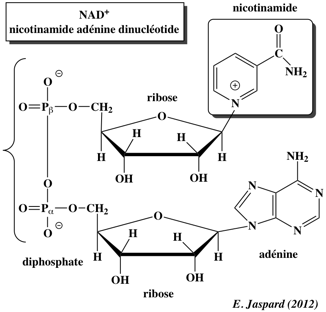
Introduction L'acétylation des lysines est une modification post-traductionnelle très importante. Par exemple, l'acétylation et la désacétylation ont une incidence sur la structure de la chromatine et donc jouent un rôle capital dans la régulation de la transcription des gènes. Voir un cours sur l'épigénétique. Les sirtuines sont des enzymes qui forment une famille conservée d'histones désacétylases nictotinamide adénine dinucléotide (NAD)-dépendantes et d'ADP-ribosyltransférases.
Elles participent à la régulation de nombreuses voies biologiques, et semblent jouer un rôle primordial dans les effets liés au vieillissement. Le nom sirtuines (aussi appelées "Silent mating type information regulation 2 " ou protéines "Sir2") vient du gène "Silent Information Regulation 2" de la levure, impliqué entre autre dans la répression de la transcription.
|
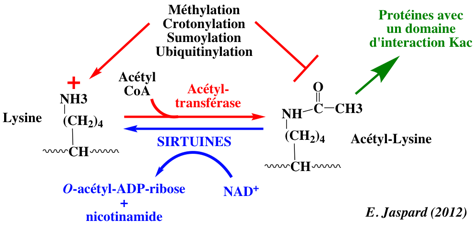
On trouve les sirtuines dans tous les règnes. Remarque : on a montré (Du et al., 2011) que les sirtuines 5 sont des désuccinylases et démalonylases et non des désacétylases (voir le paragraphe "Sirtuine 5"). On constate un intérêt grandissant pour les sirtuines en particulier à cause du rôle capital des espèces réactives de l'oxygène ("Reactive Oxygen Species" - ROS) dans certaines maladies des mammifères (notamment la longévité). Les sirtuines des rôles importants également dans la régulation du métabolisme. On a dénombré 7 sirtuines chez les mammifères: Sirt1 à Sirt7. Ces sirtuines ont un domaine catalytique commun et des extrémités N- et C-terminales différentes. Ces extrémités sont la cible de modifications post-traductionnelles qui modulent les fonctions des sirtuines. Les lysines acétylées fournissent des sites d'empilement aux protéines qui possèdent un domaine d'interaction "lysines acétylées" (Kac), c'est-à-dire un bromodomaine.
|
| Quelques caractéristiques des sirtuines | ||||||||
| Classe | Sous-classe | Bactéries | Levure | Souris | Homme | Localisation | Activité | Exemples de fonctions |
| I | a |
Sir2 ou Sir2p |
Sirt1 | hSIRT1 | Noyau - cytoplasme |
Désacétylase (inhibée par la nicotinamide - activée par le resvératrol) |
Apoptose - inflammation | |
| b | Hst2 or Hst2p | Sirt2 | hSIRT2 | Cytoplasme | Désacétylase | Cycle cellulaire | ||
| Sirt3 | hSIRT3 | Noyau - mitochondrie | Désacétylase | Métabolisme | ||||
| c | Hst3 ou Hst3p |
|||||||
| II | Sirt4 | hSIRT4 | Mitochondrie | ADP-ribosyltransférase (EC 2.4.2) | Sécrétion d'insuline | |||
| III | Sirt5 | hSIRT5 | Mitochondrie | Désuccinylases et démalonylases | Détoxification de l'ammoniaque | |||
| IV | a | Sirt6 | hSIRT6 | Noyau | ADP-ribosyltransférase et désacétylase | Métabolisme - réparation de l'ADN |
||
| b | Sirt7 | hSIRT7 | Nucléole | Désacétylase ? | Régulation de la transcription par l'ARN polymérase I | |||
| U : procaryotes | cobB | Régulation de l'acétyl-CoA synthétase - désacétylase | Métabolisme | |||||
| Source principale : Wikipedia | ||||||||
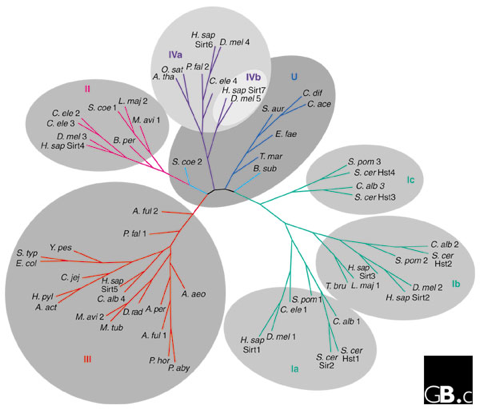
Figure ci-dessous : arbre phylogénétique de 60 sirtuines.
Source : Frye (2000) et North & Verdin (2004) |
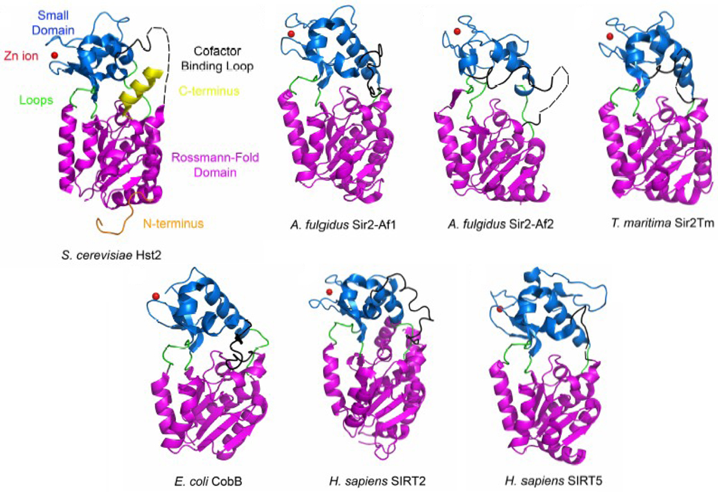
2. Structure et mécanisme catalytique des sirtuines Les sirtuines ont un domaine catalytique (environ 275 acides aminés) dont la structure est très conservée (figure ci-dessus). En revanche, les extrémités N- et C-terminales ont des séquences distinctes, des longueurs différentes et des contenus en structures secondaires différents. Figure ci-contre : comparaison de structures de sirtuines d'origines diverses.
Source : Sanders et al. (2010) |
Domaine Rossmann ("Rossmann-Fold Domain ") : domaine de fixation du NAD+. Le petit domaine ("small domain") de fixation du zinc (petite sphère rouge) est le plus variable en terme de séquence en acides aminés. Il est constitué de 3 feuillets β anti-parallèles et d'1 hélice α. Il contient la séquence consensus de fixation du zinc : Cys-X(2,4)-Cys-X(15,40)-Cys-X(2,4)-Cys. La diversité de structures de ce petit domaine suggère qu'il puisse jouer un rôle dans la discrimination des substrats des sirtuines. Le zinc a un rôle structural. Les boucles en vert ("Loops") qui joignent le petit domaine et le domaine Rossmann forment le site actif où se fixent le NAD+ et les protéines lysine-acétylées. C'est la région qui a le plus d'homologie entre les séquences de sirtuines. La boucle qui fixe le NAD+ est désordonnée (ligne pointillée) en absence du cofacteur et se structure quand elle le fixe. Le NAD+ adopte une conformation dite "étendue". Il se fixe en sens inverse du sens habituel des déshydrogénases à NAD+ : l'adénine se fixe sur la partie C-terminale du feuillet β et le groupement nicotinamide sur la partie N-terminale. Le NAD+ établit des interactions directes ou médiées par des molécules d'eau avec : Ala33, Gly34, Ser36, Thr37, Ile41, Asp43, Phe40, Phe44, Tyr52, Trp58, Gln115, Asn116, Ile117, Asp118, His135, Gly223, Val228. Les résidus Ala33, Gly34, Thr37, Phe44 et Asp43 forment la poche dite "C" ("C-pocket") où se fixe le noyau nicotinamide du NAD+. Cette poche C joue un rôle important dans le mécanisme catalytique. La fixation du substrat protéine lysine-acétylée induit un déplacement significatif du petit domaine par rapport au domaine Rossmann, ces 2 domaines adoptant alors une conformation dite en "agraphe β" ("β staple motif") qui forme le site de fixation de l'acétyl-Lysine. La partie aliphatique de la chaîne latérale de l'acétyl-Lysine établit de multiples contacts de van der Waals avec les acides aminés conservés His135, Phe184, Leu188 et Val228. Le groupement acétyle établit des interactions avec Ile117, H135 et Val182. Les acides aminés du motif "agraphe β" et ceux qui interagissent avec l'acétyl-Lysine sont hautement conservés. Les extrémités N- et C-terminales ont été moins bien résolues dans les structures 3D disponibles. Les 7 acides aminés de l'extrémité N-terminale de Hst2 semblent impliqués dans la régulation de l'oligomérisation (trimérisation) de cette sirtuine. L'extrémité C-terminale interviendrait dans la modulation de la fixation du NAD+ (mécanisme d'autorégulation). |
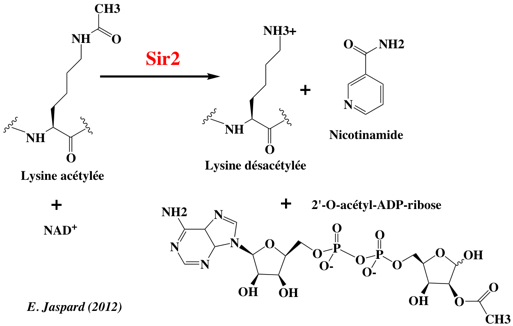
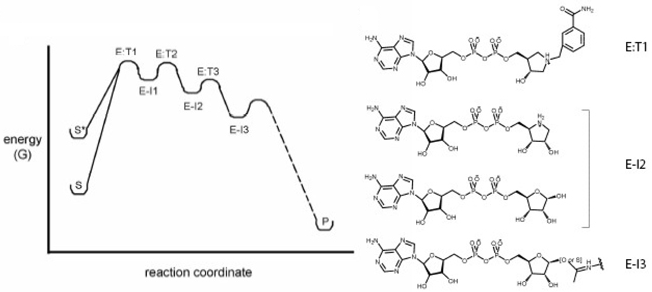
Le mécanisme catalytique des sirtuines, décrypté avec des analogues de l'état de transition, propose 3 complexes [intermédiaire/état de transition] (figure ci-dessous).
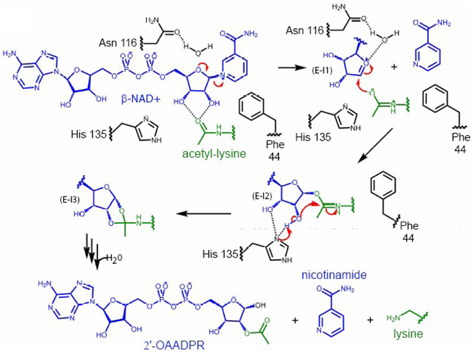
E:T1, E:T2 et E:T3 : états de transition requis pour former les intermédiaires E-I1, E-I2 et E-I3. Source : Sanders et al. (2010) Certains de ces complexes ont été cristallisés apportant des données supplémentaires quant à la stéréospécificité de ce mécanisme. La liaison glycosidique est rompue (dissociation de type SN1 ou SN2), formant l'intermédiaire ion oxocarbenium (E–I1, figure ci-dessus et détail figure ci-dessous) accompagné d'un changement de conformation du cycle du ribose lié à la nicotinamide. L'acétyl-Lysine est alors positionnée pour effectuer une attaque nucléophile du carbone C1' de l'intermédiaire ion oxocarbenium et former l'intermédiaire alkylamidate (E–I2), accompagné de nouveau d'un changement de conformation du cycle du ribose.
Source : Sanders et al. (2010) Suivent plusieurs étapes dont, notamment, l'addition d'une molécule d'eau. Les produits de la réaction (Lysine, nicotinamide et 2'-O-acétyl-ADP-ribose) sont enfin formés.
|
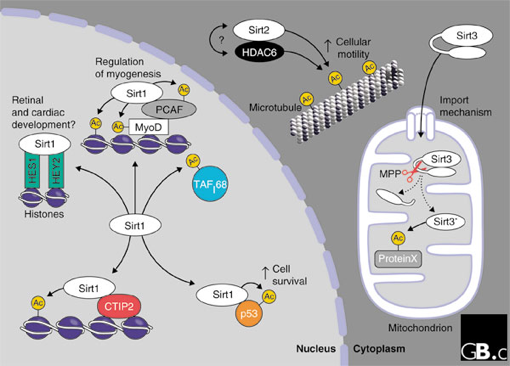
3. La sirtuine 1 et le resvératrol Sirt1 est située dans le noyau. C'est une protéine désacétylase NAD+-dépendante qui établit un lien direct entre la régulation de la transcription et le niveau énergétique de la cellule. Elle joue donc un rôle de senseur du rapport des concentrations [NAD+/NADH] cytosoliques, rapport modulé par le taux de glucose et les changements métaboliques associés à une restriction calorique. En effet, elle est un composant du complexe appelé "energy-dependent nucleolar silencing" (eNoSC). Le complexe eNoSC est un senseur du niveau énergétique de la cellule: lors d'un déficit en glucose, l'élévation du rapport des concentrations [NAD+/NADH] active Sirt1 qui désacétyle les histones H3 (modulation du fonctionnement de la chromatine), suivi par leur diméthylation sur la Lys9 (H3K9me2) par l'histone-lysine N-méthyltransférase SUV39H1 (répression de la transcription). Voir un cours sur l'épigénétique. Par ailleurs, Sirt1 désacétyle de trés nombreux facteurs de transcription, activant et réprimant ainsi l'expression de nombreux gènes
Source : North & Verdin (2004) Autres rôles de Sirt1 :
|
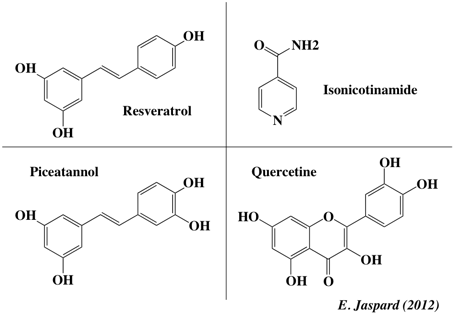
Le resvératrol (figure ci-dessous) est une phytoalexine (polyphénols de la classe des stilbènes) que l'on trouve dans de nombreuses plantes supérieures (notamment le raisin, l'arachide, la rhubarbe, …). Il augmente l'expression d'un microARN (miR-663) dans les cellules immunitaires humaines, diminuant ainsi l'expression du microARN miR-155 qui favorise la réaction inflammatoire. Figure ci-dessous : différents activateurs de Sirt1.
Le resvératrol a un effet cardioprotecteur en inhibant l'agrégation des plaquettes et en atténuant l'expression de la P-selectine 35. Cet effet serait lié à la diminution de l'activité de la phospholipase C. Le resvératrol neutralise les radicaux libres (et autres oxydants) et inhibe l'oxydation des lipoprotéines LDL. |
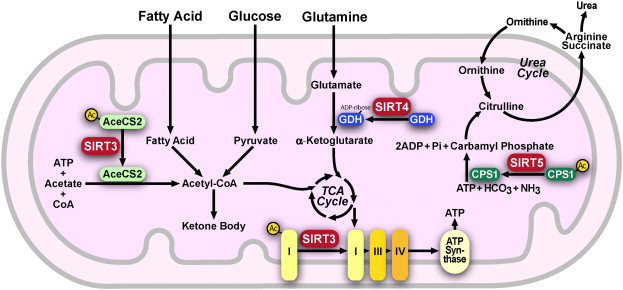
4. Les sirtuines mitochondriales Sirt3 désacétyle et active l'acétyl-CoA synthétase 2 (AceCS2) afin d'augmenter la production d'acétyl-coA.
Figure ci-dessous : Régulation des enzymes mitochondriales par Sirt3, Sirt4 et Sirt5 dans la matrice mitochondriale.
Source : Huang et al. (2010) La sur-expression de Sirt3 augmente la respiration et diminue la production de ROS. Sirt4 inhibe la glutamate déshydrogénase ce qui induit une diminue la sécrétion d'insuline. |
|
Ce travail souligne un point capital : l'annotation manuelle est une activité trés chronophage et qui requiert de vastes connaissances encyclopédiques. Elle est cependant irremplaçable car cette expertise humaine n'a pas d'équivalent. Elle n'est pas, à ce jour, supplantée par l'annotation automatique par des logiciels. L'activité désacétylase a été démontrée pour Sirt1, Sirt2 et Sirt3 des mammifères. Cette annotation a été propagée aux paralogues sur la base de la similarité entre les séquences. Cependant, Sirt4 à Sirt7 ne possèdent qu'une trés faible activité désacétylase, voire pas du tout. Une avancée majeure a été l'obtention de données structurale de la sirtuine 5, Sirt5. La poche de fixation du groupement acétyle est beaucoup plus grande chez Sirt5 que chez Sirt2 et elle peut fixer un acyl-CoA acyle chargé négativement à la place. Dans la cellule, les formes acyl-CoA avec un groupement carboxylate les plus courantes sont le malonyl-CoA et le succinyl-CoA. Sirt5 catalyse leur hydrolyse : c'est donc une démalonylase et une désuccinylase et non une désacétylase. La préférence pour les groupements succinyl et malonyl s'explique par la présence de l'Arg105 et de la Tyr102 dans la poche acyle de Sirt5. La délétion de Sirt5 chez la souris augmente le taux de succinylation de CPS1. En conséquence, la succinylation des lysines est une modification post-traductionnelle qui peut être réversée par Sirt5 . Les termes lysine-succinylation et lysine-malonylation ont été introduits dans le vocabulaire contrôlé des modifications post-traductionnelles de UniProtKB ("controlled vocabulary of posttranslational modifications"). |
|
Visualisation de la sirtuine 5 de Homo sapiens à une résolution de 1,55 Å (2011). PDB : 3RIY Complexe avec un peptide histone 3 (N-succinyl lysine).
|
| 6. Liens Internet et références bibliographiques |
|
"Principes de Biochimie" Horton, Moran, Ochs, Rawn et Scrimgeour (1994) - Ed. DeBoeck Universités - ISBN : 2-8041-1578-X |
|
| "Sirtuins and Oxidative Stress" (SA-Biosciences - Qiagen)
"NRF2-Mediated Oxidative Stress Response" (SA-Biosciences - Qiagen) "Resveratrol" |
|
|
Frye RA (2000) "Phylogenetic Classification of Prokaryotic and Eukaryotic Sir2-like Proteins" BBRC 273, 793 - 798 North & Verdin (2004) "Sirtuins: Sir2-related NAD-dependent protein deacetylases" Genome Biology 2004, 5:224 Murayama et al. (2008) "Epigenetic Control of rDNA Loci in Response to Intracellular Energy Status" Cell 133, 627 - 639 Bosch-Presegué et al. (2011) "Stabilization of Suv39H1 by SirT1 Is Part of Oxidative Stress Response and Ensures Genome Protection" Molecular Cell 42, 210 - 223 |
|
|
Huang et al. (2010) "Mitochondrial sirtuins" Biochim. Biophys. Acta 1804, 1645 - 1651 Hirschey et al. (2010) "SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation" Nature 464, 121 - 125 Nakagawa et al. (2010) "SIRT5 Deacetylates Carbamoyl Phosphate Synthetase 1 and Regulates the Urea Cycle" Cell 137, 560 - 570 Du et al. (2011) "Sirt5 is a NAD-dependent protein lysine demalonylase and desuccinylase" Science 334, 806 - 809 |
|
|
Sanders et al. (2010) "Structural Basis for Sirtuin Function: What We Know and What We Don't" Biochim Biophys Acta 1804, 1604 - 1616 Flick & Lüscher (2012) "Regulation of Sirtuin Function by Posttranslational Modifications" Front Pharmacol. 3, 29 |
|
![]()