| Les effets biologiques des rayonnements ionisants |
| Tweet |
|
I. Processus physico-chimiques à l'origine des effets biologiques
1. Les interactions physiques rayonnements ionisants - matièreII. Les lésions moléculaires 1. Lésions induites par transfert direct d'énergieIII. Les dommages cellulaires 1. Mort cellulaire, cancérogénèse et effets héréditaires |
IV. Les effets pathologiques des rayonnements ionisants chez l'homme
1. Les effets déterministesV. L'évaluation du risque d'effets stochastiquesa. Leur origine et leurs caractéristiques2. Les effets stochastiques 1. Les sources d'information sur la cancérogénèsea. Les études in vitro2. L'extrapolation des risques de cancers aux faibles doses |
I. Processus physico-chimiques à l'origine des effets biologiques Dés la découverte des rayonnements ionisants en 1895, leurs dangers ont été mis en évidence:
Les effets des rayonnements ionisants sur un organisme résultent du transfert d'énergie à la matière, transfert qui constitue un processus complexe. |
| Effets des rayonnements ionisants | Organisation schématique d'un être vivant (H = homme) | |
| interactions physiques | molécules |
|
| réactions physico-chimiques | cellules (H: 1014) |
|
| >lésions moléculaires | >tissus | >ensemble de cellules différenciées et de cellules souches qui remplit une fonction |
| >dommages cellulaires | >organes | >ensemble de tissus qui assure une fonction vitale |
| >lésions tissulaires | >organisme | >ensemble d'organes |
1. Les interactions physiques rayonnements ionisants - matière Les interactions physiques entre la matière et les particules directement ionisantes ou celles mises en mouvement par les rayonnements indirectement ionisants sont de 3 types:
En moyenne pour 1 ionisation, on a 3 excitations et un nombre plus élevé de transferts thermiques. Exemple : L'énergie moyenne pour provoquer une ionisation dans l'eau est de 33,85 eV. Ainsi dans l'eau, 1 électron de 1 MeV produit environ : (106 eV / 33,85 eV) = 3 104 ionisations et 105 excitations. |
2. Les réactions physico-chimiques Il s'agit plus précisément de réactions radiochimiques résultant des interactions physiques et qui surviennent dans des délais trés brefs de l'ordre de 10-12 secondes. a. La formation de radicaux libres Un radical libre porte sur sa couche électronique externe un ou plusieurs électrons célibataires (non apparié à un électron de spin opposé). Cette configuration confère à l'entité radicalaire une trés haute réactivité chimique: les radicaux tendent à capturer un électron pour compléter leur couche électronique. Les radicaux libres proviennent essentiellement de l'interaction des rayonnements ionisants avec les électrons des molécules d'eau, et ce en raison de la teneur extrèmement élevée en eau des organismes vivants. 1er cas : L'eau est ionisée si E > 5,16 eV, qui est
l'énergie de la liaison H-OH : hν + H2O
-> e- + H2O+ 2ème cas : La molécule d'eau est excitée : hν + H2O
-> H2O* Finalement : 10-10 secondes à 10-9 secondes aprés l'interaction avec le rayonnement ionisant, l'eau est devenue une solution plus ou moins concentrée en radicaux OH° et H° et en molécules d'hydrogène issues de la réaction : H° + H° -> H2 b. Le rôle du transfert linéique d'énergie Les réactions qui succèdent à la formation de radicaux libres surviennent dans un délai de 10-7 secondes à 10-3 secondes aprés l'interaction du rayonnement ionisant et constituent l'étape diffusionnelle. Ces réactions dépendent du transfert linéique d'énergie du rayonnement. 1er cas : Le T.L.E. est élevé, donc le nombre
d'ionisation est grand, donc la probabilité de rencontre entre
les radicaux libres formés est élevée : 2ème cas : Le T.L.E. est faible et la réaction de recombinaison est la plus probable : H° + OH° -> H2O c. L'influence de la présence d'oxygène Les réactions qui succèdent à la formation de radicaux libres dépendent également de la présence de l'oxygène: 1er cas : En absence d'oxygène, la réaction
s'arrète par dimérisation : 2ème cas : En présence d'oxygène, la réaction
suivante à lieu : H° + O2 -> OOH° L'électron arraché de la molécule
d'eau ionisée lors de l'interaction avec le rayonnement ionisant
peut réagir avec l'oxygène et former le radical superoxyde,
O2°-: d. L'influence de la présence de molécules organiques Les réactions qui succèdent à la formation de radicaux libres dépendent enfin de la présence de molécules organiques. Un grand nombre de réactions (voir ci-après) ont lieu et aboutissent à la formation:
Les peroxydes et les tétroxydes sont des oxydants puissants qui altèrent les lipides des membranes des cellules. Ces réactions sont : e. L'inactivation des radicaux libres Dans toutes cellules normales il existe des réactions qui produisent des radicaux libres et des peroxydes similaires à ceux formés en cas de radioexposition. Les cellules possèdent donc des enzymes dont la fonction est d'inactiver ces composés extrèmement
toxiques: Remarque : la réaction catalysée par la superoxyde dismutase est équilibrée en terme de radical (symbole "°") puisque le transfert électronique met en jeu un atome de cuivre lié à l'enzyme qui est successivement oxydé puis réduit. |
|
1. Lésions induites par transfert direct d'énergie L'énergie du rayonnement est transférée directement à la molécule qui est ionisée ou est excitée. L'excédent d'énergie acquis par la molécule est perdu par:
Exemple: l'excès d'énergie d'une ionisation est de 12,6 eV et celui d'une excitation est de 6,6 eV. Dans les 2 cas, l'excès est suffisant pour rompre la liaison H-OH d'une molécule d'eau dont l'énergie est de 5,16 eV. 2. Lésions induites par transfert indirect d'énergie L'énergie est transférée à des molécules au voisinage des molécules concernées qui induisent à leur tour des lésions des molécules concernées. C'est typiquement l'action des radicaux libres formés lors de la radiolyse de l'eau. Actuellement, on pense que c'est l'effet direct qui prédomine. En effet, en raison de leur courte durée de vie (10-6 secondes) les radicaux libres migrent à de faibles distances. En conséquence, seuls ceux qui sont formés à proximité d'une molécule peuvent la léser. 3. Les cibles des lésions radioinduites Toutes les molécules biologiques sont des cibles. On peut mentionner en particulier: α. l'eau: en raison de sa proportion dans tous les organismes vivants (environ 70% chez l'homme) β. l'ADN: en raison de sa fonction centrale dans la cellule (altération des chromosomes et division cellulaire, altération des gènes et mutations) Le tableau suivant donne le nombre moyen de lésions de différents types induites par une dose de 1 Gy absorbée par 1 noyau : |
| Nature de la lésion de l'ADN | Nombre moyen de lésions / Gy / noyau |
| formation de liaisons intra ou inter-chaînes | 30 |
| rupture des deux brins | 40 |
| formation de liaisons ADN - protéines | 150 |
| rupture d'un seul brin | 750 |
| modification de la structure des sucres | 1200 |
| modification de la structure des bases | 1400 |
Rappel :
γ. Enfin, l'oxydation par les radicaux libres et les peroxydes des :
|
|
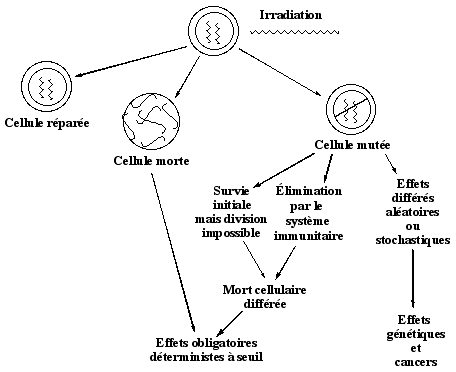
1. Mort cellulaire, cancérogénèse et effets héréditaires La mort cellulaire résulte de lésions irréparables dans des structures vitales pour la cellule telles que les chromosomes. La mort cellulaire peut être immédiate pour des doses élevées (plusieurs Gy) ou différée pour les faibles doses. Les effets des faibles doses sont les suivants :
Selon la vitesse de renouvellement du type de cellule, la mort cellulaire peut se manifester :
|
2. Les facteurs de radiosensibilité cellulaire La radiosensibilité est à la base de la compréhension des effets pathologiques des rayonnements. a. Loi de Bergonié et Tribondeau En 1906, Bergonié et Tribondeau ont montré que la radiosensibilité d'une cellule varie:
b. L'évolution Par l'étude des courbes de survie car plus un organisme est évolué plus il est radiosensible comme l'indiquent les valeurs du tableau suivant : |
| D0 (Gy) | Espèce cellulaire |
| 1000 | virus |
| 100 | levures |
| 10 | bactéries |
| 1 | mammifères |
| Remarque : Les courbes de survie étant des phènomènes exponentiels, D0 est la dose qui laisse 37% de survivants (0,37 = 1/e). | |
c. la phase du cycle cellulaire : la radiosensibilité est maximale au cours de la phase M de mitose et minimale au cours de la phase S de duplication du matériel génétique. d. l'environnement cellulaire : l'oxygène est un radiosensibilisant et ce, d'autant plus que le T.L.E. du rayonnement est faible. Ceci s'explique par l'augmentation de radicaux libres en présence d'oxygène. e. la nature du rayonnement : à dose égale, la survie cellulaire est plus importante pour des rayonnements de faible T.L.E. f. la distribution de la dose dans le temps : le fractionnement de la dose et la diminution du débit de dose permettent, à dose équivalente, d'augmenter la survie cellulaire. |
| Caractéristiques cliniques en fonction de la dose | ||
| Dose (Gy) | Dose équivalente (mSv) |
Effets |
| 0,3 - 1 | 1000 | nausées, vomissements |
| 1 - 3,5 |
3000 à 6000 |
épilation provisoire |
| syndrome hématopoïétique: atteinte des organes assurant le renouvellement des lymphocytes, des globules blancs et des plaquettes --> hémorragies | ||
| 3,5 - 5,5 | dose léthale 50: 50 % au moins des individus meurent d'hémorragies et d'infections | |
| 5 - 7 | stérilité définitive | |
| 5,5 - 7,5 | syndrome gastro-intestinal: atteinte des cellules de la mu- queuse intestinale à fort taux de renouvellement --> infections mort si pas de greffe de moelle osseuse | |
| 7,5 - 10 | > 6000 | atteinte des poumons |
| > 10 - 15 | > 10000 | syndrome neurologique: oedème cérébral et coma - mort en quelques jours |
| 20000 | mort en quelques heures | |
| c. Les effets d'une exposition partielle | |||
| Dose (Gy) | Dose équivalente (mSv) | Organe | Effets |
| réponse "tout ou rien" | embryon | avant implantation (1er - 8è jour) | |
| 0,1 | 100 | pendant l'organogénèse (9è - 60è jour) | |
| 0,1 - 0,2 | 100 - 200 | stade foetal (8è à 15è semaine) - Interruption grossesse envisagée si > 200 mSv | |
| 0,3 - 0,5 | 150 | testicules | stérilité provisoire |
| 1 | peau | fragilisation si exposition ultérieure | |
| 3 | ovaires | stérilité provisoire | |
| 5 | 3500 - 6000 | testicules | stérilité |
| 7 | 2500 - 6000 | ovaires | définitive |
| 5 - 10 | 5000 | peau | épidermite exsudative |
| cristallin | cataracte | ||
| 15 -20 | peau | ulcération et nécrose aiguë - épilation définitive | |
2. Les effets stochastiques a. Leur origine et leurs caractéristiques Les effets stochastiques sont les conséquences probabilistes à long terme, chez un individu ou chez sa descendance, de la transformation d'une cellule. Ils résultent donc de lésions mal réparées des molécules d'ADN. Les effets stochastiques sont de 2 types :
Les effets stochastiques n'ont pas de seuil de dose. Ce sont des effets aléatoires qui n'apparaissent pas chez tous les individus. Par ailleurs, les effets stochastiques sont :
|
| b. Tableau comparatif des effets déterministes et stochastiques | |
| Effets déterministes | Effets stochastiques |
| cause: destruction massive des cellules | cause : lésions non réparées de l'ADN |
| obligatoires (ils apparaissent toujours) | aléatoires |
| pathologies diverses | cancers et effets génétiques |
| dose seuil d'apparition: 0,2 - 0,3 Gy | pas de dose seuil d'apparition |
| manifestation précoce | manifestation tardive |
| gravité dépendante de la dose | gravité indépendante de la dose |
| clairements décrits | non spécifiques |
1. Les sources d'information sur la cancérogénèse a. Les études in vitro Elles ont montré :
b. L'expérimentation animale Elle a permis :
Cependant :
c. Les études épidémiologiques Le suivi des populations exposées est la seule base de données valable pour définir le facteur de risque. Ces études sont difficiles car :
Le tableau suivant résume les caractéristiques de ces modes d'étude : |
| Mode d'étude | Points favorables | Points défavorables |
| Etudes in vitro | mise en évidence de divers paramètres (cassure double-brin, débit de dose, oncogènes) |
résultats difficilement extrapolables à l'être humain |
Expérimentation animale |
- données quantitatives |
- grand nombre d'animaux
à étudier |
| Études épidémiologiques | seule base de données valable pour définir le facteur de risque |
- plus la dose est faible,
plus l'effectif de la population à étudier est grand |
2. L'extrapolation des risques de cancers aux faibles doses Ainsi, 3 populations exposées ont été choisies par l'U.N.S.C.E.A.R. ("United Nations Scientific Committee on the Effects of Atomic Radiations") : |
| Paramètres | Survivants Hiroshima Nagasaki | Patients traités pour la spondylarthrite | Patientes traitées pour le cancer du col de l'utérus |
| Taille de la population | 4801 | 14000 | 83000 |
| Age à l'exposition | 0 - 90 ans | > 15 ans | 30 - 70 ans |
| Temps moyen de suivi | 28,8 ans | 13 ans | 7,6 ans |
| Type d'exposition | instantanée corps entier | fractionnée - localisée | étalée - localisée |
| Dose (Gy) | > 0,5 | 0,8 - 6 | variable |
| Excès de décès: | |||
| par leucémies | 54 | 37 | < 100 |
| par cancers | 147 | 140 | 257 |
La figure ci-dessous montre la courbe dose-effet établie à partir des cancers observés chez les survivants d'Hiroshima et Nagasaki.
Ce résultat montre une augmentation certaine du risque cancérogène pour des expositions à des doses supérieures à 0,5 Gy, délivrées à fort débit. La forme de la courbe dose-effet suit une relation polynômiale: effet = k1 . dose + k2 . (dose)2 En revanche, l'ensemble de ces études ne permet pas de conclure dans le domaine des faibles doses inférieures à 0,2 Gy. Il faut donc extrapoler aux faibles doses, les données obtenues à fortes doses. Cependant cette extrapolation pose un problème car, pour les faibles doses, les données sont rares et l'augmentation du nombre de cancers est non significatif. L'extrapolation de la courbe des fortes doses peut donc être faite selon plusieurs types de relations compatibles avec les observations:
C'est la relation linéaire qui a été retenue. Les conséquences sont :
Ce choix amène 2 commentaires :
|
3. L'extrapolation des risques de cancers aux faibles débits de dose Les expositions sont en général du type faible débit de dose. Or celle d'Hiroshima - Nagasaki est du type fort débit de dose. L'influence du débit de dose sur la cancérogénèse est connue: pour les rayonnements de faible T.L.E., plus la dose est étalée dans le temps, plus les effets cancérogènes sont réduits. Pour tenir compte du débit de dose, on utilise un facteur appelé facteur de réduction de dose (DDREF, "Dose and Dose Rate Effectiveness Factor"), calculé à partir des courbes dose-effet. Les résultats montrent que les valeurs du facteur de réduction de dose à appliquer pour estimer l'effet délivré à faible débit de dose s'échelonnent entre 2 et 10. Le facteur le plus faible de 2 a été retenu. Remarque : Cette valeur (qui est la plus faible) majore le risque (puisque la réduction du risque est minorée) et ce, une fois encore, en réponse au principe fondamental de précaution en radioprotection. |
4. L'extrapolation des risques de cancers sur l'ensemble de la vie L'enquête épidémiologique d'Hiroshima - Nagasaki n'est pas terminée. En d'autres termes, l'estimation du facteur de rique qui a été faite jusqu'à présent n'est applicable que pour la durée de l'enquête, qui ne représente qu'une fraction de la vie des individus de la population observée. Il faut donc évaluer le risque d'apparition de cancers dans les années à venir. Pour celà, il existe 2 modèles de projection:
Dans ce cas, puisque le nombre de cancers naturels augmente avec l'âge, on doit observer une augmentation constante du nombre de cancers radioinduits. C'est le modèle du risque relatif qui a été retenu. |
5. L'extrapolation des effets héréditaires Les effets héréditaires résultent des lésions des chromosomes de la lignée germinale (spermatozoïdes et ovules) susceptibles d'entraîner des anomalies dans la descendance de l'individu exposé. Bien que les rayonnements ionisants soient l'un des agents mutagènes les mieux connus, leur action chez l'homme n'a pas été formellement mise en évidence. En effet, stérilité et décès interviennent avant qu'une quelconque descendance n'ait manifesté des effets héréditaires. Les sources d'information et les difficultés des enquêtes sont les mêmes que dans le cas de l'études des effets cancérogènes. L'enquête la plus importante a de nouveau porté sur les survivants d'Hiroshima et Nagasaki et n'a révélé aucune augmentation significative de la fréquence des anomalies génétiques. De plus, l'incidence naturelle des effets héréditaire est élevée: environ 10% des naissances. En conséquence, puisqu'il n'existe pas de données chez l'homme, l'évaluation du risque d'effets héréditaires repose sur les résultats obtenus chez la souris. Dans un premier temps, en ce qui concerne la souris proprement dit, les méthodes et les choix d'extrapolation aux faibles doses des données obtenues aux fortes doses sont les mêmes que pour l'étude chez l'homme des risques de cancers. Enfin, l'extrapolation des données obtenues chez la souris à l'homme repose sur les 60% d'homologie génétique entre l'homme et la souris. En d'autres termes, si 2 gènes sont liés chez la souris, ils le sont également chez l'homme dans 60% des cas. |
![]()