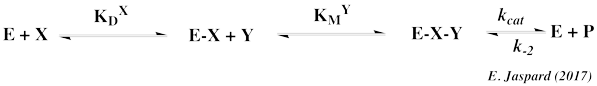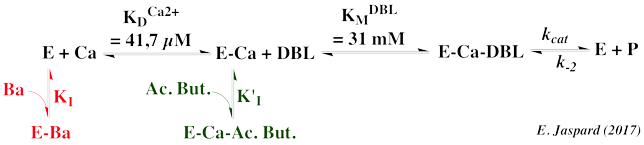| Enzymologie : Réactions enzymatiques à deux substrats |
| Tweet |
|
| Voir le cours sur les cinétiques à 2 substrats. |
| Voir les énoncés des exercices. |
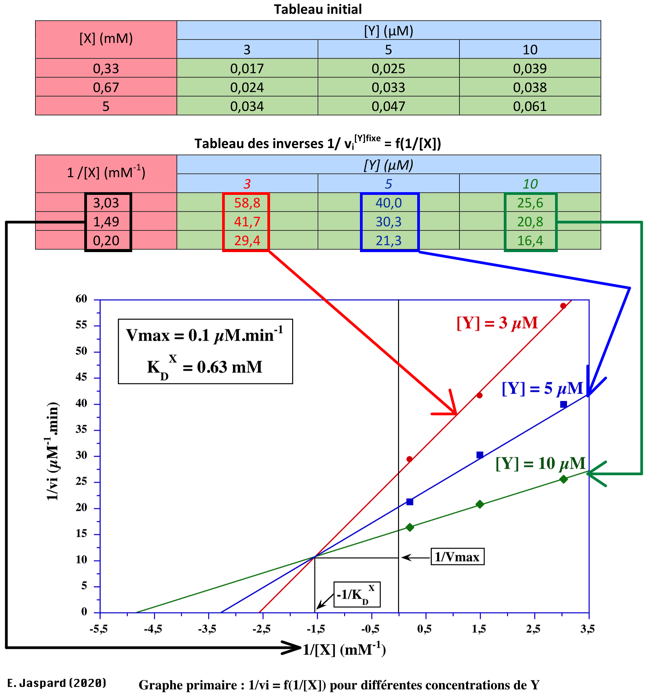
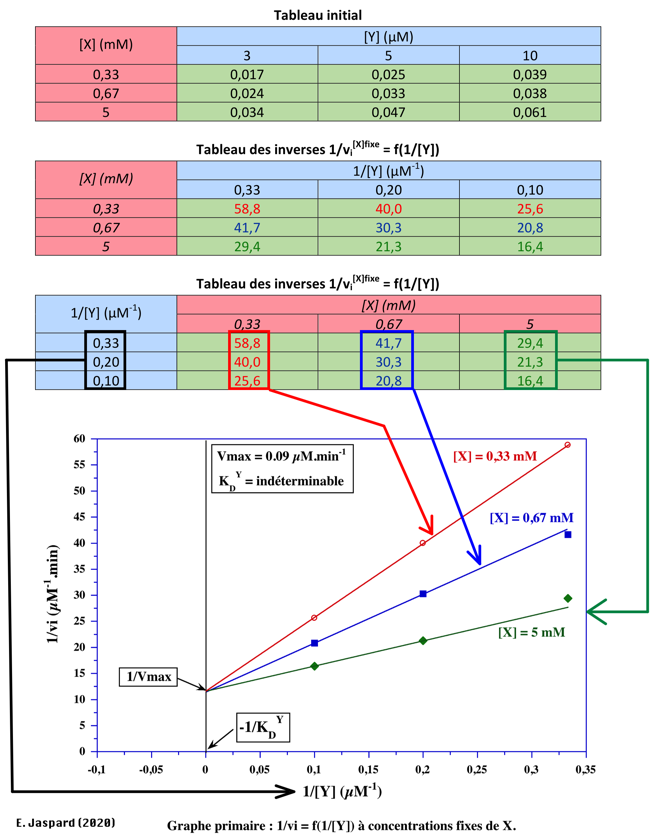
Exercice N°1 - mécanisme ordonné
1/VMax = 10 µM-1.min => VMax = 0,1 µM.min-1 1/KDY = 0 (point de concourrance sur l'axe des ordonnées) => KDY= infini => Y ne peut pas se fixer sur l'enzyme libre. C'est donc un mécanisme ordonné avec Y en second.
|
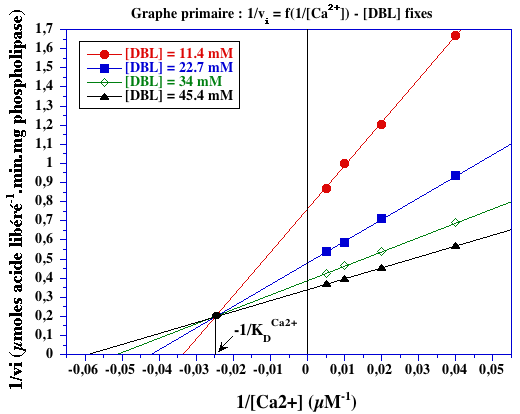
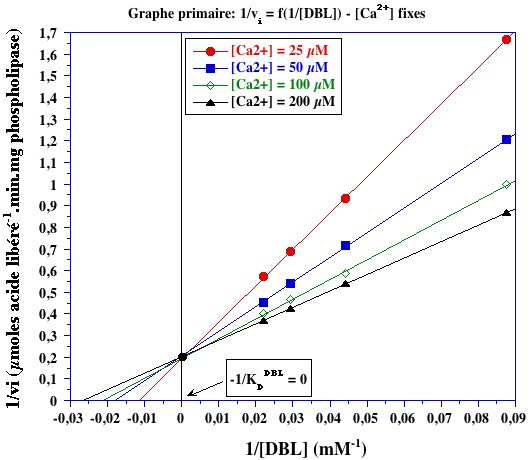
Exercice N°2 : Mécanisme ordonné calcium - DBL et inhibiteurs 1. Graphes primaires
KDCa2+ = 41,7 µM
1/KDDBL = 0 (point de concourrance sur l'axe des ordonnées) => KDDBL = infini => DBL se fixe en second. |
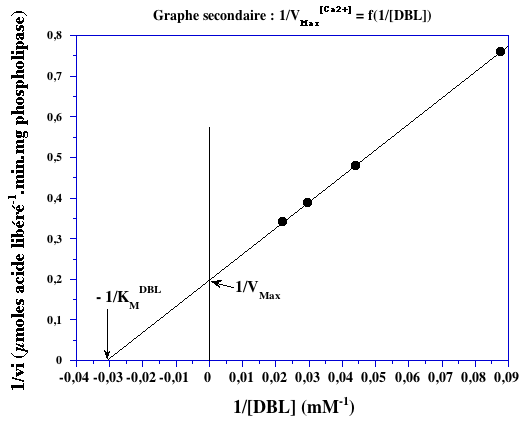
2. Graphe secondaire A partir du graphe primaire pour le calcium (ci-dessus), on obtient les valeurs du tableau ci-dessous qui permettent de tracer le graphe secondaire pour la DBL (ci-dessous) :
1/VMax = 0,2 µmol-1.min.mg et -1/KMDBL = 0,032 mM-1 => VMax = 5 µmol.min-1.mg-1 et KMDBL = 31 mM. Il s'agit d'un mécanisme ordonné.
|
| 3. Effet des inhibiteurs : baryum et acide butyrique |
| inhibiteur | 1er substrat : Ca2+ | 2ème substrat : DBL |
| baryum | Compétitif pour le Ca2+ : fixation sur la forme libre E | Puisque compétitif pour le Ca2+, alors forcément compétitif pour DBL |
| acide butyrique (Ac. but.) | Incompétitif : fixation sur E-Ca (qui est une forme ES) | Compétitif : fixation de Ac. but. sur E-Ca qui est une forme libre pour DBL |
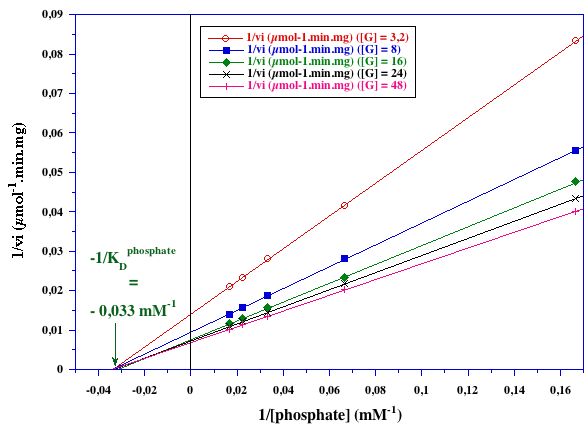
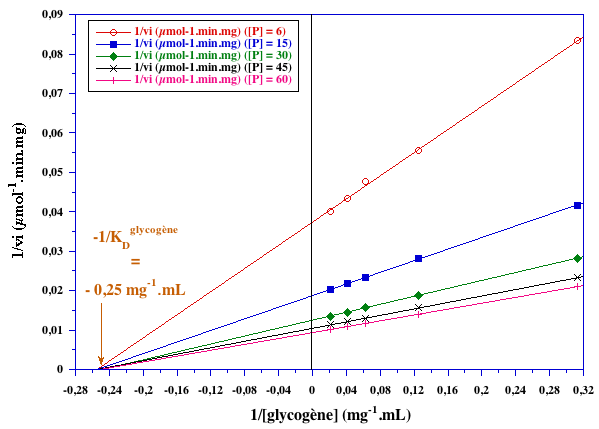
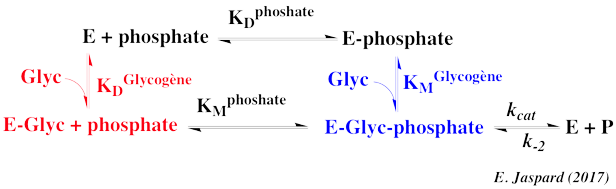
Exercice N°3 : Mécanisme au hasard - glycogène - phosphate 1. Graphes primaires
KDphosphate ≈ 30 mM
KDglycogène = 4 mg.ml-1 |
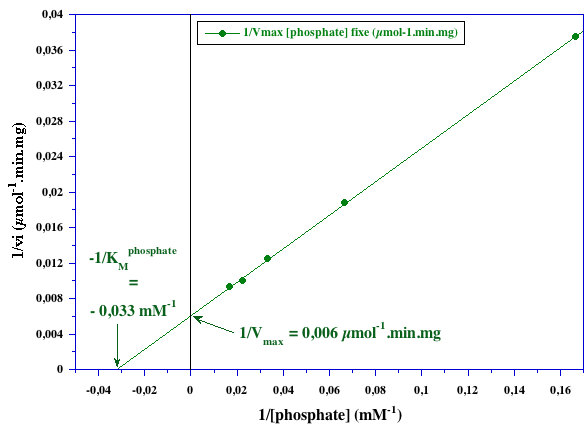
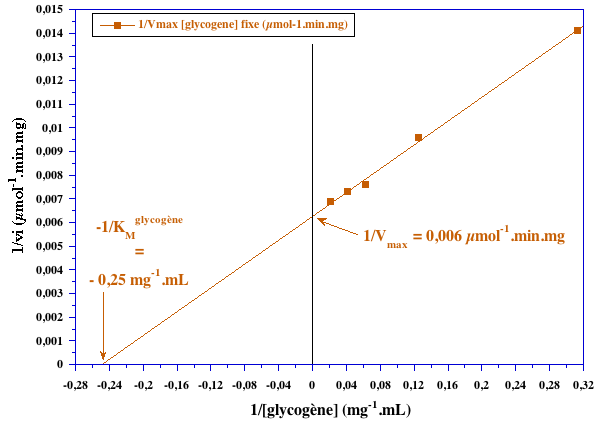
2. Graphes secondaires |
| valeurs issues du graphe primaire pour le phosphate | valeurs issues du graphe primaire pour le glycogène | ||
| [glycogène] (mg.mL-1) | 1/Vmax[glycogène] fixe (µmol-1.mn.mg) | [phosphate] (mM) | 1/Vmax[phosphate] fixe (µmol-1.mn.mg) |
| 3,2 | 0,0141 | 6 | 0,0375 |
| 8 | 0,0096 | 15 | 0,0188 |
| 16 | 0,0076 | 30 | 0,0125 |
| 24 | 0,0073 | 45 | 0,0100 |
| 48 | 0,0069 | 60 | 0,0094 |
VMax ≈ 167 µmoles.min-1.mg-1 ; KMphosphate ≈ 30 mM ; KMglycogène = 4 mg.ml-1 Il s'agit d'un mécanisme réactionnel au hasard.
Il s'agit d'un schéma carré puisque KDphosphate = KMphosphate et KDglycogène = KMglycogène. |
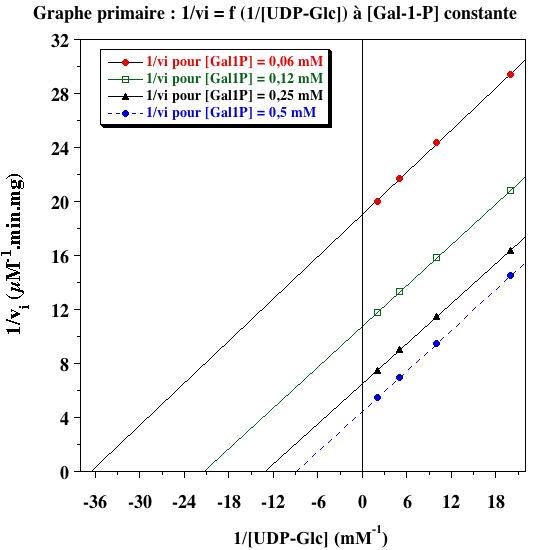
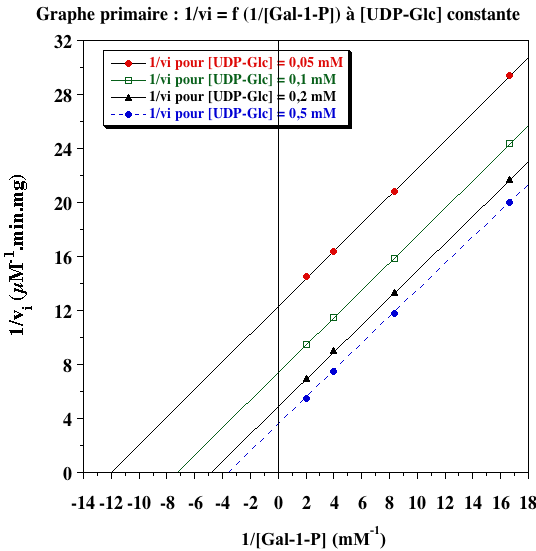
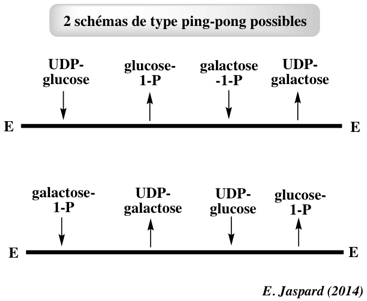
Exercice N°5 - mécanisme ping-pong 1. Graphes primaires
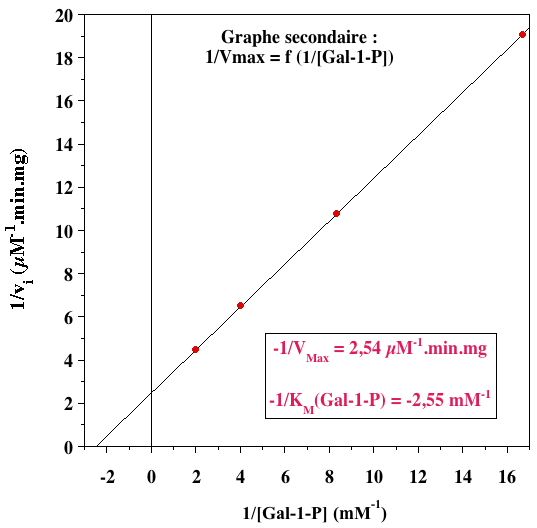
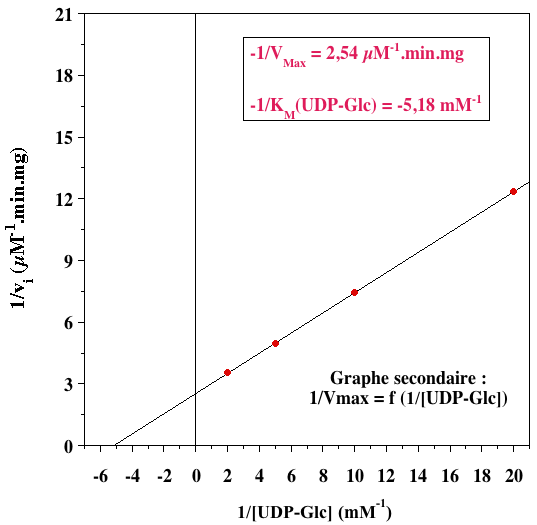
L'ensemble de droites parallèles indique un mécanisme ping-pong. 2. Graphes secondaires
VMax= 0,39 µM.min-1.mg-1 |
| Effet des inhibiteurs (voir l'énoncé) | ||
| UDP-glucose | Galactose-1-P | |
| UDP-galactose | Compétitif | Non compétitif |
| Glucose-1-P | Non compétitif | Compétitif |
Le schéma du bas de la figure ci-dessous impose une hypothèse supplémentaire : en effet, l'UDP galactose peut difficilement être synthétisé à partir du galactose-1-P sans apporter l'UDP comme substrat.
Une expérience supplémentaire est donc effectuée : en utilisant du 14C-UDP-glucose à concentration saturante, en absence de galactose-1-P, le 14C-UMP se fixe de façon covalente sur l'enzyme. 14C-UDP-glucose + Enzyme ----> 14C-UMP-Enzyme + glucose-1-P Cette réaction démontre la validité de la 1ère partie du schéma du haut ou la 2nde partie du schéma du bas. On suit aussi la libération de glucose-1-P. Le tableau suivant résume les résultats obtenus. |
| E (μM) | 14C-UMP incorporé sur l'enzyme (μM) | glucose-1-P (μM) |
| 6 | 6,1 | 6,2 |
| 6 | 5,9 | 5,8 |
| 2 | 1,9 | 1,8 |
| 2 | 2,1 | 2,2 |
D'après ces résultats, on obtient des quantités équimolaires pour les 2 substrats (UDP-glucose et glucose-1-P). Le schéma du haut est donc correct : l'enzyme doit être sous la forme libre E pour fixer l'UDP-glucose. En revanche, l'enzyme devrait être sous la forme E-UMP pour libérer l'UDP-galactose comme dans le schéma du bas. |