| Le trafic vésiculaire - l'exocytose - les protéines SNARE |
| Tweet |
|
|
1. Introduction 2. Les protéines SNARE et leur classification 3. Mécanisme de fusion vésiculaire 4. Le complexe trans-SNARE et modification de la classification des protéines SNARE |
5. Les syntaxines, la protéine SNAP-25, les protéines SM (Sec1/Munc18) et le domaine MUN de Munc13 6. Le désassemblage du complexe SNARE : NSF et α-SNAP 7. Contrôle de la fusion vésiculaire par le calcium : les complexines et les synaptotagmines 8. Modèle de fusion des membranes 9. Liens Internet et références bibliographiques |
| 1. Introduction
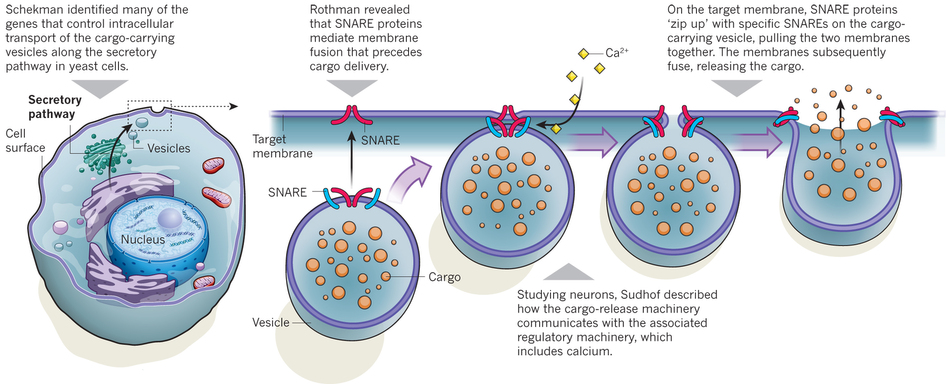
Le Prix Nobel de Physiologie et Médecine 2013 a été attribué à James Rothman, Randy Schekman et Thomas Südhof pour leurs travaux sur le trafic vésiculaire ("for their discoveries of machinery regulating vesicle traffic, a major transport system in our cells"). Le bon fonctionnement des cellules implique que les molécules nécessaires soient au bon endroit au bon moment. Certaines molécules, telles que l'insuline, doivent être exportées hors de la cellule et d'autres sont nécessaires à l'intérieur de la cellule. Les molécules synthétisées dans la cellule sont emballées dans des vésicules.
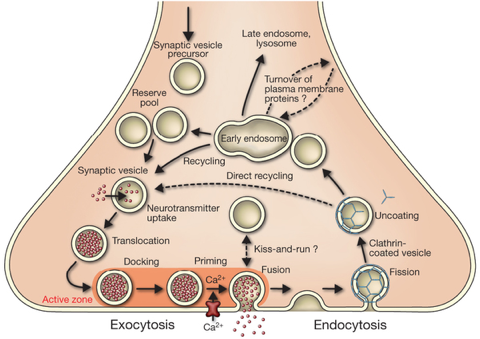
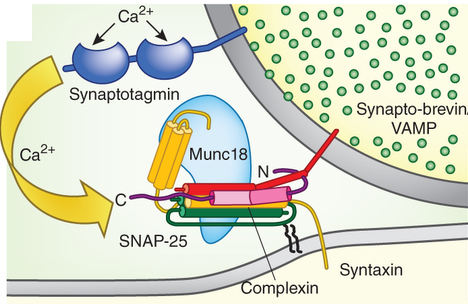
La libération de neurotransmetteurs lors du processus d'exocytose des vésicules synaptiques déclenchée par le calcium est cruciale pour la communication entre les neurones. L'exocytose et le recyclage des vésicules synaptiques contrôlent la quantité de neurotransmetteurs libérés par les terminaisons nerveuses pendant le potentiel d'action. Lors du potentiel d'action, les canaux calciques voltage-dépendants s'ouvrent et l'afflux de calcium augmente d'un facteur 105 la vitesse d'exocytose. Au repos, les vésicules synaptiques sont stockées dans le cytoplasme des terminaisons nerveuses. Certaines vésicules sont attachées à des sites spécifiques de la membrane plasmique présynaptique, appelés zones actives. Les zones actives sont composées de protéines multidomaines qui forment un échafaudage pour l'amarrage des vésicules et qui participent à l'activation de la machinerie de relarguage, processus appelé amorçage ou préparation des vésicules. Le recyclage des vésicules synaptiques semble médié par un processus d'endocytose médiée par des manteaux de clathrine.
Source : Jahn & Fasshauer (2012) Les protéines SNARE (syntaxine, SNAP-25 et synaptobrévine) sont au coeur du processus d'exocytose. D'autres protéines comme les protéines SM (Sly1, Munc18, ...), des protéines d'amorçage comme Munc13, des chaperones (HSc70, CSP et synucléines), l'AAA-ATPase NSF, la protéine adaptatrice α-SNAP ("Soluble NSF-Attachment Protein" - attention à l'acronyme : SNAP-25 et α-SNAP sont deux protéines distinctes), la complexine (action du calcium), la golgine giantine, des protéines d'attachement (p115, rab1, GM130, ...), ... participent à ce processus biologique capital. |
|
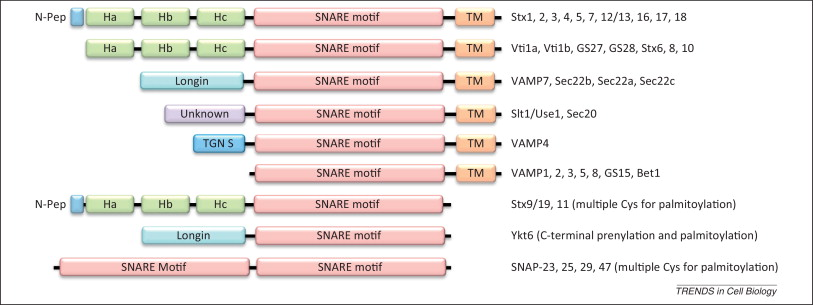
2. Les protéines SNARE et leur classification La superfamille des protéines appelée "Soluble N-ethylmaleimide-sensitive factor (NSF) Attachment protein REceptor" ou protéines SNARE est présente chez les levures et les cellules de mammifères. Elles médient la fusion entre les vésicules et la membrane de la cellule ou la membrane d'un compartiment cellulaire. Remarque : La N-éthyl-maléimide (NEM) est un composé organique dérivé de l'acide maléique qui contient une fonction imide. C'est un alcène réactif avec les fonctions thiols : il est donc utilisé pour modifier les cystéines. La NEM a été utilisée par Arthur Kornberg et al. pour comparer les activité de l'ADN polymérase I et de l'ADN polymérase III. La famille des protéines SNARE est subdivisée en 2 catégories :
Les données structurales de l'assemblage des protéines SNARE ont permis une classification plus détaillée (voir "Le complexe trans-SNARE et modification de la classification des protéines SNARE"). La plupart des protéines SNARE contiennent 1 motif SNARE : motif caractéristique d'environ 60 à 70 acides aminés contenant 8 répétitions de 7 acides aminés.
Source : Hong & Lev (2014)
|
3. Mécanisme de fusion vésiculaire L'interaction entre v-SNARE et t-SNARE conduit à la formation d'un complexe appelé trans-SNARE (ou "SNAREpin") qui déclenche la fusion des vésicules avec les membranes cibles.
Source : Wikipédia Le complexe trans-SNARE est composé de :
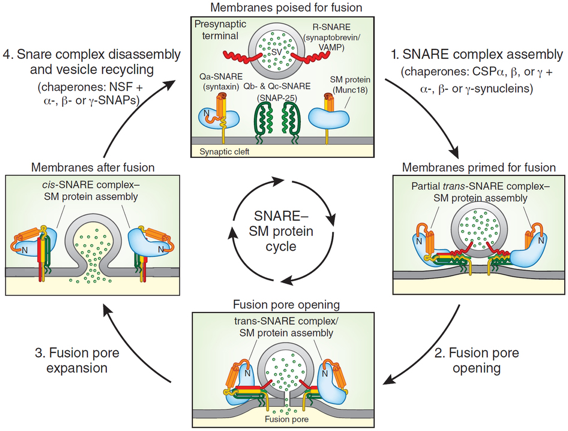
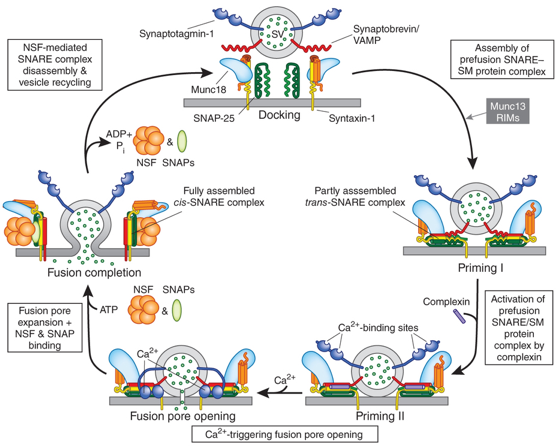
Le complexe SNARE qui demeure sur les membranes après leur fusion est appelé complexe cis-SNARE. Il est désassemblé par une AAA-ATPase (la "N-ethylmaleimide-Sensitive Fusion protein" ou "N-ethylmaleimide-sensitive factor" - NSF) et son cofacteur protéique "Soluble NSF Attachment Protein" (SNAP) pour recycler les complexes SNARE pour une nouvelle fusion. Le cycle de fusion des vésicules synaptiques
Source : Südhof T. (2013)
Dans une terminaison nerveuse, les complexes SNARE sont assemblés et dissociés en continu, avec de très nombreux processus de [repliement / dépliement] des chaînes polypeptidiques. En particulier, les intermédiaires SNARE dépliés prêts pour l'assemblage du complexe trans-SNARE sont très réactifs et sujets à un mauvais repliement et/ou à des interactions inappropriées entre les protéines de ce complexe. Deux systèmes chaperons aident au maintien des conformations correctes des protéines SNARE au cours des cycles : le système [HSc70 / CSPα ("Cysteine String Proteins") / SGT ("Small Glutamine-rich Protein")] et les synucléines (α, β et γ). |
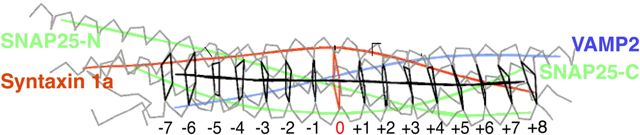
4. Le complexe trans-SNARE et modification de la classification des protéines SNARE Le complexe trans-SNARE contient les 3 protéines SNARE qui s'assemblent en un faisceau de 4 hélices parallèles tordues : (i) la syntaxine et la synaptobrevine (ou VAMP-2) forment chacune 1 hélice; (ii) SNAP-25 forme 2 hélices (appelées Sn1 et Sn2).
Source : Scales et al. (2001) Les acides aminés qui stabilisent le complexe trans-SNARE peuvent être regroupés en couches. Chaque couche contient 4 acides aminés (1 acide aminé pour chacune des 4 hélices α). Au centre du complexe se trouve la couche ionique zéro ("zero ionic layer ") composée :
Conséquences sur la classification des protéines SNARE
La plupart des protéines classées R-SNARE agissent en tant que v-SNARE et la plupart des protéines classées Q-SNARE agissent en tant que t-SNARE. Il y a des exceptions :
|
|
Visualisation du complexe SNARE de Rattus norvegicus obtenu par cristallographie à une résolution de 3,40 Å Le téléchargement de la structure peut durer plusieurs secondes.
Code PDB : 3HD7
|
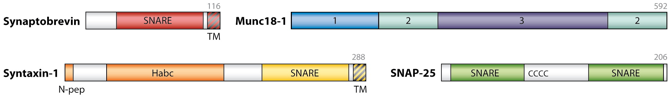
5. Les syntaxines, la protéine SNAP-25, les protéines SM (Sec1/Munc18) et le domaine MUN de Munc13
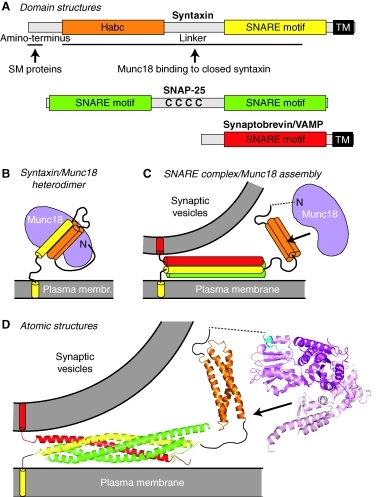
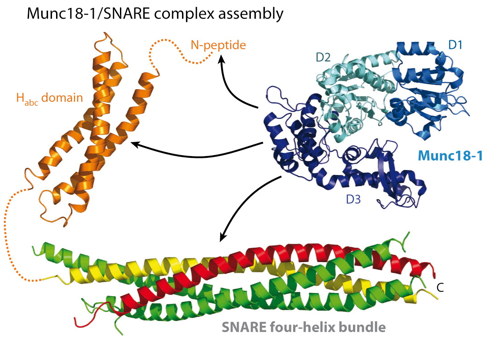
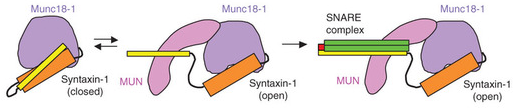
Source : Rizo & Südhof (2012) Le domaine H3 des syntaxines se fixe à la fois à la synaptobrévine et à SNAP-25 formant ainsi le noyau du complexe SNARE extrêmement stable. Cette stabilité est supposée générer l'énergie libre nécessaire pour enclencher la fusion entre la membrane de la vésicule et la membrane plasmique. Le domaine HABC est formé de 3 hélices α : quand ce domaine s'associe à l'hélice du domaine H3, la syntaxine adopte une conformation fermée inactive. Cette conformation de la syntaxine est censée être stabilisée par la fixation de Munc18 ("Mammalian UNcoordinated 18"ou Sec1-Munc18). La conformation ouverte est celle qui forme les complexes trans-SNARE. Figure ci-dessous : de nombreux points du mécanisme ne sont pas encore élucidés (ni, a fortiori, une vision unifiée du mécanisme pour tous les types d'organismes). En particulier les interactions exactes établies entre la syntaxine et les protéines SM (Munc18).
Source : Südhof & Rizo (2011) Munc18-1 possède 3 domaines (D1, D2 et D3).
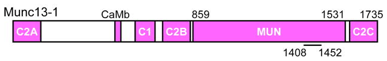
Source : Rizo & Südhof (2012) Munc18-1 semble interagir avec les 3 protéines du complexe SNARE : (i) via des interactions avec le peptide N-terminal et le domaine HABC de la syntaxine-1; (ii) via des interactions avec le faisceau à 4 hélices du complexe trans-SNARE. Les protéines Munc13 sont essentielles pour la préparation des vésicules. Les protéines Munc13 possèdent :
Source : Ma et al. (2011)
Le domaine MUN est conservé chez toutes les protéines Munc13. L'un de ses principaux rôles est d'accélérer la transition entre le complexe [syntaxine-1 en conformation fermée / Munc18-1] et le complexe [protéines SNARE / Munc18-1].
Source : Ma et al. (2011) |
6. Le désassemblage du complexe SNARE : NSF et α-SNAP L'AAA-ATPase de type II NSF ("N-ethylmaleimide-Sensitive Factor") et la protéine adaptatrice α-SNAP ("Soluble NSF Attachment Protein") sont responsables du désassemblage des complexes cis-SNARE. NSF (EC 3.6.4.6) est un homohexamère. Chaque protomère contient :
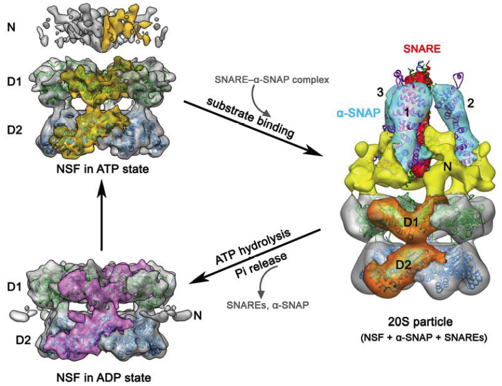
Voir un cours sur les AAA+-ATPases. Le complexe [NSF / α-SNAP / protéines SNARE] est appelé particule 20S (figure ci-dessous à droite).
Source : Chang et al. (2012) La boucle N-terminale de α-SNAP (supposée être un site d'ancrage dans la membrane) est située à l'extrémité C-terminale du complexe SNARE. Les études d'empilement ("docking") montrent que les extrémités N-terminales de 3 α-SNAP sont en contact direct avec la région C-terminale du complexe SNARE, formant ainsi une structure en forme de trépied. Les surfaces concaves électropositives des 3 α-SNAP interagissent avec la surface électronégative du complexe SNARE. Le mécanisme semble impliquer un mouvement de "haut-en-bas" nucléotide-dépendant de NSF-N au sein de l'hexamère NSF :
Par ailleurs, l'hydrolyse de l'ATP et le relarguage du groupement phosphate semblent responsables du mouvement de rotation des domaines NSF-D1 et NSF-D2 l'un par rapport à l'autre. Le couplage du mouvement de "haut-en-bas" de NSF-N après le relarguage du groupement phosphate au mouvement de rotation (anti-horaire) de NSF-D1 serait la source de la force nécessaire au désassemblage du complexe SNARE en ses 3 constituants. La détermination des structures de NSF lié à l'ATP, de NSF lié à l'ADP et de 2 complexes 20S a permis de préciser le mécanisme de désassemblage. Voir Zhao et al. (2015). |
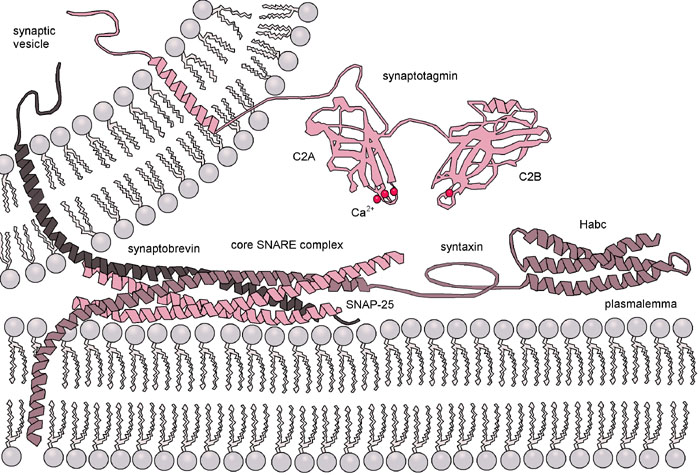
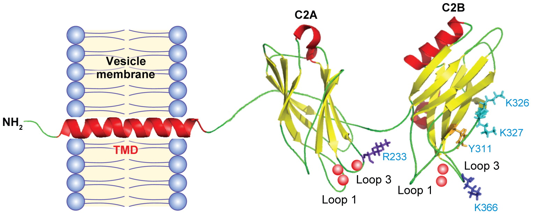
7. Contrôle de la fusion vésiculaire par le calcium : les complexines et les synaptotagmines Les complexines (appelée aussi synaphines) sont des protéines cytoplasmiques neuronales très chargées (riches en Glu et Lys) donc hydrophiles. Les synaptotagmines sont des senseurs calciques qui régulent la dynamique de fusion des pores et contrôlent les étapes finales de la fusion membranaire. Les synaptotagmines possèdent 1 domaine transmembranaire, 1 domaine intraluminal court et une grande région cytoplasmique qui contient 2 domaines C2 en tandem (C2A et C2B).
Source : Chapman E.R. (2008) Les domaines C2A et C2B sont reliés à la membrane et aussi l'un à l'autre par des "bras" flexibles (figure ci-dessus).
Les domaines C2 sont des domaines hautement conservés (140 acides aminés) qui fixent le calcium et les membranes. Les domaines C2 sont en forme de tonneaux β à 8 brins avec 2 boucles saillantes qui constituent la poche de fixation des ions calcium. Les chaînes latérales de plusieurs résidus aspartate fixent 3 ions calcium dans le domaine C2A (D172, D178, D230, D232 et D238) et 2 ions calcium dans le domaine (D303, D309, D363, D365). Mode d'action Les complexines se fixent au complexe SNARE avec une affinité élevée via leur domaine central (acides aminés 48 - 70) en formant une hélice α anti-parallèle avec la syntaxine et la synaptobrévine. Les complexines n'agissent donc que lorsque le complexe SNARE est au moins partiellement assemblé.
Source : Südhof T. (2013) Lors d'une augmentation de la concentration du calcium, la synaptotagmine déplace la complexine, ce qui permet au complexe SNARE d'arrimer la vésicule de transport à la membrane présynaptique. Les boucles de fixation du calcium des domaines C2 de la synaptotagmine (complexée au calcium) s'insèrent partiellement dans les membranes contenant des phospholipides anioniques (par exemple, des bicouches riches en phosphatidylsérine / phosphatidylcholine). Ces phospholipides complètent les sites de coordination du calcium. De plus, la synaptotagmine complexée au calcium se fixe aux protéines t-SNARE et module leur structure donc leur fonction.
Source : Südhof T. (2013) |
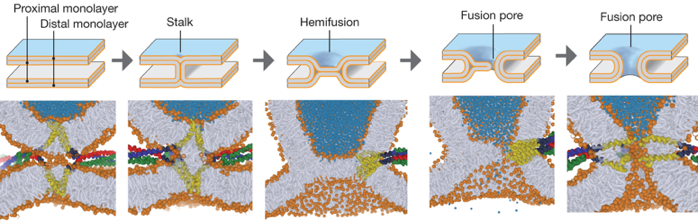
8. Modèle de fusion des membranes Dans l'étape finale de l'exocytose, la membrane de la vésicule fusionne avec la membrane plasmique. La fusion de deux bicouches implique des intermédiaires (au niveau du site de contact) qui se développent pour former un canal appelé pore de fusion. Au cours de la fusion, la barrière hydrophobe qui sépare le cytoplasme à la fois du contenu de la vésicule et de l'espace extracellulaire doit rester intacte. Figure ci-dessous : modèle de la tige ("stalk hypothesis"). Selon ce modèle, la fusion commence par un intermédiaire en forme de sablier (la tige de fusion). Il y a ensuite une semi-fusion. Puis il y a rupture ce qui entraîne la formation d'un pore de fusion.
Source : Jahn & Fasshauer (2012) |
Exemples de structures tridimensionnelles de protéines SNARE et de protéines du système d'exocytose - fusion vésiculaire |
|
| Molécule ou complexe | Code PDB |
| complexe SNARE | 1SFC - 3HD7 - 3IPD |
| Complexe [complexine / SNARE] | 1KIL |
|
Syntaxine 1A Domaine N-terminal de la Syntaxine 1A Complexe [domaine SNARE de la syntaxine / domaine N-terminal de SNAP-25] |
1EZ3 - 1S94 - 2M8R 1BR0 - 1S94 1JTH |
| Complexe [Munc18 / syntaxine 1A] | 3C98 |
|
Protéine Vps33 (Sec1/Munc18) et complexe [Vps33 / Vps16] Complexe [Sly1 / Sed5p] |
4KMO 1MQS |
| SNAP-23 | 1NHL |
| Domaine d'hexamerisation de NSF ("N-ethylmaleimide Sensitive Factor") | 1NSF |
| Domaine C2B de Munc13-1 (sans calcium) | 3KWT |
| 9. Liens Internet et références bibliographiques |
|
Articles clés de James Rothman, Randy Schekman et Thomas Südhof Novick & Schekman (1979) "Secretion and cell-surface growth are blocked in a temperature-sensitive mutant of Saccharomyces cerevisiae" PNAS 76, 1858 - 1862 Balch et al. (1984) "Reconstitution of the transport of protein between successive compartments of the Golgi measured by the coupled incorporation of N-acetylglucosamine" Cell 39, 405 - 416 Kaiser & Schekman (1990) "Distinct sets of SEC genes govern transport vesicle formation and fusion early in the secretory pathway" Cell 61, 723 - 733 Perin et al. (1990) "Phospholipid binding by a synaptic vesicle protein homologous to the regulatory region of protein kinase C" Nature 345, 260 - 263 Sollner et al. (1993) "SNAP receptor implicated in vesicle targeting and fusion" Nature 362, 318 - 324 Hata et al. (1993) "Synaptic vesicle fusion complex contains unc-18 homologue bound to syntaxin" Nature 366, 347 - 351 |
|
Jahn & Fasshauer (2012) "Molecular machines governing exocytosis of synaptic vesicles" Nature 490, 201 - 207 Südhof T. (2013) "A molecular machine for neurotransmitter release: synaptotagmin and beyond" Nature Medicine 19, 1227 - 1231 Hong & Lev (2014) "Tethering the assembly of SNARE complexes" Trends Cell. Biol. 24, 35 - 43 |
|
|
Scales et al. (2001) "The ionic layer is required for efficient dissociation of the SNARE complex by α-SNAP and NSF" PNAS 98, 14262 - 14267 Chapman E.R. (2008) "How does synaptotagmin trigger neurotransmitter release ?" Annu. Rev. Biochem. 77, 615 - 641 Südhof & Rizo (2011) "Synaptic Vesicle Exocytosis" Cold Spring Harb. Perspect. Biol. 3, a005637 Ma et al. (2011) "Munc13 mediates the transition from the closed syntaxin-Munc18 complex to the SNARE complex" Nat. Struct. Mol. Biol. 18, 542 - 549 Rizo & Südhof (2012) "The membrane fusion enigma: SNAREs, Sec1/Munc18 proteins, and their accomplices-guilty as charged ?" Annu. Rev. Cell. Dev. Biol. 28, 279 - 308 |
|
|
Rizo. J. (2010) "Synaptotagmin-SNARE coupling enlightened" Nat. Struct. Mol. Biol. 17, 260 - 262 Zhao et al. (2012) "Requirements for the catalytic cycle of the N-ethylmaleimide-Sensitive Factor (NSF)" Biochim. Biophys. Acta. 1823, 159 - 171 Chang et al. (2012) "Structural characterization of full-length NSF and 20S particles" Nat. Struct. Mol. Biol. 19, 268 - 275 Liang et al. (2013) "Prefusion structure of syntaxin-1A suggests pathway for folding into neuronal trans-SNARE complex fusion intermediate" PNAS 110, 19384 - 19389 |
|
|
Wojcik & Brose (2007) "Regulation of membrane fusion in synaptic excitation-secretion coupling: speed and accuracy matter" Neuron. 55, 11 - 24 Pang & Südhof (2010) "Cell biology of Ca2+-triggered exocytosis" Curr. Opin. Cell. Biol. 22, 496 - 505 Rizo. J. (2010) "Synaptotagmin-SNARE coupling enlightened" Nat. Struct. Mol. Biol. 17, 260 - 262 Siksou et al. (2011) "Ultrastructural organization of presynaptic terminals" Curr. Opin. Neurobiol. 21, 261 - 26 |
|
|
Risselada et al. (2012) "How SNARE molecules mediate membrane fusion: recent insights from molecular simulations" Curr. Opin. Struct. Biol. 22, 187 - 196 van der Sluijs et al. (2013) "Late steps in secretory lysosome exocytosis in cytotoxic lymphocytes" Front. Immunol. 4, 359 Zhao et al. (2015) "Mechanistic insights into the recycling machine of the SNARE complex" Nature 518, 61 - 67 |
|
![]()