| La voie des pentoses phosphates |
| Tweet |
|
|
1. Présentation générale 2. Etapes oxydatives 3. Rappel sur le NAD(P) |
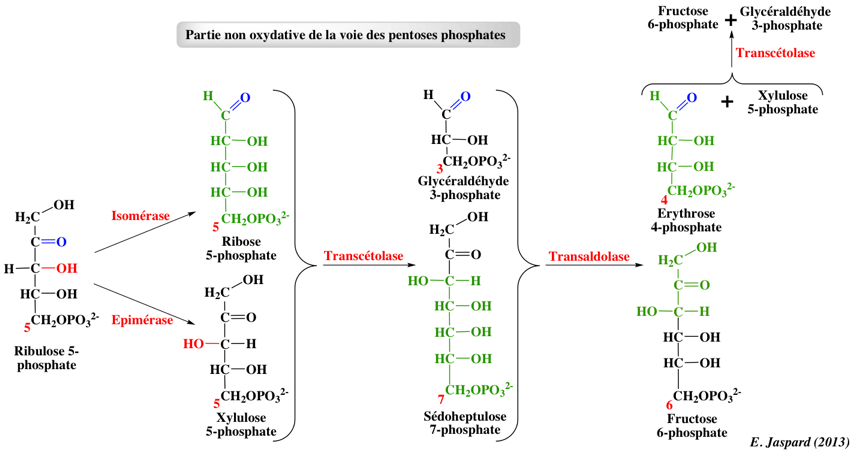
4. Etapes non oxydatives 5. Régulation de la voie des pentoses phosphates 6. Liens Internet et références bibliographiques |
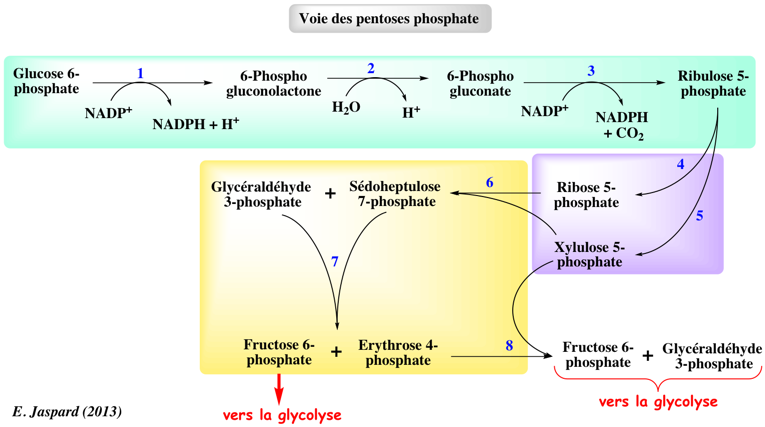
1. Présentation générale La voie des pentoses phosphates (ou voie de Otto Warburg, Frank Dickens et Bernard Horecker) est une voie du métabolisme énergétique dont les principaux rôles sont :
Cette voie :
On peut décomposer la voie des pentoses phosphates en 3 parties :
|
|
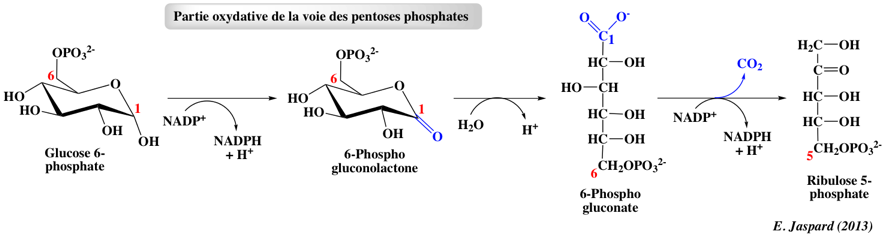
La glucose-6-phosphate déshydrogénase catalyse l'oxydation de la fonction aldéhyde (hémiacétal) portée par le carbone C1 du glucose-6-phosphate pour former un acide carboxylique dans une liaison ester, une lactone. Le NADP+ sert d'accepteur d'électrons. Cette réaction est irréversible et contrôle le flux de la voie des pentoses phosphates. Le NADPH est un inhibiteur compétitif de la glucose-6-phosphate déshydrogénase. La 6-phosphogluconolactonase catalyse l'hydrolyse de la lactone et ouvre le cycle pour former le 6-phosphogluconate. Bien que l'ouverture du cycle se produise en l'absence d'enzyme, la 6-phosphogluconolactonase accélère la réaction en diminuant la durée de vie de la 6-phosphogluconolactone très réactive et donc potentiellement toxique. La phosphogluconate déshydrogénase catalyse la décarboxylation oxydative du 6-phosphogluconate pour former le ribulose-5-phosphate (cétose à 5 carbones). L'hydroxyle en position C3 de la 6-phosphogluconate est oxydé en cétone, ce qui favorise la perte du carboxyle en C1 sous la forme de CO2. Le NADP+ sert d'accepteur d'électrons. Le ribulose 5-phosphate est aussi un intermédiaire clé du cycle de Calvin au cours de la photosynthèse. La glucose-6-phosphate déshydrogénase est régulée par la disponibilité du coenzyme NADP+. Puisque le NADPH formée est utilisé dans les voies métaboliques (réductrices) de biosynthèse, la concentration croissante du NADP+ stimule la voie des pentoses phosphates pour reconstituer le stock de NADPH. |
|
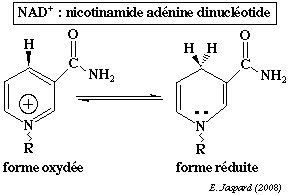
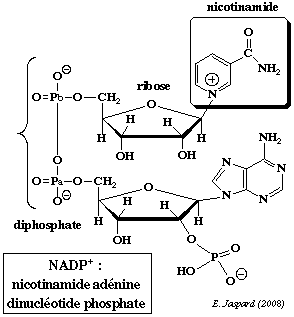
La nicotinamide adénine dinucléotide (phosphate) contient la nicotinamide qui est l'amide de l'acide nicotinique.
A cours de la réduction du coenzyme, le groupe nicotinamide capte un proton et un ion hydrure H+ d'un substrat qui est déshydrogéné : NAD(P)+ + 2H+ +2 e- <=> NAD(P)H + H+ Dans la structure de la nicotinamide adénine dinucléotide phosphate (réduite ou non / NADP+ ou NADPH+), un groupe phosphoryle supplémentaire substitue l'hydrogène de l'hydroxyle situé en position 2' du ribose lié à l'adénine.
|
|
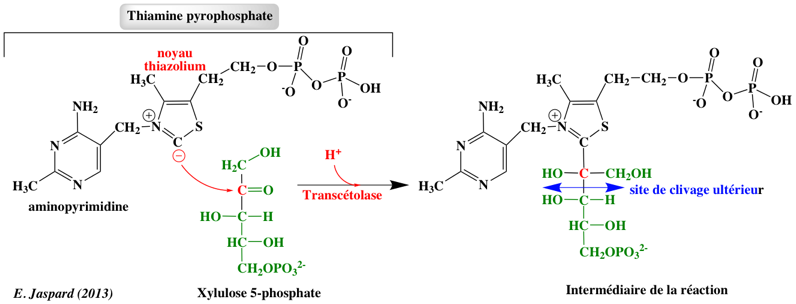
5. Régulation de la voie des pentoses phosphates En fonction des besoins de la cellule en ribose 5-phosphate, en NADPH et en ATP, la voie des pentoses phosphates peut fonctionner selon différents modes afin d'optimiser la concentration de ces métabolites : 1. Une quantité plus importante de ribose 5-phosphate que de NADPH est nécessaire. Exemple: les cellules en division rapide ont de forts besoins en ribose 5-phosphate pour la synthèse d'ADN. La transcétolase et la transaldolase forment le ribose 5-phosphate à partir du fructose 6-phosphate et du glycéraldehyde 3-phosphate : ces réactions fonctionnent alors dans le sens opposé. 2. Une quantité plus importante de NADPH que de ribose 5-phosphate est nécessaire : toute la voie des pentoses phosphates est utilisée. Exemple: les cellules des tissu adipeux ont besoin de beaucoup de NADPH pour la synthèse des acides gras. Il y a alors deux cas de figure :
3. Les besoins en ribose 5-phosphate et NADPH sont équilibrés : la partie oxydative de la voie des pentoses phosphates est utilisée. Le ribose 1-phosphate généré lors du catabolisme des nucléosides entre également de cette manière dans la glycolyse, après avoir été converti en ribose 5-phosphate. La voie des pentoses phosphates sert donc de point d'entrée dans la glycolyse pour certains oses à 5 ou 6 carbones. Une régénération efficace du NADPH est l'un des facteurs limitant dans la production par des processus de bio-transformation. Des souches d'Escherichia coli ont ainsi été manipulées par ingéniérie :
Ces modifications ont des effets remarquables sur les bioprocédés NADPH dépendants. |
| 6. Liens Internet et références bibliographiques |
|
"Pentose phosphate pathway" Horecker et al. (1951) "The enzymatic conversion of 6-phosphogluconate to ribulose-5-phosphate and ribose-5-phosphate" J. Biol. Chem. 193, 383 - 396 Lee et al. (2013) "Engineering of NADPH regenerators in Escherichia coli for enhanced biotransformation" Appl. Microbiol. Biotechnol. 97, 2761 - 2772 |
![]()