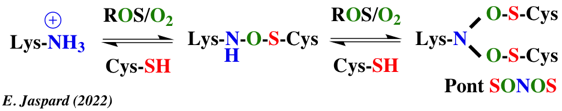| Liaison covalente lysine - cystéine : le pont NOS |
| Tweet |
|
1. Le pont NOS 2. Mécanisme supposé de formation du pont NOS |
3. La liaison NOS est un "commutateur rédox" allostérique 4. Perspectives |
|
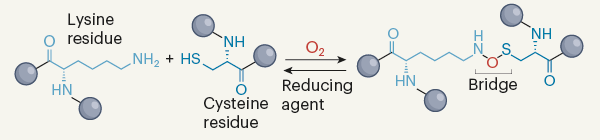
1. Le pont NOS La transaldolase (TAL, E.C. 2.2.1.2) est une enzyme clé de la voie des pentoses phosphates chez tous les organismes. Par ailleurs, Neisseria gonorrhoeae (Ng) est une bactérie pathogène responsable de la gonorrhée (maladie sexuellement transmissible). Wensien et al. (2021) ont mis en évidence un nouveau type de liaison covalente entre un résidu cystéine et un résidu lysine, appelé pont NOS, dans la transaldolase de Neisseria gonorrhoeae (NgTAL).
Source : Fass & Senenov (2021) La densité électronique à proximité immédiate de l'atome de soufre de Cys38 est compatible avec une molécule de dioxygène dissoute, ce qui suggère que l'oxydation de cette cystéine par l'oxygène moléculaire précède la formation du pont NOS. |
Visualisation du pont N(O)S dans la forme oxydée de la transaldolase de Neisseria gonorrhoeae à une résolution de 0,96 Å. Code PDB : 6ZX4
|
Le pont NOS est situé à la surface de l'enzyme et est donc accessible au solvant. Il établit de nombreuses liaisons hydrogène avec plusieurs molécules d'eau au sein d'un réseau étendu de liaisons hydrogène incluant notamment les résidus Gly36, Glu93, Thr97 et Thr101, ce qui suggère un rôle de ces résidus dans la formation du pont NOS. |
2. Mécanisme supposé de formation du pont NOS En principe, une cystéine oxydée peut réagir avec les amines mais l'atome d'azote nucléophile attaquerait préférentiellement l'atome de soufre au lieu de l'atome d'oxygène lié au soufre (formation de sulfamides ou de sulfinamides).
Les calculs de mécanique moléculaire et les valeurs de barrière d'énergie de l'état de transition théorique sont en faveur de la deuxième possibilité. |
3. La liaison NOS est un "commutateur rédox" allostérique Le pont NOS agirait, entre autre, comme un commutateur rédox permettant à l'enzyme de basculer d'un état actif à inactif et inversement ("allosteric redox switch"). Le résidu Cys38 du pont NOS (le commutateur rédox) est situé sur une structure de type "brin étendu" qui va de la surface de la protéine au site actif et qui contient notamment les résidus du site actif :
La réduction du pont NOS induit un léger ajustement conformationel de ce brin étendu qui a pour conséquence une modification de la position de Asp17, Asn43 et de certaines molécules d'eau catalytiques. Rôle biologique La formation du pont NOS chez les bactéries pathogènes pourrait être liée à la réponse au stress induit par les espèces réactives de l'oxygène puisque le stress oxydatif est un mécanisme de défense des cellules infectées par ces bactéries. La formation du pont NOS dans les TAL modifierait le flux métabolique via la voie des pentoses phosphates, modulant ainsi la formation du ribose pour la biosynthèse des acides nucléiques. |
|
Des ponts NOS ont été mis en évidence dans d'autres protéines par l'analyse des données de structure de la PDB. Pour l'instant, on peut citer :
De nombreuses protéines humaines impliquées dans des pathologies sont contrôlées par des phénomènes redox : le commutateur rédox NOS est susceptible de jouer un rôle dans la régulation de leur fonction. De plus, le pont NOS est de haute énergie d'où sa réversibilité : cette caractéristique peut être exploitée en ingénierie des peptides et des protéines pour la conception de médicaments en développant des petites molécules inhibiteurs, des anticorps et des nanocorps ciblant ce type de commutateur. Les ponts SONOS
Ces données soulignent un rôle régulateur des propriétés chimiques de ce résidu d'acide aminé dans les processus biologiques rédox. Les commutateurs [NOS / SONOS] sont fréquemment trouvés dans certaines protéines d'agents pathogènes (y compris le SARS-CoV-2) et dans des protéines chez l'homme impliquées notamment dans l'expression génique et la signalisation régulée par les processus rédox. |
| Références bibliographiques |
Wensien et al. (2021) "A lysine–cysteine redox switch with an NOS bridge regulates enzyme function" Nature 593, 460 - 464 Fass & Senenov (2021) "Previously unknown type of protein crosslink discovered" Nature 593, 343 - 344 Rabe von Pappenheim et al. (2022) "Widespread occurrence of covalent lysine-cysteine redox switches in proteins" Nat. Chem. Biol. 18, 368 - 375 |
|
![]()