| Les virus et l'infection par les virus |
| Tweet |
|
|
1. Introduction 2. Classification et taxonomie des virus 3. Entrée des virus enveloppés dans la cellule hôte a. L'enveloppe 4. Entrée des virus non enveloppés dans la cellule hôte
5. Intégration du génome viral 6. Réplication et transcription du génome viral, traduction des protéines virales
7. La capside
|
8. Assemblage de la capside et empaquetage du génome
9. Sortie du génome viral du noyau de la cellule hôte
10. Bourgeonnement et sortie de la cellule hôte avec les machineries ESCRT 11. La maturation des virions
12. Lysogénie et lyse 13. Arbitrage entre lysogènie (voie latente) et voie lytique 14. Les protéines du virus Ebola 15. Médicaments antiviraux et développement de nouveaux antiviraux 16. Mise au point de vaccins contre le coronavirus SARS-CoV-2 17. Nanomatériaux biomimétiques : thérapies antivirales et développement de vaccins 18. Apport des méthodes d'apprentissage profond 19. Liens Internet et références bibliographiques |
|
Ce cours est un aperçu de l'ensemble des connaissances en virologie. En effet, détailler les spécificités de toutes les étapes du cycle de vie de chaque classe de virus, et ce, pour tous les types de cellules infectées (animaux (mammifères, homme, ...), plantes, bactéries Gram+ et Gram-, ...) est l'objet d'ouvrages spécialisés. Malgré tout, en focalisant sur les principes des interactions de certains couples [virus - cellule hôte], ce cours décrit les caractéristiques fondamentales des virus et de l'infection virale. |
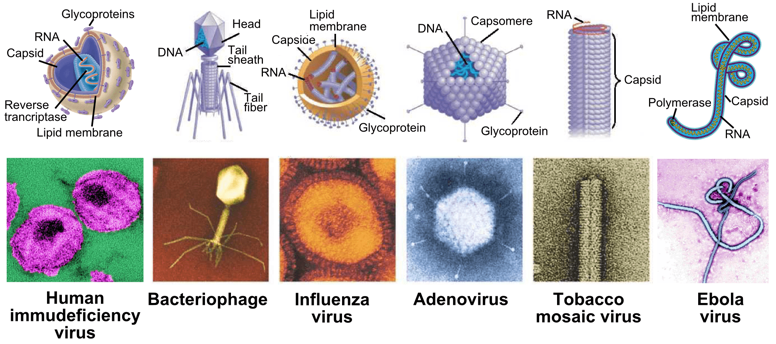
1. Introduction Les virus sont des parasites de tous les types de vie cellulaire car ils ne peuvent pas se reproduire. Ils infectent une cellule hôte et utilise sa machinerie pour produire d'autres particules de virus. La particule virale infectieuse, appelée virion, est constituée d'un acide nucléique (ADN ou ARN) entouré d'une enveloppe protéique, la capside, constituée de nombreux exemplaires d'un ou plusieurs types de protéines.
Source : Callaway E. (2017) - Animated Healthcare Ltd. Les virus représentent la forme de vie la plus abondante et la plus diverse sur Terre. Ils peuvent infecter tous types d'organismes (vertébrés et invertébrés, plantes, champignons, bactéries, archées). Les virus qui infectent les bactéries sont appelés bactériophages ou phages : ils représentent la forme biologique la plus abondante dans la biosphère avec environ 4,8 1031 phages (10 fois plus nombreux que les bactéries) et une biomasse estimée à 200 millions de tonnes.
Il est admis que les virus sont polyphylétiques et différents scénarios quant à leur origine sont envisagés :
L'étude des virus de plantes est à l'origine des premières expériences de biologie moléculaire. En 1935, Wendell Stanley (co-lauréat avec John Howard Northrop du Prix Nobel de chimie en 1946 "for their preparation of enzymes and virus proteins in a pure form") a purifié et partiellement cristallisé le virus de la mosaïque du tabac (dont il attaque les feuilles). |
Quelques virus "célèbres" Exemples d'infections virales médiatisées : virus du chikungunya (moustique "tigre" Aedes albopictus ou Aedes aegypti), virus de la dengue (moustiques du genre Aedes), virus HIV-1 (sida), virus Zika (Flavivirus, moustiques du genre Aedes). Les virus de la grippe Il existe 4 types de virus de la grippe : A, B, C et D. Le type A est celui dont l'infection est la plus virulente chez l'homme.
Sources : Centers for Disease Control / Biopharma L'ARN du virus Zika Des structures peu communes en forme de nœud de certains ARN viraux résistants aux exoribonucléases ("viral exoribonuclease-resistant RNAs" - xrRNA) empêchent leur hydrolyse par les ribonucléases de l'hôte, augmentant ainsi le pouvoir infectieux et la pathogénicité de ces ARN viraux. Ces xrRNA empêchent l'hydrolyse du fait d'une résistance mécanique à leur dépliement. Le dépliement d'un xrRNA du virus Zika avec des "pincettes optiques" montre qu'il s'agit de l'ARN le plus stable sur le plan mécanique observé à ce jour :
|
Les coronavirus Les coronavirus (Nidovirales, Coronaviridae, Coronavirinae) Ces virus tirent leur nom de leur halo en forme de couronne. Ce sont des virus enveloppés et sphériques de 100 à 160 nm de diamètre, contenant un génome à ARN simple brin positif de 27 à 32 kilo paire de bases.
Figure ci-dessous : particules virales libérées de la surface d'une cellule infectée par le coronavirus SARS-CoV-2.
Source : Nicholls et al., Université de Hong Kong
Autres exemples de coronavirus
|
Quelques définitions
Source : Clokie M.R.J. (2018)
Source : ViralZone
|
Les virus n'ont pas de structure cellulaire et ne peuvent pas effectuer de réactions métaboliques. En revanche, ils utilisent la machinerie métabolique et génétique de la cellule hôte qu'ils infectent pour se multiplier. On peut les considérer comme des "parasites génétiques" et non comme des êtres vivants indépendants. Les caractéristiques structurales et fonctionnelles de leurs gènes sont semblables à celles des gènes de leurs cellules hôtes. Contrairement aux génomes des autres êtres vivants, composés exclusivement d'ADN, certains génomes viraux sont constitués d'ARN. Certains virus sont dits à ARN négatif car leur matériel génétique est complémentaire de celui d'un ARN messager (qualifié de positif) et donc "illisible" par la cellule hôte. Leur génome contient un nombre variable de gènes. |
| Exemples de quelques virus | ||||
| Virus | Forme | Génome | Nucléotides | Gènes |
| Parovirus (virus des Vertébrés) | 1 molécule linéaire | ADN simple brin | 2 000 - 5 000 | 3 - 5 |
| Virus ΦX 174 (1er génome complet d'un virus séquencé - 1978) |
----- | ADN simple brin | 5 386 | 11 |
| Virus de la mosaïque du tabac | ----- | ARN simple brin positif | 6 395 | 4 |
| Virus de la grippe (virus des Vertébrés) | 8 molécules linéaires | ARN simple brin négatif | 13 500 | 12 |
| Bactériophage T4 (virus des Bactéries) | 1 molécule linéaire | ADN double brin | 165 000 | 200 |
| Virus de la vaccine (virus des Vertébrés) diamètre 300 - 400 nm |
----- | ADN double brin | 240 000 | > 200 |
| Voir une liste plus complète. | ||||
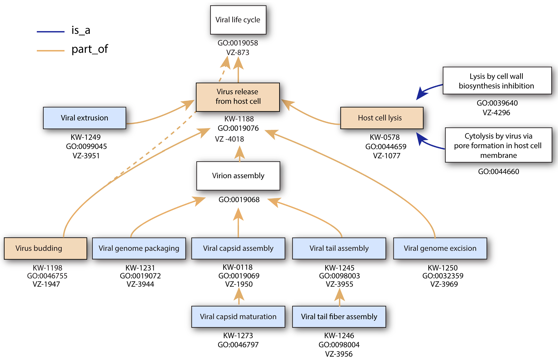
Exemple d'ontologie (termes et numéros d'accession GO, "Gene Ontology") pour l'annotation des évènements clés du cycle de vie d'un virus (VZ : page de la base de données ViralZone; KW : mot clé UniProtKB).
Source : Hulo et al. (2017) |
2. Classification et taxonomie des virus
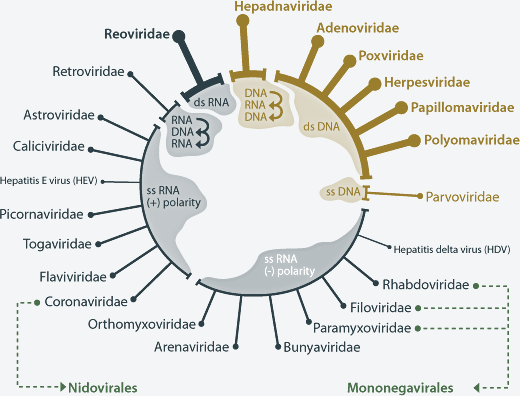
Classification selon la méthode de David Baltimore sur la structure du génome (voir "Family Groups - The Baltimore Method") :
Source : ViralZone David Baltimore a été co-lauréat du Prix Nobel de physiologie ou médecine en 1975 pour la découverte de la transcriptase inverse (enzyme des rétrovirus), conjointement à Renato Dulbecco et Howard Martin Temin "pour leurs découvertes concernant l'interaction entre les virus tumoraux et le matériel génétique de la cellule". Voir la classification taxonomique au NCBI.
Source : Antiviral InteliStrat Inc. |
ICTV : International Committee on Taxonomy of Viruses L'ICTV (anciennement "International Committee on Nomenclature of Viruses") est une organisation mondiale à but non lucratif. C'est un comité de la division de virologie de l'Union internationale des sociétés de microbiologie ("Virology Division of the International Union of Microbiological Societies" - IUMS). L'ICTV est responsable de l'élaboration de la taxonomie (y compris la classification officielle) de tous les virus, viroïdes et satellites, ainsi que de la nomenclature des taxons approuvés. L'ICTV gère également plusieurs ressources Web au service de la communauté de virologie :
|
Classification de quelques familles de virus selon leur taille
Les virus dits géants (découverts très récemment) ou virus à grand ADN nucléo-cytoplasmique ("NucleoCytoplasmic Large DNA Viruses", NCLDV) constituent l'un des plus grands groupes de virus qui infectent une grande variété d'hôtes eucaryotes. Exceptés les Poxvirus, les Pandoravirus et les Ascovirus, ils ont une forme approximativement icosaédrique. Voir "The largest virus genomes and sequences" - PIT Bioinformatics Group, Eotvos Lorand University, Budapest. Les connaissances sur les NCLDV sont principalement dérivées d'isolats viraux co-cultivés avec des protistes et des algues. Cependant, 2074 génomes de NCLDV ont été reconstruits à partir d'échantillons prélevés à travers le monde et les données des métagénomes (Schulz et al., 2020).
La diversité des NCLDV (qui sont par ailleurs associés aux principales lignées des eucaryotes) est en faveur de l'hypothèse qu'ils sont des acteurs importants des écosystèmes à travers les biomes. |
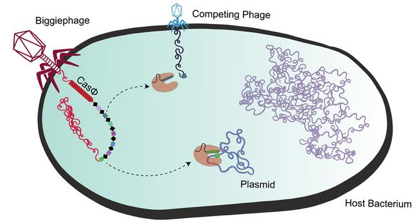
Les phages à très grand génome et leurs systèmes CRISPR-Cas Les bactériophages ont généralement de petits génomes. Cependant, il existe des centaines de génomes de phages de plus de 200.000 paires de bases, jusqu'à 735.000 paires de bases. Dix clades de phages ont été identifiés et nommés : Mahaphage, Kabirphage, Dakhmphage, Jabbarphage, Kyodaiphage, Biggiephage, Whopperphage, Judaphage, Enormephage et Kaempephage (Al-Shayeb et al., 2020). Les répertoires génétiques de ces phages à très grand génome contiennent divers systèmes CRISPR-Cas, des ARN de transfert (ARNt), des ARNt synthétases, des enzymes de modification de l'ARNt, des facteurs d'initiation et d'élongation de la traduction et des protéines ribosomales. Les systèmes CRISPR-Cas de ces phages ont la capacité de réduire au silence les facteurs de transcription de la bactérie hôte et donc les gènes qui devraient être transcrits. Les Biggiephages possèdent un système CRISPR-Cas très particulier :
Source : Pausch et al. (2020) |
3. Entrée des virus enveloppés dans la cellule hôte La reconnaissance et la fixation sur la cellule hôte est le préalable à toute infection par un virus. La membrane des cellules est caractérisée par de nombreux types de macromolécules (protéines, glycérophospholipides, glycanes, ...) dont le nombre, la structure et la composition sont la signature des cellules. En conséquence, les nombreux virus ciblent ces macromolécules (qu'on appelle récepteurs) pour entrer dans la cellule hôte. Certains virus (exemples : virus HIV, de l'herpès, ...) sont dits enveloppés ("enveloped virus") : en effet, ils possèdent une membrane, appelée enveloppe, qui provient de la cellule infectée : au cours du processus de bourgeonnement ("budding off"), les particules virales nouvellement formées ont une enveloppe qui, dès lors, entoure leur nucléocapside protéique. L'enveloppe peut aider un virus à survivre, à contourner le système immunitaire défensif de l'hôte, à infecter d'autres cellules.
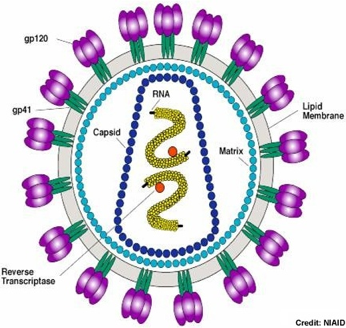
Source : Henderson C. (NIAID - NIH) - gp41 et gp120 sont des glycoprotéines d'enveloppe L'enveloppe virale est constituée par des éléments provenant de la membrane plasmique (bicouches lipidiques et protéines) de la cellule hôte ou du virus. Les virus remplacent cependant les protéines membranaires d'origine par leurs propres protéines, en particulier des glycoprotéines qui "pointent" et leur permettent de reconnaitre leurs cellules cibles et de se fixer à leur surface.
|
|
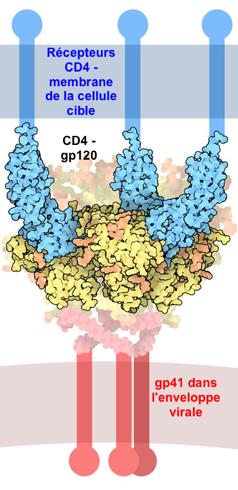
Illustration : la glycoprotéine d'enveloppe Env du rétrovirus (lentivirus) HIV-1 Cette glycoprotéine est synthétisée sous forme d'un précurseur appelé gp160 (PDB 4CC8) très flexible et recouvert de glucides. Ce précurseur est hydrolysé :
Les sous-unités gp120 et gp41 s'associent de manière non covalente pour former un trimère d'hétérodimère [gp120-gp41]3. Du point de vue ultra-structural, elles forment des pointes dans l'enveloppe du virion. Le trimère [gp120-gp41]3 du HIV-1 interagit avec le récepteur primaire CD4 et un corécepteur (tel que le récepteur de la chimiokine CCR5, un RCPG) pour fusionner les membranes du virus et de la cellule cible. L'interaction entre gp120 et le corécepteur semble l'élément déclencheur de l'acquisition du potentiel de fusion de gp41. Le récepteur CD4 ("Cluster of Differentiation 4") est une glycoprotéine présente à la surface de cellules immunitaires (cellules T auxiliaires, monocytes, macrophages, cellules dendritiques). |
Figure adaptée de "HIV Envelope Glycoprotein - PDB" |
b. Incorporation des glycoprotéines dans l'enveloppe virale
La synthèse de protéines virales dans la cellule hôte peut provoquer un stress dans le réticulum endoplasmique : les cellules hôtes disposent d'un mécanisme (appelé "Unfolded Protein Response" - UPR) pour répondre au stress lié à l'accumulation de protéines mal repliées dans le réticulum endoplasmique et ainsi l'atténuer. Une particularité importante de l'UPR est sa capacité à augmenter la voie de dégradation des protéines associée au réticulum endoplasmique ("ER-Associated protein Degradation pathway" - ERAD), notamment via son mécanisme de contrôle de la qualité du repliement des protéines. C'est donc une "lutte évolutive" incessante entre les virus et la machinerie ERAD de leurs hôtes. Modèles (non mutuellement exclusifs) d'incorporation des glycoprotéines Env dans l'enveloppe virale de nouveaux virions
Source : Freed E.O. (2015) - CA : capside / NC : nucléocapside
|
c. Protéines de fusion virales et fusion [enveloppe du virus - membrane de l'hôte] Les glycoprotéines assurent le processus dit de fusion entre l'enveloppe virale et les membranes des cellules hôtes. Les glycoprotéines de fusion virales sont en conséquence de plus en plus des cibles de médicaments antiviraux (voir Vigant et al., 2015).
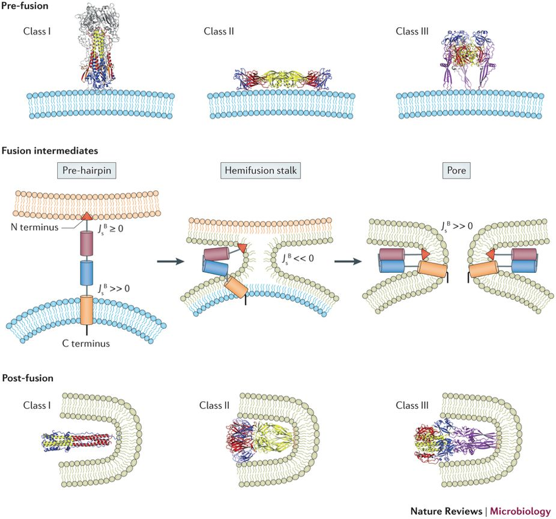
Source : Xiong Laboratory - Yale University Trois classes de glycoprotéines de fusion virales sont pour l'instant identifiées sur la base des similitudes de structures. |
| Classe | I | II | III* |
| Exemples de virus et de glycoprotéine de fusion (lien vers Uniprot) |
HIV-1, Retroviridae (Envelope glycoprotein gp160) Paramyxovirus, Paramyxoviridae (Fusion glycoprotein F0 precursor) |
Chikungunya, Alphavirus (E1 structural glycoprotein) Zika, Flaviviridae (Envelope glycoprotein) |
VSV (Vesicular stomatitis virus), Rhabdoviridae (Glycoprotein G) HSV-1 (Herpes simplex virus 1), Herpesviridae (Envelope glycoprotein B) |
| Structure quaternaire de la protéine de fusion native | trimère hélices α |
dimère feuillets β |
trimère hélices α et feuillets β |
| Orientation par rapport à la membrane virale | perpendiculaire - projections sous forme de pointes | parallèle | perpendiculaire |
| Protéolyse pour générer une forme de fusion compétente | oui (de la protéine de fusion) | oui (d'une protéine accessoire) | non |
| Localisation du peptide de fusion dans la séquence primaire | à l'extrémité N-terminale ou proche | boucle simple au sein de la séquence | boucle bipartite au sein de la séquence |
| Structure quaternaire après la fusion | trimère d'épingles à cheveux : spirale centrale d'hélices α | trimère d'épingles à cheveux : principalement feuillets β | trimère d'épingles à cheveux : spirale centrale d'hélices α et feuillets β |
| Structure 3D (PDB) | 2HMG - 1QU1 | 1SVB - 1URZ | 2J6J - 2CMZ |
|
|||
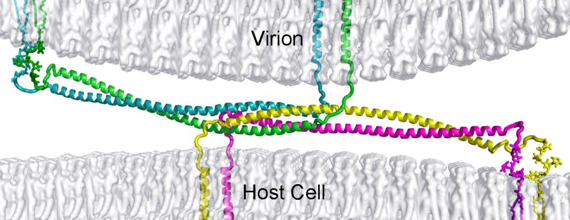
Mécanisme de fusion [enveloppe du virus - membrane de l'hôte] C'est la mise en contact intime de 2 bicouches membranaires distinctes suivie de leur fusion en une seule. Les peptides de fusion sont les séquences hydrophobes qui interagissent directement avec la membrane cible. Les étapes (quasiment identiques pour les 3 classes de protéines de fusion) sont les suivantes :
Source : Vigant et al. (2015) Les protéines adoptent des structures quaternaires très particulières :
|
|
Induit par un signal associé à l'entrée dans la cellule à infecter (exemples : liaison au [récepteur/co-récepteur], fixation de protons dans un endosome), les glycoprotéines de fusion subissent plusieurs changements de conformation. Un segment hydrophobe de la chaîne polypeptidique, appelé peptide de fusion (ou boucle de fusion) interagit avec la membrane de la cellule cible et cet intermédiaire de pontage rapproche les deux membranes (virus et cellule). Les peptides de fusion sont localisés de 3 manières dans la séquence polypeptidique :
|
| Caractéristiques de quelques peptides de fusion viraux de protéines de fusion de la classe I | |||||||
| Famille | Espècea | Séquence d'acides aminés | Position | AAAb | ΔG Interfacec | % Alad | % Glyd |
| Retroviridae | HTLV-1 | AVPVAVWLVSALAMGAGVAGGITGS | N-ter | oui | - 1,24 | 24/9 | 20/8 |
| HIV-1 | AVGIGALFLGFLGAAGSTMGARS | N-ter | oui | - 2,29 | 22/7 | 26/7 | |
| ASLV | GPTARIFASILAPGVAAAQALREIERLA | interne | oui | 5,94 | 29/8 | 7/7 | |
| SIV | GVFVLGFLGFLATAGSAMGAAS | N-ter | oui | - 3,86 | 23/7 | 23/7 | |
| BLV | SPVAALTLGLALSVGLTGINVAVS | N-ter | non | - 0,58 | 17/7 | 13/3 | |
| Filoviridae | EBOV | GAAIGLAWIPYFGPAAE | interne | oui | - 1,3 | 29/5 | 18/6 |
| MARV | LAAGLSWIPFFGPGI | interne | oui | - 4,45 | 13/5 | 20/7 | |
| Coronaviridae | SARS-CoV | MYKTPTLKYFGGFNFSQIL | N-ter | oui | - 3,07 | 0/8 | 11/7 |
| GAALQIPFAMQMAYRF | interne | oui | - 1,42 | 25/8 | 6/7 | ||
| Orthomyxoviridae | IFV-A | GLFGAIAGFIENGWEGMIDGWYG | N-ter | oui | - 2,52 | 9/8 | 30/6 |
| PIV5 | FAGVVIGLAALGVATAAQVTAAVALV | N-ter | oui | 0,04 | 35/11 | 13/4 | |
| Paramyxoviridae | NDV | FIGAIIGSVALGVATAAQITAA | N-ter | oui | - 0,45 | 30/8 | 13/8 |
| HeV | LAGVVMAGIAIGIATAAQITAGV | N-ter | non | 0,27 | 30/6 | 17/6 | |
Source : Apellaniz et al. (2014) |
|||||||
| Caractéristiques de quelques peptides de fusion viraux de protéines de fusion de la classe II | |||||||
| Famille | Espècea | Séquence d'acides aminés | Position | Etat avant fusion | ΔG Interface | % Ala | % Gly |
| Flaviviridae | TBEV | DRGWHNGCGLFGKGSI | interne | homodimère | 0,5 | 0/8 | 31/9 |
| DENV-1 | DRGWGNGCGLFGKGSL | interne | homodimère | - 0,7 | 0/7 | 38/10 | |
| Togaviridae | SFV | VYTGVYPFMWGGAYCFCDS | interne | hétérodimère | - 3,86 | 4/8 | 13/7 |
| CHIKV | VYPFMWGGAYCFCDTENT | interne | hétérodimère | - 2,11 | 6/11 | 12/6 | |
|
aTEBV: Tick-Borne Encephalitis Virus, DENV-1: Dengue Virus type 1, SFV: Semliki Forest Virus, CHIKV: Chikungunya Virus |
|||||||
| Caractéristiques de quelques peptides de fusion viraux de protéines de fusion de la classe III | |||||||
| Famille | Espècea | Séquence d'acides aminés | Position | Etat avant fusion | ΔG Interface | % Ala | % Gly |
| Rhabdoviridae | VSV | WY/YA | bipartite | trimère | - 3,56 | 25/5 | 0/7 |
| Herpesviridae | HSV | VWFGHRY/RVEAFHRY | bipartite | trimère | 0,7 | 7/11 | 7/7 |
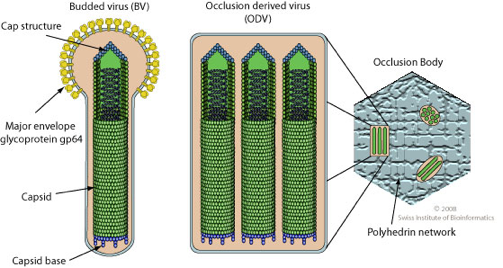
| Baculoviridae | AcMNPV | YAYNGGSLDPNTRV/VKRQNNNHFAHHTCNK | bipartite | trimère | 8,36 | 7/5 | 7/5 |
|
aVSV: Vesicular Stomatitis Virus, HSV: Herpes Simplex Virus and AcMNPV: Autographa californica Multiple Nucleopolyhedrovirus |
|||||||
e. Incidence du pH des endosomes sur la fusion des membranes Un pH bas est un élément déclencheur de la fusion pour certains virus (orthomyxovirus, rhabdovirus, alphavirus, flavivirus, bunyavirus et arénavirus). Ces virus pénètrent dans les cellules par endocytose et fusionnent avec des endosomes précoces (exemple, le virus VFS) ou tardifs (exemple, le virus de la grippe). Un pH faible induit des réarrangements de la structure des 3 classes de protéines de fusion qui permettent de positionner le peptide de fusion afin qu'il puisse s'enfouir dans la membrane cible. Pour les protéines de fusion de la classe I, cela implique la séparation des domaines de la tête globulaire qui fixent la sous-unité de fusion dans son état de pré-fusion. Par exemple, la glycoprotéine G attache le virus VSV aux récepteurs LDL de l'hôte, induisant une endocytose du virion dépendante de la clathrine. Dans l'endosome, le pH acide induit des modifications conformationnelles du trimère de la glycoprotéine qui déclenchent la fusion entre le virus et la membrane endosomale. Illustration du virus de la grippe Dans le cas du virus de la grippe, la glycoprotéine de liaison au récepteur de la cellule cible est l'hémagglutinine (HA) :
HA est une protéine trimérique et chaque sous-unité est constituée de 2 domaines (HA1 et HA2 ) reliées par des ponts disulfures :
Le faible pH des endosomes active la fusion en facilitant les changements conformationnels irréversibles de la glycoprotéine :
|
|
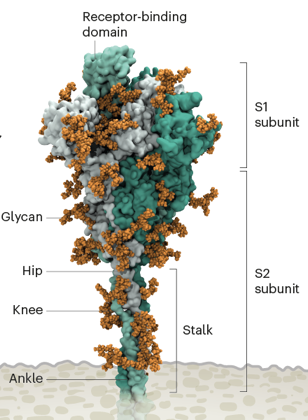
La glycoprotéine de pointe et le domaine de fixation à la cellule hôte Un coronavirus pénètre dans la cellule hôte via sa glycoprotéine de pointe : celle-ci est synthétisée sous forme d'un précurseur inactif qui doit être protéolysé pour médier la fusion des membranes du virus et de l'hôte. Chaque virion du SARS-CoV-2 possède 24 à 40 glycoprotéines de pointe (ou protéine d'enveloppe S - "spike glycoprotein") distribuées au hasard à sa surface :
La glycoprotéine de pointe est synthétisée sous forme d'un précurseur inactif protéolysé en :
Trois hétérodimères [S1/S2] s'assemblent pour former la glycoprotéine de pointe qui sort de l'enveloppe virale.
Source : Scudellari M. (2021) Un domaine particulier situé dans la sous-unité S1 de la glycoprotéine de pointe du SARS-CoV-2 est appelé "Receptor Binding Domain" (RBD) : il se fixe au récepteur ACE2 situé en surface de la plupart des cellules hôtes de la gorge et des poumons de l'homme.
Voir une remarquable vidéo : "How does SARS-CoV-2 enter and replicate in cells ?" ("Comment le SARS-CoV-2 pénètre-t-il et se réplique-t-il dans les cellules ?") |
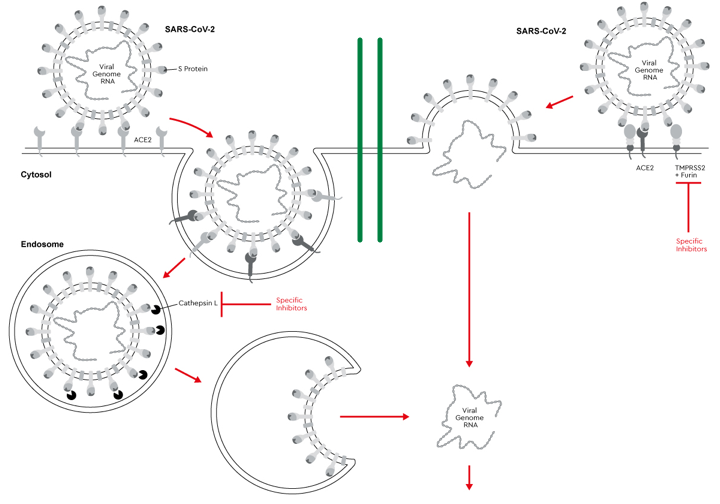
Entrée du SARS-CoV-2 dans la cellule hôte La glycoprotéine de pointe S est une protéine de fusion de la classe I trimèrique. Elle doit être activée par protéolyse pour médier la fusion des membranes du virus et de la cellule hôte. Selon la séquence en acides aminés à la jonction entre les sous-unités S1 et S2, cette protéolyse est effectuée :
Source : Promocell Exemple de la protéase à sérine transmembranaire TMPRSS2 ("TransMembrane PRoteaSe Serine 2" - E.C. 3.4.21.B60) présente en grand nombre à la surface des cellules hôtes respiratoires.
|
Les variants du SARS-CoV-2 Le récepteur ACE2 est également le point d'ancrage du SARS-CoV : cependant le SARS-CoV-2 se lie à l'ACE2 deux à quatre fois plus fortement que ne se lie le SARS-CoV. La protéine de pointe du SARS-CoV et celle du SARS-CoV-2 ont 76,5% d'identité d'acides aminés et une forte homologie.
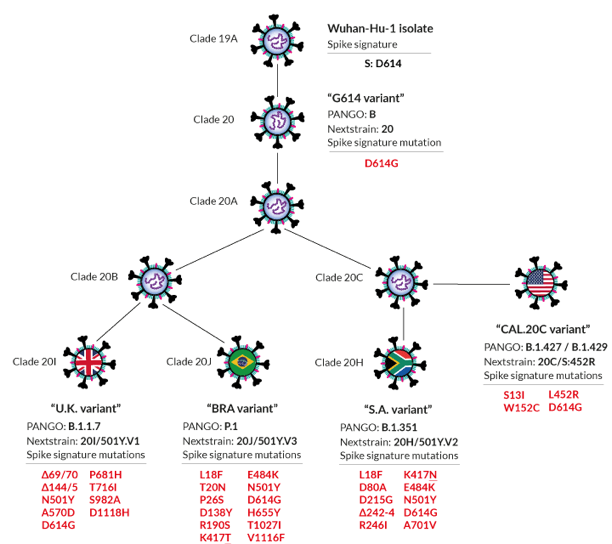
L'isolat Wuhan-Hu D614 a été la première souche du SARS-CoV-2 décrite au début de la pandémie (décembre 2019) et il appartient au clade 19A.
Source : InvivoGen |
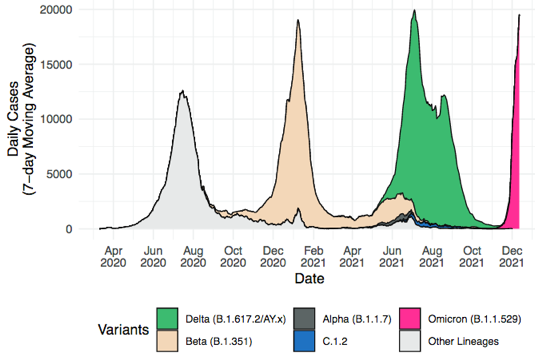
Description de quelques variants du SARS-CoV-2
Source : Viana et al. (2022) Voir des explications complémentaires de la nomenclature des variants. Des infections par le SARS vieilles de plusieurs décennies déclenchent une réponse puissante aux vaccins contre la COVID. La production spectaculaire d'anticorps chez les personnes infectées lors de l'épidémie de 2002 - 2004 renforce les espoirs d'un vaccin contre de nombreux coronavirus (Moyo-Gwete et al., 2021). |
4. Entrée des virus enveloppés dans la cellule hôte Les cellules de mammifères sont couvertes de protéines et de lipides glycosylés et les acides sialiques (exceptés les glycosaminoglycanes) sont les unités les plus externes des constituants glycosylés de la membrane plasmique. Les acides sialiques sont donc les récepteurs de plusieurs classes de virus (les rôles spécifiques de l'acide sialique et des autres récepteurs du virus dépendent du virus et du type de cellule). Les deux formes les plus courantes d'acide sialique reconnues par les virus qui ciblent des cellules de l'homme et de mammifères sont l'acide 5-N-acétylneuraminique (Neu5Ac) et l'acide 5-N-glycolylneuraminique (Neu5Gc). De manière générale, le terme acide sialique désigne les dérivés de l'acide neuraminique.
Source : Neu et al. (2008) La plupart des virus non enveloppés ne fusionnent pas avec la membrane plasmique car ils ne possèdent pas de revêtement sous forme d'une bicouche lipidique (à l'inverse des virus enveloppés) : ces virus transitent par des vésicules d'endosomes, par l'appareil de Golgi ou le réticulum endoplasmique. Ainsi, après s'être liés aux récepteurs à la surface de la cellule cible, les virus non enveloppés pénètrent dans les cellules hôtes par macropinocytose, par endocytose médiée par la clathrine, par endocytose médiée par cavéole (invagination) ou par une combinaison de ces modes d'entrée. |
| virus | récepteur à la surface de la cellule ciblée | rôle | Mode d'entrée |
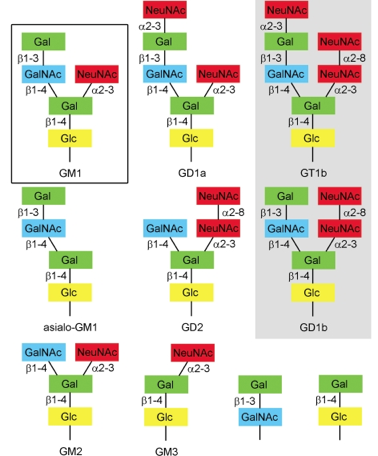
| virus simien 40 (SV40) | ganglioside GM1 | faire entrer le virus | endocytose cavéoline-dépendente et indépendente, cholestérol-dépendente |
| polyomavirus de souris (MPyV) | gangliosides GD1a et GT1b | faire entrer le virus | pinocytose Cav-1-dépendente |
| virus BK (BKPyV) | gangliosides GD1b et GT1b | faire entrer le virus | endocytose Cav-1-dépendente et indépendente, clathrine-indépendente, cholestérol-dépendente |
| adénovirus humain (Ad) | GD1a lié à une glycoprotéine (liaison O-); acide polysialique | fixer le virus | endocytose médiée par la clathrine |
| réovirus souris | ganglioside GM2; glycanes dérivés par l'acide sialique | renforcer l'adhésion du virus | endocytose clathrine-dépendente |
| SARS-CoV-2 | gangliosides monosialilés (glycanes dérivés par l'acide sialique, en particulier les glycolipides) | faciliter l'entrée du virus | Voir Nguyen et al. (2022) |
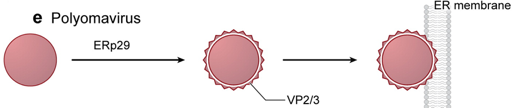
a. Modification subséquente de la membrane par les protéines de la capside des virus non enveloppés Après leur internalisation, les virus non enveloppés (exemples : Adenoviridae, Papillomaviridae, Polyomaviridae, Parvoviridae, ...) doivent [rompre / solubiliser / dégrader] :
La libération dans le cytosol de la nucléocapside de ce type de virus est généralement le fruit de peptides (de 10 à 60 résidus d'acides aminés*) de caractère global hydrophobe ou amphipathique.
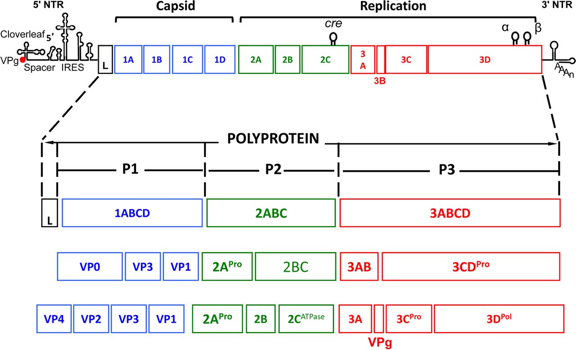
Illustration : structure du génome des picornavirus et maturation des polyprotéines par protéolyse Le génome des picornavirus est un ARN simple brin de 7100 à 8900 paires de bases.
Source : Jiang et al. (2014) La longue région 5' non traduite contient un site d'entrée interne des ribosomes qui dirige la traduction des polyprotéines. Le cadre de lecture ouvert unique du génome est organisé en 1ABCD / 2ABC / 3ABCD, les numéros indiquant les 3 domaines différents et chaque lettre représentant une polyprotéine.
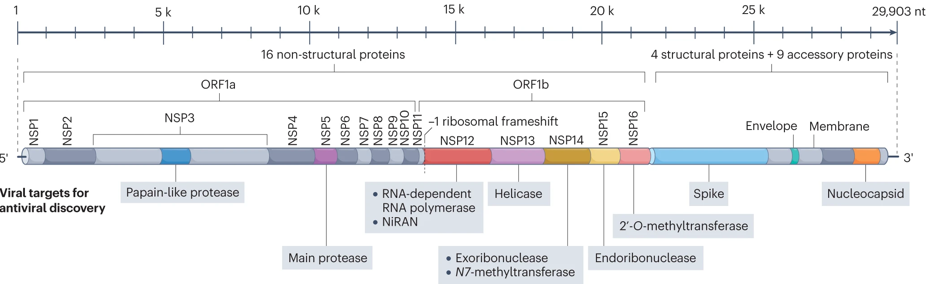
Ces polyprotéines sont des entités biologiques remarquables : elles possèdent toutes les propriétés (structurales et fonctionnelles) nécessaires à ces étapes du cycle de vie du virion. Voir un détail de l'une d'entre elles. Illustration : génome à ARN monocaténaire à sens positif du SARS-CoV-2
Source : Li et al. (2023) Les cibles des médicaments antiviraux sont indiquées sous la carte du génome. Protéines accessoires non cartographiées. NiRAN : domaine nucléotidyltransférase associé au nidovirus RdRp; NSP : protéine non structuralle; ORF : cadre de lecture ouvert. |
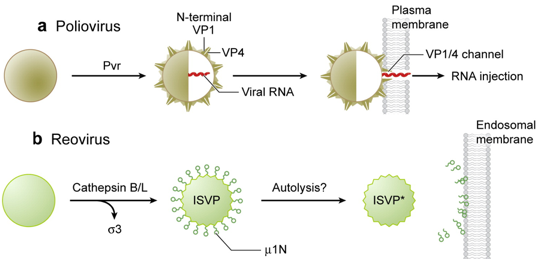
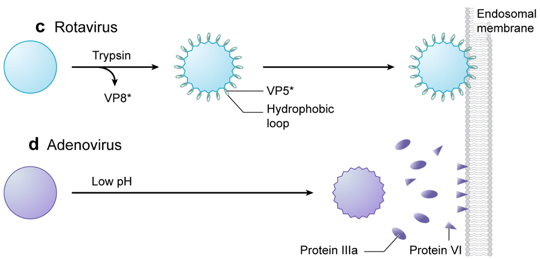
b. Mécanismes de perturbation des membranes pour la libération du contenu de la capside L'extrémité N-terminale de VP1 est un domaine phospholipase A2 (PLA2) : l'activité PLA2 réarrange la bicouche lipidique de l'endosome de manière transitoire et sur une échelle limitée. La protéine μ1 (76 kDa, protéine majeure de la capside des Réovirus de mammifères) est hydrolysée (autocatalytise) en deux fragments protéiques associés au virion : μ1N (4 kDa, myristoylé) et μ1C (72 kDa). La protéine μ1N forme des pores sélectifs dans la membrane endosomale. Il n'est pas certain que ces pores soient liés à la lyse osmotique de l'endosome, mais ils pourraient expliquer que le gros noyau de ce type de virus soit transfèré dans le cytoplasme.
La protéine VI (issue de la pré-protéine VI par protéolyse) des Adenovirus s'insère dans la foliole de la membrane endosomale, induisant une courbure positive de celle-ci et une fragmentation totale de l'endosome.
Figures adaptées de : Tsai B. (2007) |
| Famille / type de génome |
Site de pénétration | Protéine lytique | Domaine fonctionnel | Evènement déclanchant la libération | Virus | Mécanisme de lyse |
| Nodavirus / ARN simple brin (+) |
endosome | peptide γ |
APHa | pH faible | FHV | formation de pore ou fragmentation |
| Picornavirus / ARN simple brin (+) |
endosome | VP1 / VP4 | APH (VP1) / Myr (VP4) | fixation à un récepteur | PV | formation de pore |
| endosome | VP1 / VP4 | APH (VP1) / Myr (VP4) | pH faible | groupe mineur HRV | formation de pore | |
| endosome | VP1 / VP4 | APH (VP1) / Myr (VP4) | pH faible | groupe majeur HRV | fragmentation | |
| Reovirus / ARN double brin |
endosome | μ1N (μ1 -> μ1N+μ1C, autocatalytique) |
Myr | hydrolyse par les cathépsines B et L | reovirus de mammifères | formation de pore - lyse par osmose |
| membrane plasmique | hydrolyse trypsique de VP4 en VP8 et VP5 | résidus hydrophobes | hydrolyse trypsique / chélation du calcium |
rotavirus | fusion membranaireb | |
| Parvovirus / ADN simple brin |
endosome | VP1 | phospholipase A2 | pH faible / chélation de cations divalents |
AAV/CPV/MVM | réarrangement transitoire des lipides membranaires |
| Polyomavirus / ADN double brin |
réticulum endoplasmique | VP2 | Myr | pH faible / réarrangement par des chaperones |
polyomavirus murin | formation de pore |
| Adenovirus / ADN double brin |
endosome | VI | APH | pH faible et/ou fixation à une intégrine | HAdV | incurvation de la membrane - fragmentation totale ? |
|
aAPH : hélice α amphipathique; Myr : groupe myristoyle Source : Moyer & Nemerow (2011) |
|
|||||
|
Les virus qui infectent une bactérie doivent tout d'abord traverser une enveloppe dont la structure et la composition sont complexes. L'enveloppe des bactéries Gram(-) est généralement constituée d'une membrane cellulaire, d'une paroi mince de peptidoglanes périplasmiques recouverte d'une membrane externe contenant des lipopolysaccharides. L'enveloppe des bactéries Gram(+) n'a pas de membrane externe, mais sa couche de peptidoglycane est beaucoup plus épaisse et contient les polymères d'oses de la paroi cellulaire. De plus, ces structures peuvent être entourées d'autres structures protectrices telles que des polysaccharides de la capsule (exemples : l'acide polysialique, l'acide hyaluronique, des protéines de la couche S ou les acides mycoliques).
Dégradation de la capsule De nombreuses enveloppes bactériennes sont protégées par un réseau de polysaccharides (la capsule) qui les protège de l'environnement et empêche les virus d'atteindre leur récepteur d'entrée à la surface de ces cellules. En conséquence, les virus qui infectent des bactéries encapsulées possèdent une enzyme qui dégrade l'exopolysaccharide pour atteindre la membrane externe ou la couche de peptidoglycane. La capsule peut être composée d'alginates, d'acides polysialiques et de l'acide hyaluronique. Exemples de dépolymérases codées par certains phages qui dégradent la capsule :
|
5. Intégration du génome viral Il existe 3 types d'intégration du matériel génétique viral dans les chromosomes de la cellule hôte.
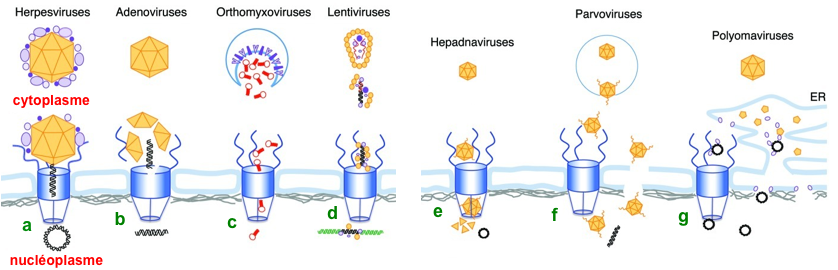
De nombreux virus n'entrent pas dans le noyau. Exemples :
D'autres virus se répliquent dans le noyau et doivent passer la barrière de l'enveloppe nucléaire lors de l'infection. Il existe plusieurs mécanisme qui dépendent du type de virus (figure ci-dessous). (a) Dans le cas du virus de l'herpès, la capside à laquelle sont attachées les protéines du tégument interne arrive au complexe du pore nucléaire (CPN). Après changement de conformation et ouverture de l'anneau "portail" au sommet de la capside, l'ADN est éjecté dans le noyau. (b) Cas des adénovirus : après avoir été relargué de l'endosome, la capside de l'adénovirus est amarrée au CPN, où des moteurs moléculaires perturbent la capside et la structure du CPN, libérant ainsi l'ADN viral dans le noyau. (c) Les ribonucléoprotéines des orthomyxovirus sont libérées de l'endosome dans le cytoplasme après la fusion de l'enveloppe virale (enrichie en glycoprotéines virales) avec la membrane de l'endosome. Les ribonucléoprotéines diffusent vers le CPN et elles entrent dans le noyau par un transport actif à l'aide de karyophérine (importines, exportines).
Figure adaptée de : Kobiler et al. (2012) - ADN viral en noir; ADN hôte en vert; ARN viral en rouge; CPN en bleu (d) Après désassemblage dans le cytoplasme, l'ARN des lentivirus est rétrotranscrit en ADN double brin. Le complexe [ADN - protéines virales], appelé complexe de pré-intégration, favorise l'entrée de l'ADN viral dans le noyau (par interaction avec les protéines du CPN). Celui-ci s'intègre alors dans le chromosome de l'hôte. (e) Les capsides des hépadnavirus pénètrent dans le CPN. Elles sont cependant trop volumineuses pour passer au travers des filaments qui adoptent une disposition dite en panier ("basket") et elles ne peuvent pas penétrer dans le noyau. Les capsides matures contenant l'ADN sont donc désassemblés dans le panier ce qui libère le génome viral circulaire dans le nucléoplasme. (f) Les capsides de parvovirus pénètrent intactes dans le noyau. Le domaine N-terminal de VP1 est extrudé dans l'endosome, ce qui déclenche l'activité de la phospholipase A. L'extrusion de l'extrémité N-terminale expose également 4 domaines signaux de localisation nucléaire qui semblent participer à l'entrée dans le noyau par le biais du NPC. Un autre modèle suggère une entrée directe dans le noyau via des perturbations locales de l'enveloppe nucléaire. (g) Les polyomavirus sont désassemblés dans le réticulum endoplasmique. Le génome "nu" quitte ce compartiment via les viroporines créées par les protéines de la capside interne. Il existe deux mécanismes : l'un provient directement de la lumière du réticulum endoplasmique à travers la membrane nucléaire interne et l'autre via le cytoplasme et le CPN. Les viroporines sont une famille de petites protéines hydrophobes de nombreux virus enveloppés. Elles interagissent avec différentes membranes cellulaires et s'auto-assemblent pour former des pores qui transportent des ions. |
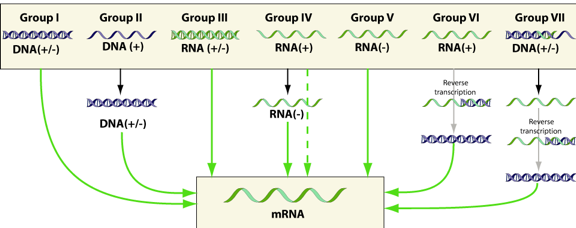
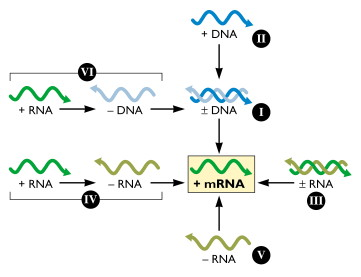
6. Réplication et transcription du génome viral, traduction des protéines virales Ces processus sont aussi divers qu'il existe de classes de virus donc de types de génome : ADN (+/-) ou ARN (+/-), simple brin ou double brin, linéaire ou circulaire, plusieurs segments ou pas, ... La figure suivante résume les "opérations" pour aboutir aux ARN messagers selon le type de génome viral d'origine :
Source : "Virology blog" Racaniello V. |
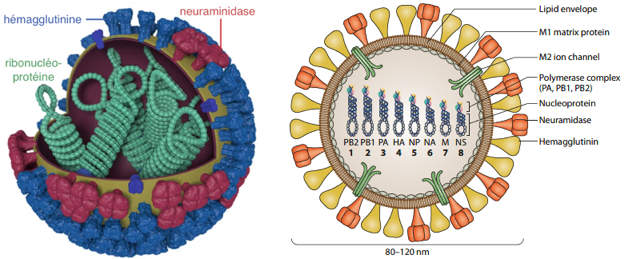
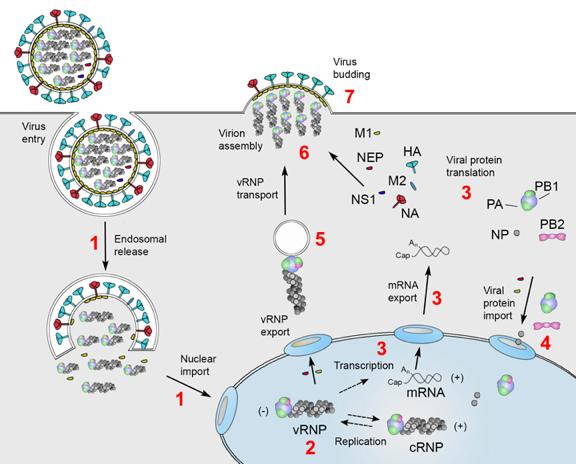
a. Exemple du virus de la grippe (influenza virus) 1. Après la fusion des membranes virale et endosomale, les ribonucléoprotéines virales (vRNP, "viral ribonucleoproteins") sont libérées dans le cytoplasme de la cellule hôte puis transportées dans le noyau (figure ci-dessous). Le terme ribonucléoprotéine désigne l'ARN génomique encapsidé. Dans le noyau :
Figure adaptée de : Te Velthuis & Fodor (2016) - HA: hémagglutinine; M1 et M2 : protéines de la matrice; NA : neuraminidase 4. Les sous-unités d'ARN polymérases virales nouvellement synthétisées (trimère constitué des polymérases basiques 1 et 2, PB1 et PB2, et de la polymérase acide, PA) et la nucléoprotéine (NP) sont à leur tour importées dans le noyau : elles se lient aux segments génomiques de vRNA et aux cRNA pour assembler les vRNP et les cRNP, respectivement. NP encapside le vRNA et le protége des nucléases. 5. Les vRNP nouvellement synthétisées sont exportées dans le cytoplasme. Elles sont transportés (de manière dépendante de RAB11) dans des endosomes de recyclage vers la membrane cellulaire. 6. L'assemblage des nouveaux virions s'effectue au niveau de la membrane cellulaire en incorporant de nombreuses protéines de la cellule hôte. 7. Enfin, les virions matures sont libérés par bourgeonnement. |
|
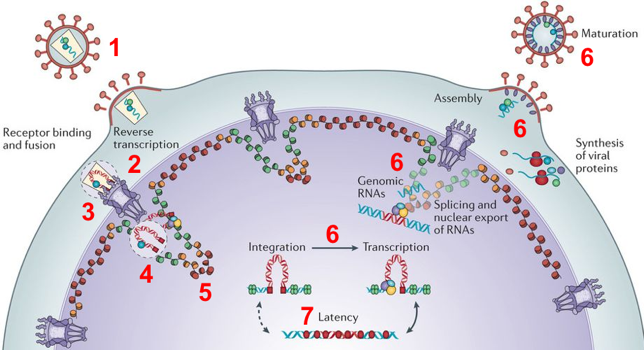
1. Le cycle du rétrovirus HIV-1 commence par la liaison des glycoprotéines de l'enveloppe virale aux récepteurs de la cellule hôte, suivie de la fusion des membranes virale et cellulaire puis de la libération du "noyau viral" dans le cytoplasme de la cellule hôte (figure ci-dessous). Le "noyau viral" contient :
2. L'ARN viral subit une transcription inverse en ADN par la transcriptase inverse virale et la capside est désassemblée. 3. L'ADN viral nouvellement formé est assemblé sous forme d'un complexe de pré-intégration ("pre-integration complex", CPI) avec l'intégrase et certaines protéines de la capside et cellulaires.
Figure adaptée de : Lusic & Siliciano (2017) 4. Le CPI entre dans le noyau au travers des complexes de pores nucléaires. L'entrée s'effectue dans des cellules qui ne sont pas en division : les chromosomes y sont séparés du cytoplasme par l'enveloppe nucléaire. 5. Le CPI assure l'intégration de l'ADN viral dans le génome de la cellule hôte :
6. Si l'infection est productive, les transcrits viraux sont épissés et exportés du noyau. Il s'en suit l'assemblage de nouvelles particules virales qui sont produites via la membrane plasmique et deviennent infectieuses après maturation. 7. Alternativement, l'ADN viral intégré dans le génome de la cellule hôte peut être temporairement "silencieux" via plusieurs mécanismes qui aboutissent à la formation de réservoirs viraux "en attente" (phénomène de latence pro-virale). |
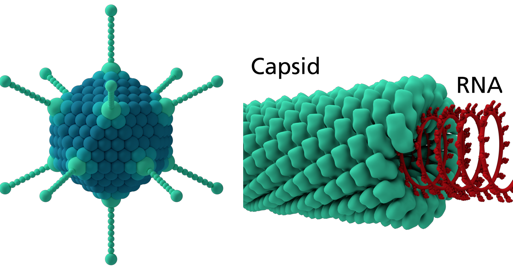
7. La capside À quelques exceptions près, les capsides des virus non enveloppés sont construites à partir d'une protéine de capside dite majeure (qui détermine l'assemblage et l'architecture du virion) et de une ou plusieurs protéines de capsides dites mineures. Les nucléocapsides des virus enveloppés sont souvent construites à partir d'un complexe nucléoprotéique [protéine - génome viral], des protéines de la matrice (qui relient la nucléoprotéine à la membrane lipidique) et des protéines d'enveloppe (pour la reconnaissance de l'hôte et la fusion membranaire.) Les capsides peuvent être icosaédrales (figure de gauche, ci-dessous), hélicoïdales (figure de droite) ou complexes :
Source : Splettstoesser T. (SCIstyle)
Source : "Assigment expert" |
| Exemples de virus | Hôte | Diamètre de la capside | Nombre de la triangulation | Type de génome et taille approximative |
| Procaryotes - bactéries | ||||
| PRD1, Tectiviridae | Bacterie Gram(-) | 70 nm | T = 25 | ADN linéaire double brin, 15 kilo paires de bases (kpb) |
| PM2, Corticoviridae | Pseudoalteromonas | 60 nm | T = 21d | ADN circulaire double brin, 10 kpb |
| Salisaeta phage 1 icosahèdral (SSIP-1), Sphaerolipoviridae ?, non classé | Salisaeta sp | 100 nm (repliement en "single jelly roll" ?) | T = 49 | ADN circulaire double brin, 44 kpb |
| Infection des Flavobacterium, phage riche en lipides (FLiP), non classé | Flavobacterium sp. | 55 nm | T = 21d | ADN circulaire simple brin, 9 kb |
| Procaryotes - archées | ||||
| STIV, Turriviridae | Sulfolobus solfataricus | 73 - 96 nm | T = 31d | ADN circulaire double brin, 18 kpb |
| HHIV-2, Sphaerolipoviridae | Haloarcula hispanica | 80 nm ("single jelly roll") | T = 28d | ADN linéaire double brin, 30 kpb |
| Eucaryotes | ||||
| Paramecium bursaria chlorella virus 1 (PBCV-1), Phycodnaviridae | Chlorella variabilis | 190 nm | T = 169d | ADN double brin, 330 kpb |
| virus 1 Phaeocystis pouchetii (PpV01), Phycodnaviridae ou Mimiviridae | Phaeocystis pouchetii (phytoplancton) | 220 nm | T = 219 | 485 kpb |
| virus Cafeteria roenbergensis, Cafeteriavirus, Mimiviridae | Cafeteria roenbergensis (zooplancton) | 300 nm | T = 499 | 730 kpb |
| Acanthamoeba polyphaga Mimivirus (APMV), Mimivirus, Mimiviridae | Acanthamoeba polyphaga | 500 nm | T = 972 - 1200 | ADN linéaire double brin, 1180 kpb |
| Sputnik, Lavidaviridae | Amoebae / Mimivirus (virophage) | 75 nm | T = 27 | ADN circulaire double brin, 18 kpb |
| Melbournevirus, Marseilleviridae | Acanthamoeba castellanii | 230 nm | T = 309 | ADN double brin, 369 kpb |
| Faustovirus, non classé | Vermamoeba vermiformis | 260 nm | T = 277 | ADN circulaire double brin, 466 kpb |
| Pacmanvirus, non classé | Acanthamoeba castellanii | 250 nm | T = 309 | ADN double brin, 395 kpb |
| virus Chilo iridescent (CIV), Iridoviridae | Invertébrés, amphibiens, poissons | 185 nm | T = 147 | ADN linéaire double brin, 212 kpb |
| Adenovirus, Adenoviridae | Vertébrés | 95 nm | T = 25 | ADN linéaire double brin, 27 - 43 kpb |
| virus Vaccinia, Poxviridae | Vertébrés | 200 - 300 nm | Non icosahèdral | ADN linéaire double brin, 130 - 375 kpb |
| Source : San Martin & van Raaij (2018) | ||||
|
Un grand nombre de virus (environ 38,5%) possèdent des protéines de la capside qui adoptent un repliement appelé "jelly roll" :
Au moins 16 familles de virus (à ARN et à ADN) contiennent des protéines de capside (principalement icosaédriques) qui adoptent un repliement "jelly roll" unique :
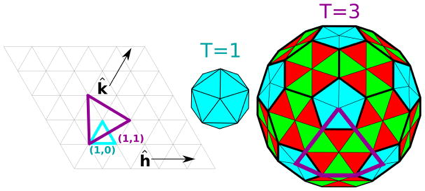
Certaines familles de virus construisent leurs capsides avec des protéines qui adoptent un repliement "jelly roll" double (2 repliements simples reliés), voire triple. c. Le nombre de triangulation T La description des structures des capsides virales en forme de sphère repose sur le "principe de quasi-équivalence" de D. Caspar & A. Klug (A. Klug a obtenu le Prix Nobel de chimie en 1982) :
De manière générale Le nombre de triangulation T est une métrique utile pour la mesure quantitative de la taille d'une capside : dans la plupart des cas, une capside de nombre de triangulation T est constituée de 60T sous-unités, ou 12 pentamères et 10 (T-1) hexamères. Ces structures caractérisent la grande majorité des virus sphériques, par exemple la capside du bromovirus CCMV ("cowpea chlorotic mottle virus") qui a une structure T = 3.
Source : Perlmutter & Hagan (2015) |
d. Description de quelques protéines de capsides Les polyomavirus sont des petits virus non enveloppés ayant une capside de symétrie icosaédrique et un diamètre d'environ 45 nm. Les virions contiennent un génomes à ADN double brin circulaire d'une longueur d'environ 5000 paires de bases : l'ADN est associé à des histones et code pour 5 à 6 protéines. Leur enveloppe protéique est constituée principalement de 360 molécules de protéine VP1, assemblées en 72 pentamères reliés par des ponts disulfure :
Les protéines mineures de la capside, VP2 et VP3 (qui ont une forte similarité de séquence), sont attachées à la cavité intérne centrale du pentamère VP1 :
|
| Caractéristiques des protéines de la capside de petits virus à ADN non enveloppés | ||||
| Caractéristique | Adenoviridae | Parvoviridae | Papillomaviridae | Polyomaviridae |
| PCPMa | VI | VP1 | L2 | VP2, VP3 |
| proportion des PCPM par rapport à toutes les protéines de la capside | 5% | 10% | 12% | 14% |
| PCPM exposée en surface (>15%) | non | non | non | non |
| mode de sortie du génome viral de la vésicule formée | endosome | endosome | réseau Trans-Golgi ? |
réticulum endoplasmique |
| Conditions pour l'exposition des PCPM | désassemblage partiel de la capside et relargage d'autres protéines de la capside | température élevée, faible pH, hydrolyse d'autres protéines de la capside | Interaction de la capside avec des récepteurs, enzymes extracellulaires | enzymes du réticulum endoplasmique, interaction avec la membrane |
| association PCPM - capside pendant la solubilisation de la membrane | non | oui | non (mais les PCM s'associent au génome) | pas connu |
| mécanisme de pénétration dans la membrane | interaction avec la membrane | enzymes | interaction avec la membrane | interaction avec la membrane |
| conformation de la protéine nécessaire pour la pénétration membranaire | hélices α amphipathiques | hélices α | segment transmembranaire | segment transmembranaire |
aPCPM ("Membrane Penetration Capsid Protein") : protéines de la capside qui permettent de pénétrer dans la membrane. Source : Bilkova et al. (2014) |
||||
8. Assemblage de la capside et empaquetage du génome L'assemblage de la capside désigne la formation de la coquille de la capside et l'empaquetage du génome désigne l'insertion et la disposition du génome viral dans la capside ou dans l'enveloppe. Ces processus sont également très divers et dépendent du type de virus :
|
| Les virus à ARN simple brin (tels que le bactériophage MS2) co-assemblent leur capside avec le génome. | La plupart des capsides s'auto-assemblent spontanément autour du génome viral dans le cytoplasme. |
| Les bactériophages à ADN double brin volumineux, les virus de l'herpès, les adénovirus et les microvirus disposent d'un mécanisme de translocation de l'ADN qui encapside le génome viral dans une capside préassemblée ou procapside. | Les capsides hélicoïdales sont assemblées autour de l'ARN ou de l'ADN génomique. Cet assemblage repose sur des interactions entre les capsides et sur des interactions avec l'acide nucléique à empaqueter : l'assemblage et l'empaquetage sont liés. |
| Les capsides de structure complexe nécessitent des protéines "d'échafaudage" ("scaffolding proteins") et sont assemblées dans des procapsides vides. Les protéines d'échafaudage sont retirées de la capside (événements de maturation) avant l'empaquetage. | Les capsides des virus Reoviridae et Totiviridae sont assemblées autour d'ARN messagers qui sont ensuite répliqués sous forme d'ARN génomique double brin, les masquant ainsi des molécules antivirales cellulaires. |
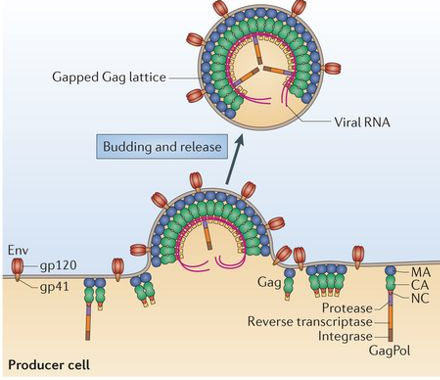
| La particule du rétrovirus HIV est grobalement sphérique et mesure environ 100 nm de diamètre. Elle est initialement assemblée sous forme d'un virion immature contenant l'ARN et construit principalement à partir d'environ 2000 copies de la protéine Gag. | Le génome des virus à ARN (-) est encapsidé pendant la réplication. L'empaquetage de ces virus se produit avant le bourgeonnement au niveau de la membrane plasmique. |
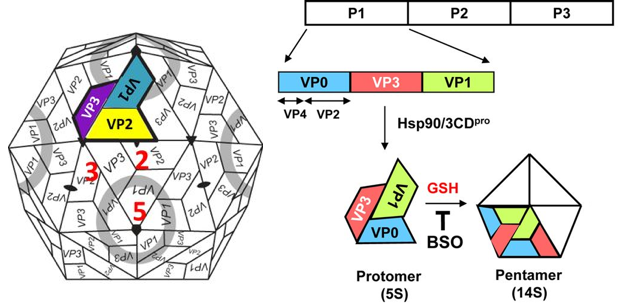
a. Illustration : modèle de morphogenèse d'entérovirus
Source : Jiang et al. (2014)
|
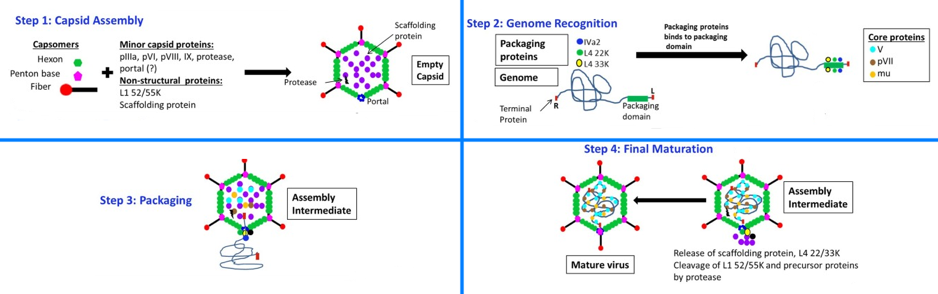
b. Illustration : modèle pour l'assemblage de la capside et l'empaquetage des adénovirus Étape 1 : les capsides icosaédriques vides sont assemblées à partir :
Étape 2 - reconnaissance du génome : les protéines d'empaquetage IVa2, L4 33K, L1 52/55K et L4 22K se lient au domaine d'empaquetage situé près d'une extrémité du génome (ADN double brin). Le génome est associé à la protéine terminale pTP à chaque extrémité. L et R indiquent les extrémités gauche et droite du génome viral, respectivement.
Figure adaptée de : Ahi & Mittal (2016) Étape 3 - conditionnement : le génome est inséré dans les capsides vides à travers une protéine portail située au sommet par l'action de IVa2, L4 22K et L4 33K. Les protéines centrales V, VII et mu sont incorporées dans les capsides vides pendant ou après l'incorporation du génome. Étape 4 - maturation finale : les protéines d'échafaudage et les protéines L4 22K et 33K sont libérées pendant ou après l'incorporation du génome. La protéase codée par le virus clive ses substrats pIIIa, L1 52/55K, pVI, pVII, pVIII, mu et pTP, entraînant des changements de structure de la capside et la maturation de la particule virale. |
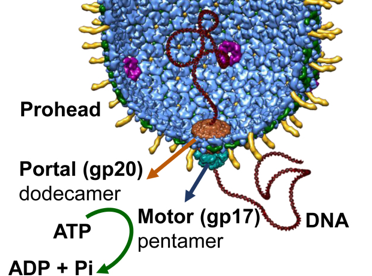
c. Illustration : capside pré-formée et complexe "portail" des bactériophages Chez les bactériophages à ADN et les virus de l'herpès, la condensation du génome viral dans les capsides préformées est une réaction non spontanée catalysée par un complexe protéique d'empaquetage du génome constitué :
Le portail est en forme de cône : il relie la tête au moteur et à l'ADN. Son extrémité plus large est à l'intérieur de la capside et son extrémité plus étroite fait saillie vers l'extérieur. Il forme un canal central d'un diamètre d'environ 35 Å, au travers duquel l'ADN entre dans la capside.
Source : Lin et al. (2017) Mécanisme supposé
|
9. Sortie du génome viral du noyau de la cellule hôte Tous les virus qui se répliquent dans le noyau doivent exporter leur matériel génétique. Ainsi, pour assembler leurs virions "progéniture", ces virus nécessitent un mécanisme d'export du noyau pour transporter le complexe de ribonucléoprotéines virales vers le cytoplasme (avant son transfert vers la membrane plasmique où le processus d'assemblage aura lieu). Trois voies principales permettent à ce type de virus de sortir du noyau. a. Export du génome par les pores nucléaires et une protéine navette L'export de macromolécules à travers l'enveloppe nucléaire nécessite une séquence consensus riche en leucine : Hyd1-X(2,3)-Hyd2-X(2,3)-Hyd3-X-Hyd4, où Hyd représente des résidus hydrophobes (leucine, isoleucine, valine ou méthionine) et X représente n'importe quel acide aminé. Cette séquence consensus s'appelle signal d'export du noyau ("Nuclear Export Signal" - NES). Elle est reconnue par le récepteur de transport cellulaire appelé "Chromosome Region maintenance 1" (CRM1). La protéine Rev du virus HIV-1 contient une séquence signal d'export du noyau de l'ARN viral non épissée. La protéine NS2 du virus de la grippe était supposée intervenir dans l'export du noyau de ribonucléoprotéines virales : elle a été baptisée "Nuclear Export Protein" (NEP). |
| Export du génome par les pores nucléaires | ||
| Virus | Type de génome | Protéine d'export |
| Baculoviridae | ADN encapsidé | ac66 ("desmo-N domain-containing protein") |
| Bornaviridae | ADN encapsidé | p23 ou p24 (phosphoprotéine) |
| Caulimoviridae | ARN pré-génomique non épissé | protéine de la capside ? |
| Deltavirus | ARN simple brin | L-HDag |
| Geminiviridae | ADN simple brin | protéine motrice |
| Hepadnaviridae | ARN pré-génomique non épissé | protéine C de la capside ? |
| Nanoviridae | ADN | protéine DNA-M motrice potentielle |
| Parvoviridae | ADN encapsidé | NS2 (parvovirus) |
| Orthomyxoviridae | ADN encapsidé | NEP ("Nuclear Export Protein" - NS) |
| Lentivirus | ARN pré-génomique non épissé | Rev |
| Alpharetrovirus | ARN pré-génomique non épissé | Gag |
| Sources : ViralZone - Uniprot | ||
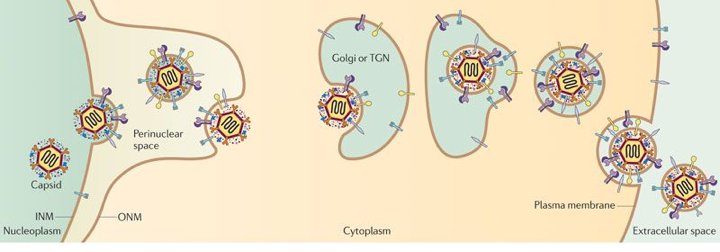
b. Export par remodelage des membranes nucléaires ("nuclear egress") Les virus de l'herpès possèdent de grands génomes (de 125 à 235 kilo paires de bases, codant jusqu'à plus de 200 protéines). Les nucléocapides virales sont donc trop grandes (jusqu'à 125 nm) pour traverser les pores nucléaires. L'export du noyau de ce type de virus est en conséquence inhabituelle pour des virus enveloppés car leurs capsides naissantes sortent du noyau en bourgeonnant 2 fois avec la membrane nucléaire tout en n'acquérant qu'une seule enveloppe.
Source : Johnson & Baines (2011) - TGN : réseau trans-Golgi Le complexe viral de sortie nucléaire ("viral Nuclear Egress Complex", NEC1 ou UL31 et NEC2 ou ul34) est un acteur essentiel de la sortie du noyau par ce mécanisme. Le NEC recouvre les membranes en formant un manteau en "nid d'abeille" ("honeycomb coat") : c'est donc un système de bourgeonnement de la membrane nucléaire (par opposition au système de bourgeonnement de la membrane cytoplasmique). |
c. Perturbation de l'enveloppe nucléaire Les nucléocapsides virales peuvent s'échapper du noyau en perturbant et en dégradant l'enveloppe nucléaire. Ce processus peut avoir lieu avant toute lyse cellulaire et, pour certains virus, l'assemblage du virion est terminé dans le cytoplasme. La perturbation de l'enveloppe nucléaire est un processus actif induit par des protéines virales tardives spécifiques. Les parvovirus sont des petits virus à ADN non enveloppés. Malgré leur petite taille, ils ne traversent pas les pores nucléaires mais sont liés aux protéines du complexe de pores nucléaires. Cette liaison modifie la structure des parvovirus et ce réarrangement structural est essentiel pour déclencher une cascade de signaux qui dégrade l'enveloppe nucléaire. Cette dégradation se produit au cours de la prophase de la mitose. L'exposition de domaines riches en hélices amphipathiques est nécessaire à la dégradation de l'enveloppe nucléaire (perturbation de la membrane nucléaire interne et externe). L'efflux de Ca2+ à partir de la lumière, entre les membranes nucléaires interne et externe, indique que ce cation est essentiel au désassemblage nucléaire en activant la protéine kinase C. Une fois activée, elle déclenche à son tour l'activation de cdk-2 (ensuite sur-activée par la caspase-3). L'interaction [virus - enveloppe nucléaire] montre qu'un ensemble d'enzymes permet le désassemblage de l'enveloppe nucléaire dans des cellules quiescentes. |
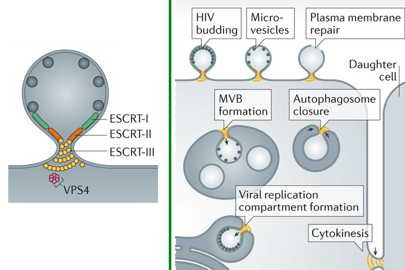
10. Bourgeonnement et sortie de la cellule hôte avec les machineries ESCRT Beaucoup de virus enveloppés sortent des cellules infectées en bourgeonnant au travers des membranes et acquièrent ainsi leurs bicouches lipidiques (l'enveloppe). Certains de ces virus (notamment le rétrovirus dont le virus HIV-1) emploient une stratégie qui consiste à usurper la voie cellulaire ESCRT ("Endosomal Sorting Complexes Required for Transport"). Les machineries ESCRT sont composées de 3 sous-complexes (ESCRT-I, -II et -III).
Source : Schoneberg et al. (2017)
Certains virus bourgeonnent pas des mécanismes indépendants de ESCRT. |
|
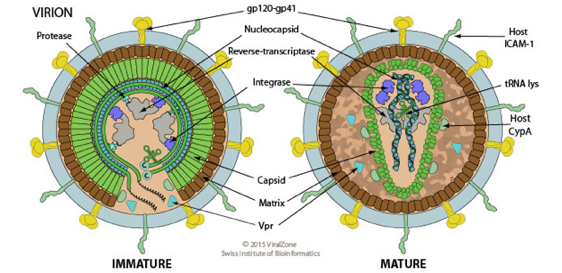
La maturation des virions est un processus qui a lieu après la séparation de la particule infectieuse virale de la cellule hôte par bourgeonnement ou scellement de la capside sphérique. Elle peut également se produire à l'intérieur de la cellule hôte. Voir la définition selon l'ontologie. La maturation consiste en des modifications du repliement des protéines virales et/ou un clivage irréversibles des protéines virales qui activent les virions afin qu'ils deviennent à leur tour compétents pour la réinfection. La maturation correspond donc aux très profonds changements de structure et aux changements biochimiques programmés entre la formation de la particule initiale et le développement du virion infectieux.
Source : ViralZone Figure ci-dessus : virion d'un lentivirus enveloppé, sphérique, de 80 à 100 nm de diamètre. La capside mature est constituée de 1572 protéines. La maturation est un processus qui se produit chez la grande majorité des virus animaux et dans tous les bactériophages. La dernière étape de la maturation d'un virus animal implique fréquemment :
|
a. La transcriptase inverse ("reverse transcriptase") des rétrovirus La majorité des rétrovirus compétents pour la réplication dépendent de 3 gènes : le gène "antigène spécifique de groupe" ("group specific-antigen", gag), le gène polymérase (pol) et le gène enveloppe (env). La structure du génome d'un rétrovirus est schématiquement : 5'LTR-gag-pol-env-LTR3' (figure ci-dessous).
Source : Azevedo-Pereira et al. (2015) - Intechopen
Les rétrovirus utilisent donc une transcriptase inverse pour répliquer leurs génomes. Ce type d'enzyme a été découverte indépendamment en 1970 par Howard Temin et David Baltimore. Une transcriptase inverse (E.C. 2.7.7.49, "RNA-directed DNA polymerase") est une enzyme qui synthétise un brin d'ADN complémentaire (ADNc double brin) à partir d'un brin matrice ARN simple brin et ce processus est appelé transcription inverse. Une transcriptase inverse possède 3 activités enzymatiques : une activité ADN polymérase dépendante de l'ARN, une activité ribonucléase H (qui dégrade les régions U5 et R situées à l’extrémité 5' de l’ARN matrice) et une activité ADN polymérase dépendante de l'ADN.
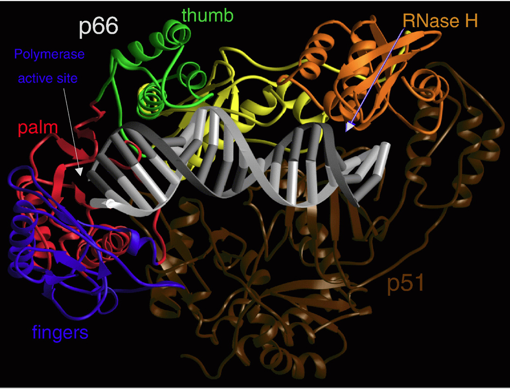
La transcriptase inverse du virus HIV-1 est un hétérodimère asymétrique composé de 2 sous-unités apparentées, appelées p66 (560 acides aminés) et p51 (440 acides aminés). Ces deux sous-unités sont issues de l'hydrolyse, par la protéase de ce même virus, d'une polyprotéine Gag-Pol synthétisée à partir de l'ARN viral non épissé.
Source : Sarafianos et al. (2009)
|
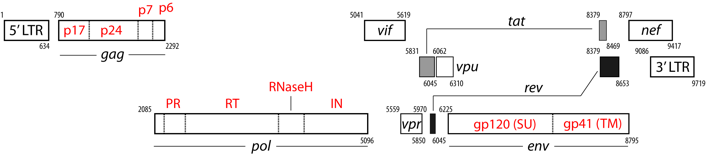
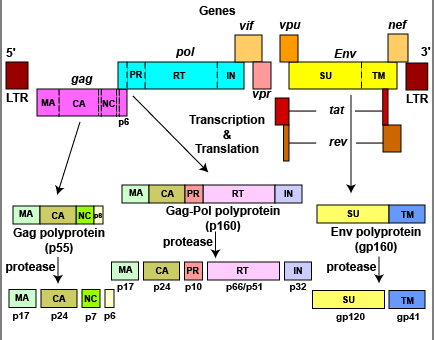
b. Maturation des poly-protéines Gag-Pol et Gag La transcriptase inverse rétrovirale est synthétisée sous forme d'un précurseur qui s'appelle poly-protéine Gag-Pol qui subit un clivage protéolytique pour donner les protéines matures. La poly-protéine Gag-Pol (P03366, aussi appelée Pr160Gag-Pol) est en effet clivée en 11 chaînes polypeptidiques : Matrix protein p17 (MA); Capsid protein p24 (CA); Spacer peptide 1; Nucleocapsid protein p7 (NC); Transframe peptide; p6-pol; aspartyl protease (E.C. 3.4.23.16; HIV-1 retropepsin); p66 reverse transcriptase/ribonuclease H (E.C. 2.7.7.49, E.C. 2.7.7.7, E.C. 3.1.26.13); p51 reverse transcriptase; p15; Integrase (E.C. 2.7.7).
Source :The Life Cycle of HIV La polyprotéine Gag-Pol associée à la polyprotéine Gag (P04591) sont responsables des événements essentiels de l'assemblage des virions, en particulier :
La poly-protéine Gag-Pol peut réguler sa propre traduction en fixant la région 5'-UTR (non traduite) de l'ARN génomique. La protéase à aspartate (aspartyl protease, E.C. 3.4.23.16) catalyse les clivages protéolytiques des poly-protéines Gag et Gag-Pol pendant ou peu après la libération du virion de la membrane plasmique (l'activité est maximale pendant le processus de bourgeonnement juste avant la libération des particules). Les clivages ont lieu par étapes (sous forme de cascade ordonnée) pour donner des protéines matures. La protéase du virus HIV-1 (O90777) mature est un homodimère de 22 kDa, chaque sous-unité étant composée de 99 acides aminés. Un seul site actif se situe entre les sous-unités identiques : il contient la triade catalytique Asp25-Thr26-Gly27, caractéristique de ce type de protéase. Cette protéase est une cible thérapeutique majeure d'un grand nombre d'inhibiteurs (médicaments) car son inhibition abolit le cycle viral. |
|
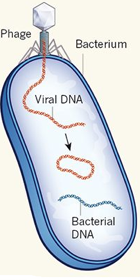
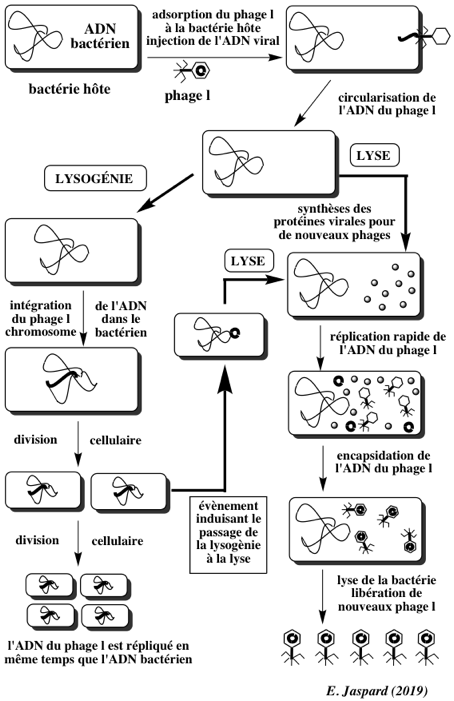
Cycle lysogène : cycle de réplication d'un phage au cours duquel le génome du virus est incorporé dans l'hôte bactérien sous forme de prophage. Ce cycle ne tue pas la bactérie hôte. Cycle lytique : cycle de réplication du virus qui libère de nouveaux phages après la lyse de la cellule hôte. Un phage virulent se reproduit par un cycle lytique. La lyse de la cellule résulte de la dislocation des membranes de la cellule infectée, entraînant sa mort et la libération des composés cytoplasmiques dans l'espace extracellulaire. La lyse est induite par de nombreux virus, car les cellules déclenchent rarement leur propre lyse : les cellules eucaryotes ont tendance à déclencher le processus d'apoptose lorsqu'elles sont attaquées par des virus. Il existe 3 principaux groupes de bactériophages :
Le bactériophage λ s'adsorbe à la paroi de la bactérie hôte et y injecte son ADN qui se circularise. Il y a ensuite deux possibilités :
|
13. Arbitrage entre lysogènie (voie latente) et voie lytique Ce "choix" est contrôlé par le taux respectif de protéines appartenant à des systèmes de "commutation génétique". Ce rapport dépend du type de cellule hôte, de sa forme ou de la disponibilité en nutriments.
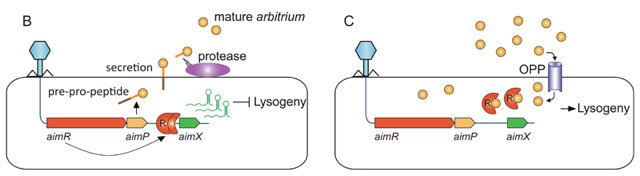
Exmple du le bactériophage phi3T Lors de la première rencontre d'un phage avec une population bactérienne, les gènes précoces aimR et aimP sont transcrits immédiatement après l'infection (figure B , ci-dessous).
Ultérieurement au cours de l'infection, ce peptide régulateur accumulé dans le milieu extracellulaire est internalisé dans la bactérie par le transporteur OPP (figure B , ci-dessous). Dès lors, lorsqu'un phage infecte la bactérie, la protéine AimR fixe ce peptide et ne peut plus activer la transcription du gène aimX : il en résulte une orientation vers la lysogénie.
Source : Erez et al. (2017) |
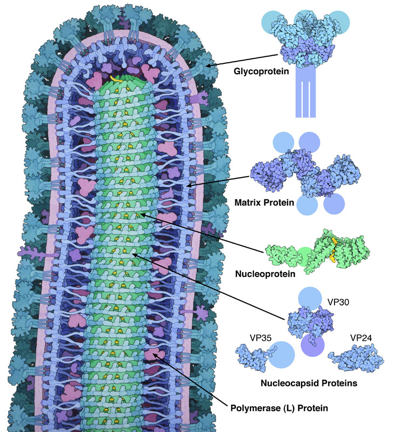
14. Les protéines du virus Ebola Le virion du virus Ebola entre dans la cellule hôte par macropinocytose. La formation de ce type de vésicules d'endocytose (appelée macropinosome) résulte d'une stimulation de la cellule entraînant la formation de plis à leur surface qui internalise le matériel extracellulaire. De nombreux virus utilisent cette voie d'entrée non spécifique après s'être fixé à la surface de la cellule hôte. Le génome du virus Ebola contient l'information pour la synthèse de 7 protéines qui s'assemblent avec l'ARN génomique pour former un virus hautement pathogène :
Source : PDB "Molecule of the month" La glycoprotéine synthétisée par Ebola (PDB 3CSY) se lie aux récepteurs situés à la surface des cellules cibles. Elle partage de nombreuses caractéristiques avec d'autres protéines de fusion de virus (exemples, l'hémagglutinine de l'influenza ou la glycoprotéine d'enveloppe du HIV). Elle recouvre la surface du virus. Elle est elle-même "recouverte" de chaînes glucidiques qui la "cachent" au système immunitaire. C'est une protéine dont la conformation est très dynamique. En effet, elle change de forme quand elle se fixe à la surface de la cellule cible, ce qui approche le virus et la cellule de sorte que leurs membranes fusionnent. La protéine de la matrice (hexamère) synthétisée par Ebola (VP40, PDB 4LDD) façonne le virus et dirige le processus de bourgeonnement. De nombreuses copies de la protéine s'associent sur la membrane et sont supposées établir des connexions à la fois avec la membrane et avec la nucléocapside. La nucléocapside (composée de plusieurs types de protéines) protège le génome viral :
|
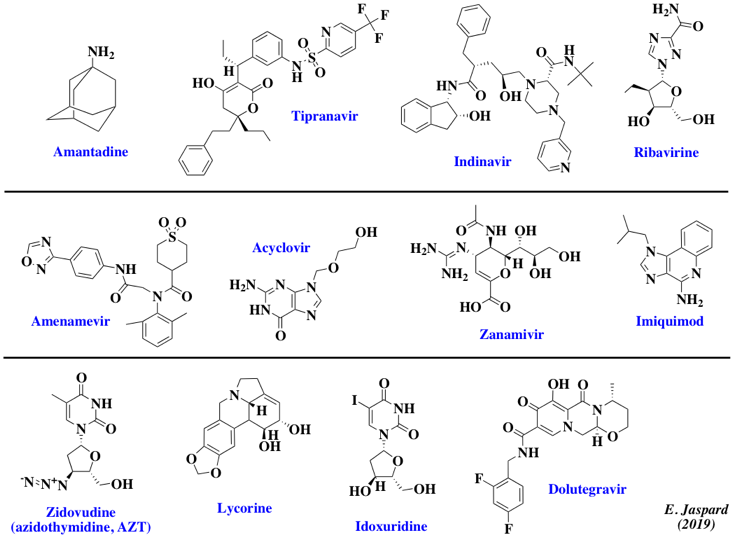
15. Médicaments antiviraux et développement de nouveaux antiviraux a. Exemples de quelques médicaments antiviraux
|
| Virus | type d'inhibiteurs |
| HIV | inhibiteurs de la protéase, de l'intégrase, inhibiteurs de l'entrée du virus dans la cellule hôte, inhibiteurs nucléosidiques de la transcriptase inverse, inhibiteurs non nucléosidiques de la transcriptase inverse, analogues de nucléosides phosphonates acycliques |
| hépatite B | lamivudine, interférons, analogues de nucléosides et analogues nucléosidiques phosphonates acycliques |
| hépatite C | ribavirine, interférons, inhibiteurs de la protéase NS3/4A, inhibiteurs de NS5A et NS5B |
| herpès | analogues 2'-désoxyuridine substitués en 5, inhibiteurs de l'entrée du virus dans la cellule hôte, analogues de nucléosides, analogues du pyrophosphate |
| grippe | ribavirine, inhibiteurs de la protéine 2 de la matrice , inhibiteurs de l'ARN polymérase et de la neuraminidase |
| cytomégalovirus humain | analogues acycliques de la guanosine, analogues du pyrophosphate et oligonucléotides (letermovir) |
| varicelle-zona | analogues acycliques de la guanosine, analogues de la 2′-désoxyuridine substituée et anticorps |
| virus respiratoire syncytial | ribavirine et anticorps |
| papillomavirus humain | imiquimod, sinécatéchines et podofilox |
| Source : De Clercq & Li (2016) | |
| Exemple d'inhibition d'étapes ou d'enzymes du cycle de vie des virus | ||
| Etape ou enzyme | Type d'inhibition / inhibiteur | Remarque |
| Fixation d'un virus à son récepteur spécifique à la surface de la cellule hôte | Ligands qui se fixent sur le récepteur, anticorps anti-récepteur ou, à l'inverse, analogues structuraux du récepteur lui-même | |
| Entrée d'un virus | Plus grande difficulté du virus à développer une résistance | |
| Etape de "déshabillage" ("uncoating") | Amantadine et rimantadine (virus de la grippe), Pleconaril (rhinovirus) | |
| Transcriptase inverse | Analogues de nucléotides ou de nucléosides : aciclovir, penciclovir (herpès), zidovudine (HIV), lamivudine (hépatite B) | aciclovir : analogue de la guanosine converti par la thymidine kinase virale en aciclovir monophosphate puis en aciclovir triphosphate par les kinases des cellules hôtes |
| Intégrase | dolutegravir, MK-2048, ... | |
| ribozymes ou ADN / ARN antisens | Fomivirsen : oligonucléotide antisens contenant un oligonucléotide phosphorothioate de 21 nucléotides (5'-GCG-TTT-GCT-CTT-CTT-CTT-GCG-3') | |
| Assemblage d'un virus | rifampicine | |
| Libération d'un virus | zanamivir et oseltamivir | inhibition de la neuraminidase du virus de la grippe |
| Complexe primase / hélicase | aménamevir ou pritelivir : inhibition du complexe hélicase / primase du virus de l'herpès | ST-610 |
|
La vipérine (viperin, "Virus Inhibitory Protein, Endoplasmic Reticulum-associated, INterferon-INducible") ou protéine inhibitrice de virus, associée au réticulum endoplasmique, inductible par l'interféron ou RSAD2 ("Radical S-Adenosyl methionine Domain-containing 2") appartient à la superfamille d'enzymes "radical SAM (S-adénosylmethionine) à centres [4Fe-4S]+" (PFAM PF04055). La vipérine est une enzyme multifonctionnelle inductible par différentes types d'interférons. Elle est impliquée dans l'inhibition de la réplication de nombreux virus à ARN et à ADN (exemples : virus de la dengue, West Nile, de l'hépatite C, de la grippe, de la rage, HIV, …). La vipérine catalyse la conversion de la cytidine triphosphate (CTP) en 3'-désoxy-3', 4'-didéhydro-CTP (ddhCTP) : le ddhCTP agit en tant que terminateur de chaîne pour les ARN polymérases ARN-dépendantes de plusieurs membres du genre Flavivirus et le ddhCTP inhibe directement la réplication du virus Zika. Ces activités antivirales étendues résulteraient d'interactions avec un grand nombre de protéines des cellules hôtes et des virus. La vipérine est également impliquée dans la régulation de la poly-ubiquitination de la Lys63 de la kinase 1 associée au récepteur de l'interleukine-1 ("Interleukin-1 Receptor-Associated Kinase-1", IRAK1) par l'ubiquitine-ligase E3 TRAF6. |
| Exemples de médicaments antiviraux potentiels susceptibles d'être repositionnés pour traiter la maladie à coronavirus 2019 (COVID-19) | ||
| Médicaments | Cible | Mécanisme |
| Remdesivir | ARN polymérase ARN-dépendante | Interfère avec les ARN polymérases virales pour inhiber la réplication virale. |
| Ribavirin | ARN polymérase ARN-dépendante | Inhibe la biosynthèse des nucléotides en inhibant l'ARN polymérase virale. |
| Favipiravir | ARN polymérase ARN-dépendante | Inhibe la transcription et la réplication du virus. |
| Lopinavir/Ritonavir | Principales protéases virales | Inhibe les protéases virales et bloque la multiplication. |
| Darunavir | Principales protéases virales | Inhibe la maturation du virus en inhibant les protéases virales. |
| Chloroquine/Hydroxychloroquine | Entrée du virus | Modifie le pH endosomal. Inhibe l'entrée du virus et interfère avec la fonction ARN polymérase dépendante de l'ARN. |
| Arbidol | Glycoprotéine de pointe ("Spike") | Inhibe l'entrée du virus et les étapes postérieures. |
| Source : Won & Lee (2020) | ||
|
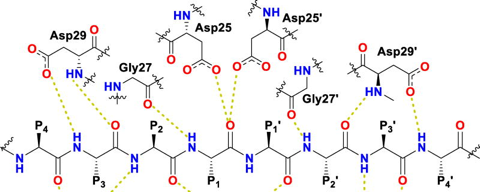
b. Exemples d'inhibiteurs de la protéase du HIV-1 Classification : "HIV-1 retropepsin", E.C. 3.4.23.16, PDB "Molecule of the month", PFAM PF00077. L'une des principales cibles (et l'une des premières) des antiviraux anti-HIV-1 sont les inhibiteurs de la protéase aspartique. Exemples : dérivés de pseudopeptides symétriques (Langlois, Quintard & Abalain, 1994) ou du dibenzyl-iminodiacétate (Abalain & Langlois, 1998). La protéase aspartique est biosynthétisée sous forme du précurseur polyprotéines Gag-Pol : elle est située entre p6pol (du côté N-terminal de la protéase) et la transcriptase inverse (du côté C-terminal). La protéase mature est un homodimère de 22 kDa (99 acides aminés par monomère) : les sous-unités forment un tunnel dans lequel est situé le site actif constitué des 2 séquences conservées Asp25 - Thr23 - Gly27 caractéristique des aspartyl protéases.
Source : Ghosh et al. (2016) Le médicament à base de tipranavir inhibe l'activité enzymatique et la dimérisation de la protéase du HIV-1. |
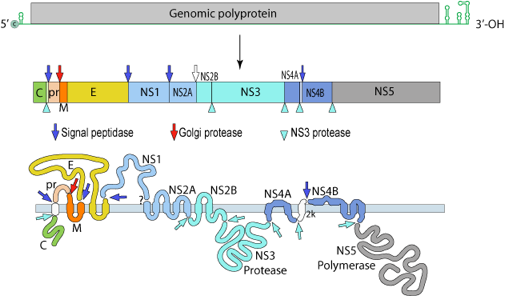
c. Traitement thérapeutique de la dengue La dengue est une maladie virale transmise chez l'homme par les moustiques. Aucun traitement n'est actuellement disponible. Or cette infection est un énorme fardeau de santé publique : environ 3,9 milliards de personnes à risque d'infection et plus de 50 millions de cas de dengue chaque année dans le monde. Plusieurs critères ont, jusqu'à lors, empêché le développement d'un traitement efficace : (i) une administration orale; (ii) une diminution rapide de la quantité de virus dans le sang pour enrayer la progression vers la forme sévère de la dengue; (iii) une même efficacité contre les 4 principaux types de DENV (sérotypes DENV-1 à DENV-4); (iv) un profil de sécurité acceptable. Le virus de la dengue (DENV) est un virus enveloppé à ARN simple brin (polarité +, groupe IV de la classification Baltimore) appartenant au genre Flavivirus. Son génome a une longueur de 10 à 11 kb : il code pour une polyprotéine.
Source : ViralZone - Flaviviridae
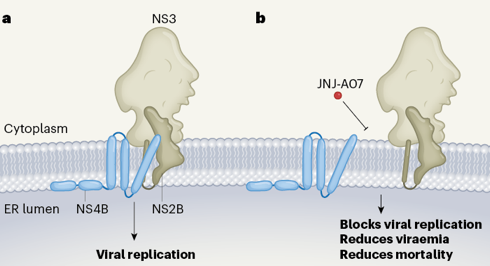
Efficacité thérapeutique prometteuse de JNJ-A07 Une molécule appelée JNJ-A07 est très efficace contre tous les variants génétiques testés des 4 sérotypes. Voir Kaptein et al. (2021).
La description de l'interaction de la molécule JNJ-A07 avec la protéine virale NS4B est un premier pas pour décrypter la façon dont cette molécule empêche la formation du complexe viral de réplication [NS3-NS4B], inhibant ainsi la réplication de DENV.
Source : Biering & Harris (2021) Figure a. La réplication du virus de la dengue nécessite la formation d'un complexe qui comprend les protéines virales [NS4B, NS3, NS5, NS2A, NS2B, NS4A et NS1] dans la membrane du réticulum endoplasmique ("ER lumen") de la cellule hôte. Figure b. La molécule JNJ-A07 semble bloquer la liaison [NS3-NS4B], empêchant la formation du complexe de réplication virale, inhibant ainsi la réplication du virus. |
d. Autres modes de défense anti-virale : exemple des "ilôts de défense" Des analyses informatiques ont montré que dans les génomes bactériens, les gènes de défense se regroupent dans des régions spécifiques appelées îlots de défense ("defence islands").
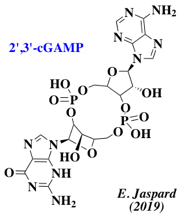
Exemple du 2',3' GMP-AMP cyclique La voie ["cyclic guanosine monophosphate (GMP) - adenosine monophosphate (AMP) synthase"/STING] ou voie [cGAS - STING] est un élément central du système immunitaire inné chez l'animal. L'enzyme cGAS (GMP - AMP cyclique synthase, EC 2.7.7.86) est un détecteur d'ADN cytosolique. Lorsqu'elle détecte un ADN viral dans le cytosol, elle synthétise une molécule signal, le di-nucléotide 2',3' GMP-AMP cyclique (2',3'-cGAMP), qui se lie à la protéine STING ("STimulator of INterferon Genes", protéine membranaire du réticulum endoplasmique) qui active la réponse immunitaire.
L'activité antivirale de cGAS a été rapportée pour divers types de virus, notamment les virus à ADN (virus de l'herpès simplex, adénovirus et virus de l'hépatite B) et les rétrovirus (HIV-1). La signalisation cGAMP fait partie d'un système de défense anti-phage commun aux bactéries (Cohen et al., 2019) :
|
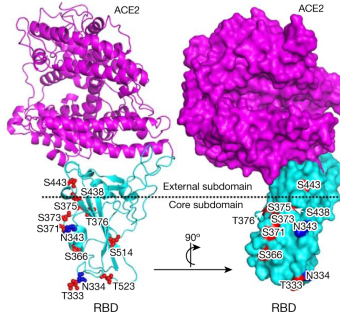
16. Mise au point de vaccins contre le coronavirus SARS-CoV-2 Le coronavirus 2 (famille des Coronaviridae) du syndrome respiratoire aigu sévère ("Severe Acute Respiratory Syndrome coronavirus 2", SARS-CoV-2) provoque la maladie respiratoire COVID-19 dont la propagation conduit à une pandémie. Il existe un très grand nombre d'approches pour la mise au point de vaccins : plus de 42 vaccins candidats sont entrés dans des essais cliniques chez l'homme et 10 sont actuellement en phase III (voir "Draft landscape of COVID-19 candidate vaccines"). Le Prix Nobel 2023 de Physiologie et Médecine a été attribué à Katalin Kariko et Drew Weissman pour leurs découvertes concernant les modifications des bases nucléosidiques qui ont permis le développement de vaccins à ARNm efficaces contre la COVID-19. a. Vaccin utilisant la glycoprotéine d'enveloppe S L'entrée du coronavirus dans la cellule hôte utilise le domaine de liaison au récepteur ("Receptor-Binding Domain", RBD) de la glycoprotéine d'enveloppe S (ou protéine de pointe S - "Spike") du SARS-CoV-2 pour interagir avec le récepteur de la cellule hôte : l'enzyme de conversion de l'angiotensine 2 (ACE2, E.C. 3.4.17.23). En effet, le site de clivage cette glycoprotéine contient plusieurs résidus arginine : elle constitue donc un excellent substrat de nombreuses protéases des cellules hôtes. Or l'ACE2 est une carboxypeptidase de la famille M2.
Source : Yang et al. (2020) Cette glycoprotéine (1273 acides aminés) est donc une cible pour la neutralisation médiée par les anticorps. Un vaccin recombinant qui correspond aux résidus d'acides aminés 319 à 545 du domaine RBD de la glycoprotéine d'enveloppe S induit une réponse anticorps fonctionnelle puissante chez les souris immunisées, les lapins et les primates non humains (Macaca mulatta) dès 7 ou 14 jours après l'injection d'une seule dose de vaccin. La glycoprotéine S est une protéine de fusion de la classe I trimèrique :
Plus de 20 vaccins contre le SARS-CoV-2 sont autorisés et plus de 90 sont en cours de développement : voir "COVID-19 vaccine tracker". |
| Vaccins utilisés dans les campagnes de vaccination mondiales contre le SARS-CoV-2 | ||||
| Nom du vaccin | Entreprise pharmaceutique | Plateforme vaccinale | Antigène | Première utilisation - lieu |
| CoronaVac | Sinovac | virus inactivé (adjuvant alum) | virion entier - WT S protein | 29/08/20 (Chine) |
| BNT162b2 (Comirnaty) | Pfizer, BioNTech | ARN messager – nanoparticules lipidiques | S protein 2P | 02/12/20 (Royaume Uni) |
| Gam-COVID-Vac (Sputnik V) | Gamaleya Research Institute | vecteurs adénovirus non-répliquants rAd26 et rAd5 | WT S protein | 05/12/20 (Russie) |
| Spikevax (mRNA-1273) | Moderna, Barda | ARN messager – nanoparticules lipidiques | S protein 2P | 18/12/20 (Etats Unis) |
| AZD1222 (Covishield) | AstraZeneca | vecteur adénovirus non-répliquant ChAdOx1 | WT S protein | 30/12/20 (Royaume Uni) |
| BBV152 (Covaxin) | Bharat Biotech International | virus inactivé (adjuvant Algel-IMDG) | virion entier - WT S protein | 02/01/21 (Inde) |
| Ad26.COV2.S | Janssen Pharmaceutical (Johnson & Johnson) | vecteur adénovirus non-répliquant rAd26 | S protein (FKO + 2P) | 17/02/21 (Afrique du sud) |
| NVX-CoV2373 | Novavax | sous-unité protéique - nanoparticules lipidiques | S protein (FKO + 2P) | En attente |
Source : Jackson et al. (2021) |
||||
| Autres types de vaccins contre le SARS-CoV-2 en cours de développement | ||
| Approche | Type de vaccin | Principe de fabrication du vaccin |
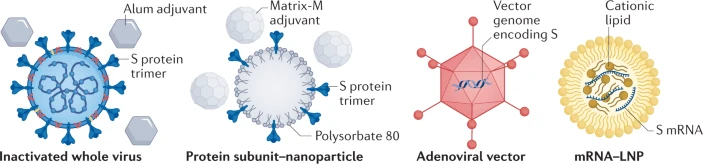
Le virus SARS-CoV-2 lui-même Au moins 7 équipes développent des vaccins en utilisant le virus lui-même (affaibli ou inactivé). Beaucoup de vaccins existant sont fabriqués de cette manière (exemples : rougeole et polio) mais ils nécessitent des tests de sécurité approfondis. |
Virus affaibli | Un virus est affaibli en le passant dans des cellules animales ou humaines jusqu'à l'apparition de mutations qui le rendent moins pathogène. |
| Virus inactivé | Le virus est rendu non infectieux en utilisant des produits chimiques (exemple, la formaldéhyde) ou la chaleur. La fabrication de ces vaccins nécessite cependant de commencer par de grandes quantités de virus infectieux. |
|
Les acides nucléiques Au moins 20 équipes essayent d'utiliser l'information ADN ou ARN pour la synthèse d'une protéine du coronavirus qui déclenche une réponse immunitaire |
La plupart de ces vaccins ciblent la glycoprotéine de pointe du virus. | L'acide nucléique (ADN ou ARN) est inséré dans les cellules humaines qui synthétisent des copies de la protéine virale. Les vaccins à base d'ARN ou d'ADN sont sûrs et faciles à produire : il faut fabriquer du matériel génétique, pas le virus. Mais ils n'ont pas encore fait leur preuve : aucun vaccin homologué n'utilise cette technologie. |
Vecteurs viraux Environ 25 équipes travaillent sur des vaccins vecteurs viraux. Ces virus sont affaiblis et ne peuvent donc pas provoquer de maladie. Il en existe 2 types: ceux qui peuvent encore se répliquer dans les cellules et ceux qui ne le peuvent pas parce que les gènes clés ont été désactivés. |
Réplication d'un vecteur viral (exemple, la rougeole affaiblie). |
Un virus tel que celui de la rougeole est génétiquement modifié afin de synthétiser des protéines de coronavirus dans l'organisme. Ces vaccins sont sûrs et provoquent une forte réponse immunitaire. L'immunité existante contre le vecteur peut diminuer l'efficacité du vaccin. |
| Vecteur viral non réplicatif (exemple, l'adénovirus). | Un virus tel que l'adénovirus est génétiquement modifié afin de synthétiser des protéines de coronavirus. Aucun vaccin homologué n'utilise cette méthode, mais il a une longue histoire en thérapie génique. Des injections de rappel peuvent être nécessaires pour induire une immunité durable. |
|
Les protéines virales De nombreuses équipes souhaitent injecter directement des protéines du coronavirus dans le corps. Des fragments de protéines ou des protéines qui imitent la couche externe du coronavirus peuvent également être utilisés. |
Sous-unités de protéines. | Vingt-huit équipes travaillent sur des vaccins visant des sous-unités de protéines virales (la plupart de ces équipes se concentrent sur la protéine de pointe du virus ou sur le domaine de liaison au récepteur- voir ci-dessus). Pour fonctionner, ces vaccins peuvent nécessiter des adjuvants (molécules immuno-stimulantes délivrées en même temps que le vaccin) et des doses multiples. |
| Particules ressemblant au virus. | Les coquilles virales vides imitent la structure externe du coronavirus, mais ne sont pas infectieuses car elles n'ont pas le matériel génétique. Cinq équipes travaillent sur de tels vaccins qui peuvent déclencher une forte réponse immunitaire mais sont difficiles à fabriquer. |
|
Source : Callaway E. (2020) Exemples de vaccins en cours de développement : CoronaVac de Sinovac, vaccin "Inactivated whole virus COVID-19" de Sinopharm, vaccin "AdV5-based vaccine" de CanSino, ChAdOx1 nCoV-19 de AstraZeneca, mRNA-1273 de Moderna, BNT162b1 & BNT162b2 de Pfizer, NVX-CoV2373 de Novavax. |
||
b. Les nanoparticules lipidiques Les membranes des cellules sont composées de différents types de glycérophospholipides amphipathiques (une partie hydrophobe et une partie hydrophile).
Source : Ondrugdelivery.com Ces processus sont utilisés pour empaqueter des agents thérapeutiques ou diagnostiques dans le liposome.
|
c. Réponses immunitaires provoquées par les vaccins à ARNm contre le SARS-CoV-2 Ces vaccins sont administrés par voie intramusculaire. Dans le vaccin, l'ARN messager (ARNm) est à l'intérieur d'une nanoparticule lipidique (voir le paragraphe ci-dessous) qui a pour fonctions :
Les nanoparticules lipidiques contenant l'ARNm ou alors l'antigène (dans le cas de la protéine de pointe S traduite et synthétisée par l'organisme vacciné) sont absorbées par les cellules présentatrices d'antigène ("Antigen-Presenting Cells" - APC). Ces APC se dirigent ensuite vers les ganglions lymphatiques où elles amorcent l'action des lymphocytes T CD4 et CD8. Les cellules APC :
Voir la figure 1 de l'article Bettini & Locci (2021). |
17. Nanomatériaux biomimétiques : thérapies antivirales et développement de vaccins Contrairement aux thérapies qui ciblent une espèce virale spécifique et peuvent perdre leur efficacité quand le virus mute, les nanomatériaux antiviraux ciblent les propriétés chimiques et physiques communes à de nombreux types de virus. Plusieurs stratégies antivirales sont en cours de développement avec des résultats prometteurs. Elle reposent sur des nanostructures notamment : (i) à base d'ADN pour piéger les virus; (ii) à base de polymères qui agissent comme des leurres en imitant la membrane de la cellule cible d'un virus ; (iii) à base de polymères qui brisent les membranes virales pour empêcher l'infection.
Source : Jackman et al. (2021) |
a. Les leurres membranaires De nombreux virus utilisent des glycoprotéines situées à leur surface pour se fixer à leur cellule hôte. Certains nanomatériaux imitant ces points d'ancrage cellulaire peuvent potentiellement agir comme des antiviraux. Exemple des "nano-éponges"
L'équipe de Zhang et collaborateurs (2020) a démontré que des nano-éponges constituées de membranes dérivées de cellules épithéliales pulmonaires humaines de type II ou de macrophages humains piégent le SARS-CoV-2 et préviennent l'infection in vitro. Les membranes de ces nano-éponges contiennent l'ACE2, récepteur du SARS-CoV-2. |
b. Les coquilles ADN pièges Les règles qui régissant la structure des capsides virales ont été décrites par Casper et Klug : le nombre de triangulation T (ou nombre de sous-unités triangulaires quasi-équivalentes distinctes) peut être calculé par l'arrangement de pentamères et d'hexamères au sein d'un icosaèdre. Ces principes permettent de construire des capsides icosaédriques synthétiques à partir d'aptamères d'ADN triangulaires auto-assemblants :
Source : Figure du dessus : Peplow M. (2021) - Figure du dessous : Sigl et al. (2021)
|
c. Exemples d'autres stratégies impliquant des nanomatériaux biomimétiques La structure de certains nanomatériaux antiviraux est optimisée pour être complémentaire de la morphologie d'un virus et ainsi mieux le piéger.
Le génome des virus est dans la capside protéique. Dans le cas des virus enveloppés (par exemple, le SARS-CoV-2), la capside est recouverte d'une membrane, ou enveloppe, essentielle à la fusion des membranes [virus - cellule cible].
|
18. Apport des méthodes d'apprentissage profond L'ensemble des mécanismes du cycle viral mettant en jeu des protéines du virus et des protéines de la cellule infectée, il est évident qu'établir un catalogue le plus exhaustif possible des interactions protéine-protéine (IPP) est un outil indispensable pour décrire les relations [cellule hôte - virus] (et bien d'autre processus, de manière plus générale) et concevoir des médicaments antiviraux. Les jeux d'apprentissage de plus en plus fournis et les méthodes d'apprentissage profond permettent de prédire un grand nombre de facteurs biologiques, notamment les IPP. Par exemple, la méthode (et l'interface web) P-HIPSTer ("Pathogen-Host Interactome Prediction using Structure similarity", prédiction d'interactome pathogène-hôte utilisant la similitude de structure) prédit les IPP [cellule hôte - virus] en tenant compte des interactions domaine - domaine et peptide - domaine. P-HIPSTer utilise 3 types de preuves pour prédire si 2 protéines interagissent.
Source : Lasso et al. (2019) Le résultat est formulé sour la forme d'un rapport de vraisemblance intégré ("integrated Likelihood Ratio", LR dans la figure ci-dessus) pour une IPP donnée :
Actuellement plus de 282.000 IPP entre plus de 12.000 protéines virales et plus de 20.000 protéines humaines sont décrites. Exemples d'autres programmes de prédiction ou d'analyse via l'apprentissage profond
Voir un cours sur l'apprentissage profond (intelligence artificielle) appliqué à la biologie. |
| 19. Liens Internet et références bibliographiques |
ViralZone
|
|
Institut Pasteur Virus-Host DB (KEGG) VICTOR : Virus Classification and Tree Building Online Resource Human Coronavirus 2019-nCoV Variants |
|
Virus Particle Explorer database : site Web qui décrit les diverses structures de la capside du virus icosaédrique (données PDB) en termes de capsides complètes. Global Initiative on Sharing All Influenza Data VirusHostPPI : A generalized approach to predicting protein-protein interactions between virus and host Accès libre à un très grand nombre d'articles scientifiques consacrés au SARS-CoV-2 (Springer Nature 2020) Prix Nobel 2023 de Physiologie et Médecine attribué à Katalin Karikó et Drew Weissman |
|
Caspar & Klug (1962) "Physical principles in the construction of regular viruses" Q. Biol. 27, 1 - 24 Baltimore D. (1971) "Expression of animal virus genomes" Bacteriol. Rev. 35, 235 - 241 Weis et al. (1988) "Structure of the influenza virus haemagglutinin complexed with its receptor, sialic acid" Nature 333, 426 - 431 Jaspard et al. (1993) "Differences in the properties and enzymatic specificities of the two active sites of angiotensin I-converting enzyme (kininase II). Studies with bradykinin and other natural peptides" J. Biol. Chem. 268, 9496 - 9503 |
|
Langlois Michel, Quintard Damien & Abalain Cécile (1994) "Synthesis of as potential inhibitors of the human immunodeficiency virus-1 protease" Eur. J. Medicin. Chem. 29, 639 - 647 Abalain Cécile & Langlois Michel (1998) "Synthesis of dibenzyl iminodiacetic derivatives as potential inhibitors of HIV-1 aspartyl protease" Eur. J. Medicin. Chem. 33, 155-160 |
|
La Scola et al. (2003) "A Giant Virus in Amoebae" Science 299, 2033 Moak & Molineux (2004) "Peptidoglycan hydrolytic activities associated with bacteriophage virions" Mol. Microbiol. 51, 1169 - 1183 Karikó, Buckstein, Ni & Weissman (2005) "Suppression of RNA Recognition by Toll-like Receptors: The impact of nucleoside modification and the evolutionary origin of RNA" Immunity 23, 165 - 175 Tsai B. (2007) "Penetration of nonenveloped viruses into the cytoplasm" Annu. Rev. Cell. Dev. Biol. 23, 23 - 43 White et al. (2008) "Structures and Mechanisms of Viral Membrane Fusion Proteins - Multiple Variations on a Common Theme" Crit. Rev. Biochem. Mol. Biol. 43, 189 - 219 Neu et al. (2008) "Structural basis of GM1 ganglioside recognition by simian virus 40" Proc. Natl. Acad. Sci. 105, 5219 - 5224 Sarafianos et al. (2009) "Structure and function of HIV-1 reverse transcriptase: molecular mechanisms of polymerization and inhibition" J. Mol. Biol. 385, 693 - 713 Tu et al. (2010) "Structural basis of HIV-1 resistance to AZT by excision" Nat. Struct. Mol. Biol. 17, 1202 - 1209 |
|
Hulo et al. (2011) "ViralZone: a knowledge resource to understand virus diversity" Nucleic Acids Res. 39, D576 - D582 Hyun et al. (2011) "Membrane Remodeling by the Double-Barrel Scaffolding Protein of Poxvirus" PLoS Pathog. 7, e1002239 Moyer & Nemerow (2011) "Viral Weapons of Membrane Destruction: Variable Modes of Membrane Penetration by Non-Enveloped Viruses" Curr. Opin. Virol. 1, 44 - 99 Johnson & Baines (2011) "Herpesviruses remodel host membranes for virus egress" Nat. Rev. Microbiol.9, 382 - 394 |
|
Kobiler et al. (2012) "Virus strategies for passing the nuclear envelope barrier" Nucleus 3, 526 - 539 Choi K.H. (2012) "Viral polymerases" Adv. Exp. Med. Biol. 726, 267 - 304 Cheng & Brooks (2013) "Viral capsid proteins are segregated in structural fold space" PLoS Comput. Biol. 9, e1002905 |
|
Fagan & Fairweather (2014) "Biogenesis and functions of bacterial S-layers" Nat. Rev. Microbiol. 12, 211 - 222 Apellaniz et al. (2014) "The three lives of viral fusion peptides" Chem. Phys. Lipids 81, 40-55 Badani et al. (2014) "Peptide entry inhibitors of enveloped viruses: the importance of interfacial hydrophobicity" Biochim. Biophys. Acta. 1838, 2180 - 2197 Bilkova et al. (2014) "Coat as a dagger: the use of capsid proteins to perforate membranes during non-enveloped DNA viruses trafficking" Viruses 6, 2899 - 2937 Jiang et al. (2014) "Picornavirus morphogenesis" Microbiol. Mol. Biol. Rev. 78, 418 - 437 |
|
Freed E.O. (2015) "HIV-1 assembly, release and maturation" Nat. Rev. Microbiol. 13, 484 - 496 Vigant et al. (2015) "Broad-spectrum antivirals against viral fusion" Nat. Rev. Microbiol. 13, 426 - 437 Perlmutter & Hagan (2015) "Mechanisms of virus assembly" Annu. Rev. Phys. Chem. 66, 217 - 239 |
|
Te Velthuis & Fodor (2016) "Influenza virus RNA polymerase: insights into the mechanisms of viral RNA synthesis" Nat. Rev. Microbiol. 14, 479 - 493 Ahi & Mittal (2016) "Components of Adenovirus Genome Packaging" Front. Microbiol. 7, 1503 Ghosh et al. (2016) "Recent Progress in the Development of HIV-1 Protease Inhibitors for the Treatment of HIV/AIDS" J. Med. Chem. 59, 5172 - 5208 De Clercq & Li (2016) "Approved Antiviral Drugs over the Past 50 Years" Clin. Microbiol. Rev. 29, 695 - 747 |
|
Latka et al. (2017) "Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process" Appl. Microbiol. Biotechnol. 101, 3103 - 3119 Erez et al. (2017) "Communication between viruses guides lysis-lysogeny decisions" Nature 541, 488 - 493 Hulo et al. (2017) "Bacterial Virus Ontology; Coordinating across Databases" Viruses 9, 126 Lusic & Siliciano (2017) "Nuclear landscape of HIV-1 infection and integration" Nat. Rev. Microbiol. 15, 69 - 82 Lin et al. (2017) "Altering the speed of a DNA packaging motor from bacteriophage T4" Nucleic Acids Res. 45, 11437 - 11448 Schoneberg et al. (2017) "Reverse-topology membrane scission by the ESCRT proteins" Nat. Rev. Mol. Cell Biol. 18, 5 - 17 |
|
Sugita et al. (2018) "Cryo-EM structure of the Ebola virus nucleoprotein-RNA complex at 3.6 angstrom resolution" Nature 563, 137 - 140 Clokie M.R.J. (2018) "Bacterial defence molecules target viral DNA" Nature 564, 199 - 200 Halldorsson et al. (2018) "Shielding and activation of a viral membrane fusion protein" Nat. Commun. 9, 349 San Martin & van Raaij (2018) "The so far farthest reaches of the double jelly roll capsid protein fold" Virol. J. 15, 181 Gizzi et al. (2018) "A naturally occurring antiviral ribonucleotide encoded by the human genome" Nature 558, 610 - 614 Doron et al. (2018) "Systematic discovery of antiphage defense systems in the microbial pangenome" Science 359, eaar4120 |
|
Krupovic et al. (2019) "Origin of viruses: primordial replicators recruiting capsids from hosts" Nat. Rev. Microbiol. 17, 449 - 458 Rolland et al. (2019) "Discovery and Further Studies on Giant Viruses at the IHU Mediterranee Infection That Modified the Perception of the Virosphere" Viruses 11, 312 Faure et al. (2019) "Serendipitous discovery in a marine invertebrate (phylum Chaetognatha) of the longest giant viruses reported till date" Virol. Curr. Res. 3, 1 |
|
Liu et al. (2019) "Cryo-EM structures of herpes simplex virus type 1 portal vertex and packaged genome" Nature 570, 257 - 261 Ahmed et al. (2019) "The regulation of Endosomal Sorting Complex Required for Transport and accessory proteins in multivesicular body sorting and enveloped viral budding - An overview" Int. J. Biol. Macromol. 127, 1 - 11 Cohen et al. (2019) "Cyclic GMP-AMP signalling protects bacteria against viral infection" Nature 574, 691 - 695 Lasso et al. (2019) "A Structure-Informed Atlas of Human-Virus Interactions" Cell 178, 1526 - 1541 |
|
Schulz et al. (2020) "Giant virus diversity and host interactions through global metagenomics" Nature 578, 432 - 436 Wu et al. (2020) "A new coronavirus associated with human respiratory disease in China" Nature 579, 265 - 269 Hillen et al. (2020) "Structure of Replicating SARS-CoV-2 Polymerase" Nature 584, 154 - 156 Benton et al. (2020) "Structural transitions in influenza haemagglutinin at membrane fusion pH" Nature 583, 150 - 153 |
|
Korber et al. (2020) "Tracking changes in SARS-CoV-2 Spike: evidence that D614G increases infectivity of the COVID-19 virus" Cell 182, 812 - 827 Al-Shayeb et al. (2020) "Clades of huge phages from across Earth's ecosystems" Nature 578, 425 - 431 Pausch et al. (2020) "CRISPR-CasΦ from huge phages is a hypercompact genome editor" Science 369, 333 - 337 |
|
Callaway E. (2020) "The race for coronavirus vaccines: a graphical guide" Nature 580, 576 - 577 Yang et al. (2020) "A vaccine targeting the RBD of the S protein of SARS-CoV-2 induces protective immunity" Nature 586, 572 - 577 Krammer F. (2020) "SARS-CoV-2 vaccines in development" Nature 586, 516 - 527 |
|
Gryseels et al. (2020) "A near-full-length HIV-1 genome from 1966 recovered from formalin-fixed paraffin-embedded tissue" PNAS 117, 12222 - 12229 Won & Lee (2020) "The current status of drug repositioning and vaccine developments for the COVID-19 pandemic" Int. J. Mol. Sci. 21, 9775 ICTV (2020) "The new scope of virus taxonomy: partitioning the virosphere into 15 hierarchical ranks" Nat. Microbiol. 5, 668 - 674 Casalino et al. (2020) "Beyond Shielding: The Roles of Glycans in the SARS-CoV-2 Spike Protein" ACS Cent. Sci. 6, 1722 - 1734 Zhang et al. (2020) "Cellular Nanosponges Inhibit SARS-CoV-2 Infectivity" Nano Lett. acs.nanolett.0c02278 |
Article |
Zhao & Woodside (2021) "Mechanical strength of RNA knot in Zika virus protects against cellular defenses" Nat. Chem. Biol. 17, 975 - 981 Scudellari M. (2021) "How the coronavirus infects cells — and why Delta is so dangerous" Nature Harvey et al. (2021) "SARS-CoV-2 variants, spike mutations and immune escape" Nat. Rev. Microbiol. 19, 409 - 424 Biering & Harris (2021) "A step towards therapeutics for dengue" Nature doi: 10.1038/d41586-021-02638-9 |
|
Kaptein et al. (2021) "A pan-serotype dengue virus inhibitor targeting the NS3–NS4B interaction" Nature doi: 10.1038/s41586-021-03990-6 Jackson et al. (2021) "Mechanisms of SARS-CoV-2 entry into cells" Nat. Rev. Molec. Cell Biol. 1 - 18 Peplow M. (2021) "Nanotechnology offers alternative ways to fight COVID-19 pandemic with antivirals" Nat. Biotechnol. 39, 1172 - 1174 Sigl et al. (2021) "Programmable icosahedral shell system for virus trapping" Nat. Mater. 20, 1281 - 128 |
|
Nguyen et al. (2022) "Sialic acid-containing glycolipids mediate binding and viral entry of SARS-CoV-2" Nat. Chem. Biol. 18, 81 - 90 Malone et al. (2022) "Structures and functions of coronavirus replicationtranscription complexes and their relevance for SARS-CoV-2 drug design" Nat. Rev. Mol. Cell Biol. 23, 21 - 39 Viana et al. (2022) "Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa" Nature doi: https://doi.org/10.1038/d41586-021-03832-5 |
|
Li et al. (2023) "Therapeutic strategies for COVID-19: progress and lessons learned" Nat. Rev. Drug Discov. 22, 449 - 475 |
Article |
![]()