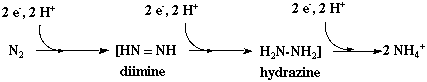| Assimilation de l'azote en ammoniac - Principales voies de la synthèse des acides aminés Catabolisme de la partie azotée des acides aminés : le cycle de l'urée |
| Tweet |
|
|
A. ASSIMILATION DE L'AZOTE ATMOSPHERIQUE 1. Introduction 2. Assimilation de l'azote par le système nitrogénase 3. Assimilation de l'azote par la nitrate réductase et la nitrite réductase 4. Rôle de la ferrédoxine B. PRINCIPALES VOIES DE LA SYNTHESE DES ACIDES AMINES 1. Introduction 2. 1ère incorporation d'ammoniac : formation du glutamate par la glutamate déshydrogénase (EC 1.4.1.4) 3. Rôle prépondérant du glutamate dans les réactions de transamination 4. 2ème incorporation d'ammoniac : formation de la glutamine à partir du glutamate par la glutamine synthétase 5. La proline est synthétisée à partir du glutamate 6. Le glutathion : cofacteur issu du glutamate |
7. Synthèse de la glycine à partir de la sérine : rôle du tétrahydrofolate 8. Synthèse de la cystéine à partir de la sérine ou à partir de la méthionine : rôle de la S-adénosylméthionine 9. Aperçu de la synthèse des acides aminés aromatiques C. CATABOLISME DE LA PARTIE AZOTEE DES ACIDES AMINES : LE CYCLE DE L'UREE 1. Aperçu du devenir de la chaîne carbonée 2. Formes d'excrétion des déchets azotés 3. Présentation générale de l'ammoniogénèse et du transport de l'ammoniac 3a. La désamination directe 3b. La désamidation 3c. L'ammoniac 4. Le cycle de l'urée 5. Bilan de la synthèse de l'urée et conclusion |
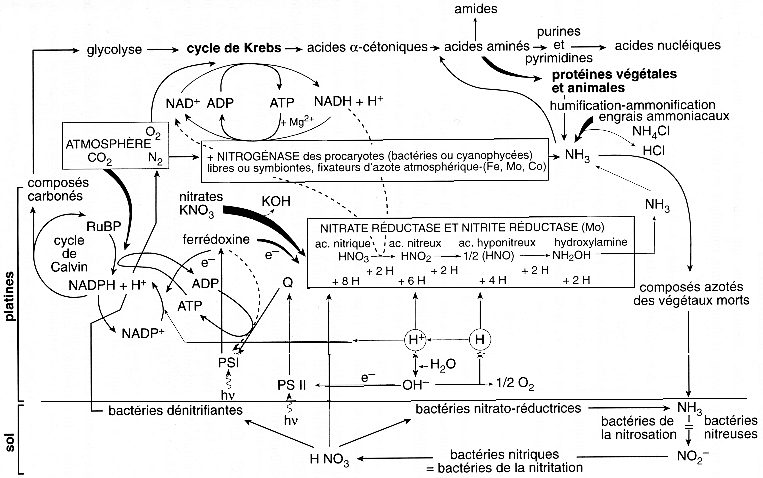
A. Assimilation de l'azote atmosphérique 1. Introduction - Voir le schéma général a. Les acides aminés sont tous constitués :
b. Les protéines sont continuellement synthétisées et dégradées à une très grande vitesse. Cette dégradation aboutit aux acides aminés, mais ces acides aminés ne sont pas ou peu ré-utilisés pour synthétiser de nouvelles protéines. c. Le produit de dégradation des groupements aminés des protéines est une molécule capitale dans l'élimination de l'ammoniac, initialement introduit dans les acides aminés : il s'agit de l'urée, fabriquée au cours d'un cycle qui porte son nom. d. Enfin, les excréments de toutes sortes et les cadavres animaux et végétaux sont décomposés par des bactéries dites dénitrifiantes, qui vont reformer de l'azote gazeux que l'on retrouve dans l'atmosphère. L'azote N2 représente 80% des gaz de l'atmosphère. C'est un gaz extrémement stable dont l'énergie libre de la triple liaison entre les atomes d'azote est de 225 kcal.mol-1, ce qui explique le coût élevé de la fabrication d'engrais chimique du fait que le procédé utilisé qui est la réduction directe de N2 par H2, nécessite beaucoup d'énergie : La réaction est : N2 + 3 H2 <=> 2 NH3 ΔG' = + 147 kcal.mol-1 Dans l'industrie, les conditions sont : température = 500°C / pression = 300 atm / fer = catalyseur |
|
2. Assimilation de l'azote par le système nitrogénase La réduction de l'azote gazeux en ammoniac s'appelle la fixation de l'azote. Certaines plantes (notamment les légumineuses) vivent en symbiose avec des micro-organismes procaryotes, qui sont seuls capables de fixer l'azote atmosphérique. Ces micro-organismes, par exemple les bactéries Rhizobium, possèdent un ensemble de gènes qui codent pour un système que l'on appelle le système nitrogénase. L'assimilation de l'azote atmosphérique par les organismes s'effectue donc par ce système.
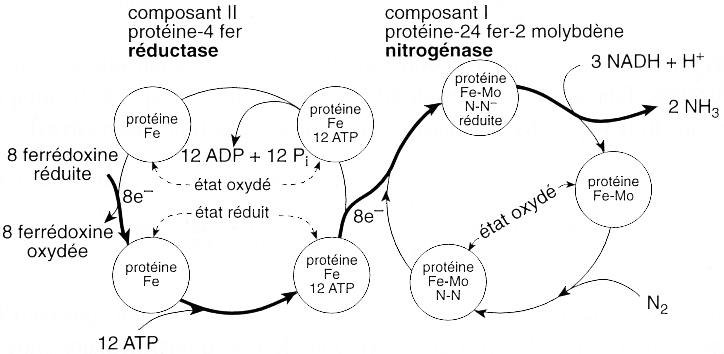
Source : "Physiologie végétale I - Nutrition et métabolisme" D. Laval-Martin & P. Mazliak (1995) - Ed. Hermann, coll. Méthodes Le système nitrogénase est un assemblage de 3 protéines : 1. la nitrogénase proprement dite qui est un tétramère de 220 kDa (2 x 51 kDa et 2 x 59 kDa) :
2. une réductase qui est un dimère de 60 kDa (2 x 30 kDa), le dimère contenant un centre [4 Fe - 4 S] 3. la ferrédoxine chez les bactéries symbiotiques ou une flavodoxine à FAD chez les bactéries libres. C'est la protéine transporteuse des électrons du fer vers le molybdène. Ci-dessous : mécanisme supposé de la réduction de l'azote en ammoniac par le système nitrogénase.
Le mécanisme de transfert des électrons (figure ci-dessous) entre les protéines du système nitrogénase met en jeu des états réduits et oxydés de ces protéines.
Source : "Physiologie végétale I" - Laval-Martin & Mazliak (1995) La réaction globale , irréversible, catalysée par le système nitrogénase, est la suivante : N2 + (6 H+ + 6 e-) + 6 ferrédoxine réduite + (2 H+ + 2 e-) + 2 ferrédoxine réduite + 12 ATP + 12 H2O ---> 2 NH3 + H2 + 8 ferrédoxine oxydée + 12 ADP + 12 Pi On constate que cette réaction est très coûteuse en énergie puisque 12 molécules d'ATP sont consommées par molécule d'azote réduit. Le rôle de l'ATP est le suivant :
|
|
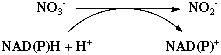
3. Assimilation de l'azote par la nitrate réductase et la nitrite réductase L'autre mode d'assimilation de l'azote est indirect puisque en premier lieu, les décharges à haut potentiel électrique de la foudre catalysent l'oxydation de N2 par l'oxygène atmosphérique et l'on aboutit à la formation d'oxyde d'azote : les ions nitrate NO3- ou nitrite NO2-. Ces ions sont ensuite ramenés au sol par les pluies. Puis deux enzymes interviennent : - une nitrate réductase qui catalyse la réaction :
- une nitrite réductase qui catalyse la réduction du nitrite en ammoniac selon le mécanisme supposé :
|
|
La ferrédoxine intervient dans les deux modes d'assimilation précédents comme donneur - accepteur d'électrons :
|
|
Visualisation de la ferrédoxine [2Fe - 2S] de Aphanothece sacrum à une résolution de 1,46 Å (2012). PDB : 3AV8 Une seule des 4 chaînes de l'homotétramère est représentée. La réaction de transfert des électrons des deux atomes de fer des centres [Fe - S] est : Fe3+ + e- ---> Fe2+
|
|
Comment l'ammoniac est-il relargué des organismes ? Si l'on revient au schéma général, le relarguage d'azote gazeux est effectué par des bactéries qui transforment l'ammoniac (issus des produits d'excrétion et des cadavres) en nitrite puis celui-ci en nitrate : c'est ce que l'on appelle la nitrification. Enfin, d'autres bactéries réduisent le nitrate en azote gazeux, c'est la dénitrification et le cycle de l'azote est fini. La conclusion de ce chapitre sur l'origine de l'azote dans les biomolécules est que la plupart des plantes dépendent de l'azote qui a déjà été fixé. a. Les plantes prélèvent donc l'azote dans leur environnement soit :
Cet aspect souligne la dépendance des animaux, et en particulier de l'homme, à leur environnement. |
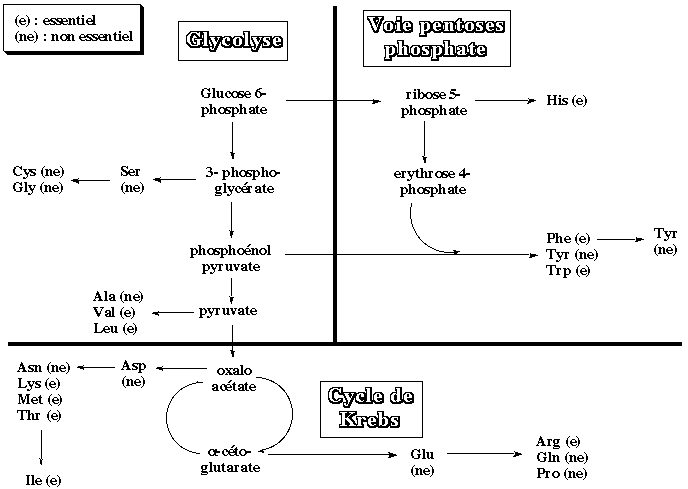
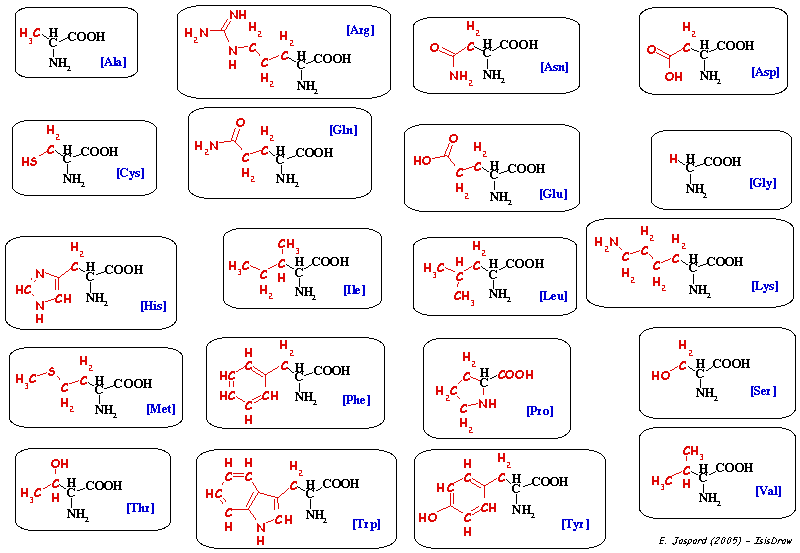
B. Principales voies de la synthèse des acides aminés Les acides aminés sont au nombre de 20 et l'on pourrait s'attendre à un nombre extrèmement grand de voies de biosynthèse de ces composés :
Voir un développement sur les acides aminés La plupart des chimiotrophes, et les mammifères en particulier, ne peuvent synthétiser que 10 des 20 acides aminés : ces acides aminés sont dits non-essentiels (ne - figure ci-dessous). Pour les 10 autres acides aminés, soit ils ne possèdent pas les enzymes qui les catalysent, soit ceux-ci sont synthétisés en quantité trop minime pour le bon fonctionnement de l'organisme. Ces 10 acides aminés sont dits essentiels (e - figure ci-dessous) et doivent être apportés par l'alimentation.
A l'inverse, les plantes et un bon nombre de microorganismes sont eux capables de synthétiser les 20 acides aminés en utilisant, comme on l'a vu, les oxydes d'azote ou l'ammoniac comme source d'azote. Enfin, contrairement aux sucres at aux acides gras dont la finalité est essentiellement de servir de combustible à la cellule pour fournir de l'énergie, le but premier des acides aminés est la synthèse de protéines. On peut même souligner que la synthèse des acides aminés nécessite beaucoup d'ATP, comme le résume le tableau suivant. |
| acides aminés non essentiels | moles d'ATP consommées par mole d'acide aminé formé | acides aminés essentiels | moles d'ATP consommées par mole d'acide aminé formé |
| glycine | 12 | thréonine | 31 |
| sérine | 18 | valine | 39 |
| cystéine | 19 | histidine | 42 |
| alanine | 20 | arginine | 44 |
| aspartate | 21 | méthionine | 44 |
| asparagine | 22 | leucine | 47 |
| glutamate | 30 | lysine | 50 ou 51 |
| glutamine | 31 | isoleucine | 55 |
| proline | 39 | phénylalanine | 65 |
| tyrosine | 62 | tryptophane | 78 |
Ces chiffres incluent l'ATP consommé pour la synthèse des précurseurs et leur conversion en produit. Source : Atkinson (1977) "Cellular energy metazbolism and its regulation" NY Acadamic Press |
|||
| De ce tableau, on ne peut rien conclure de particulier, si ce n'est que la synthèse de la glycine qui est l'acide aminé le plus simple consomme le moins d'énergie, et qu'à l'inverse la synthèse des acides aminés aromatiques qui ont des structures cycliques complexes correspondent aux voies les plus gourmandes en ATP. |
|
2. 1ère incorporation d'ammoniac : formation du glutamate par la glutamate DH La première étape de la synthèse des acides aminés est l'assimilation de NH4+. L'une des principales portes d'entrée utilisée par la cellule pour introduire l'ammoniac au coeur du métabolisme des acides aminés est l'amination réductrice de l'a-cétoglutarate en glutamate catalysée par la glutamate déshydrogénase :
|
|
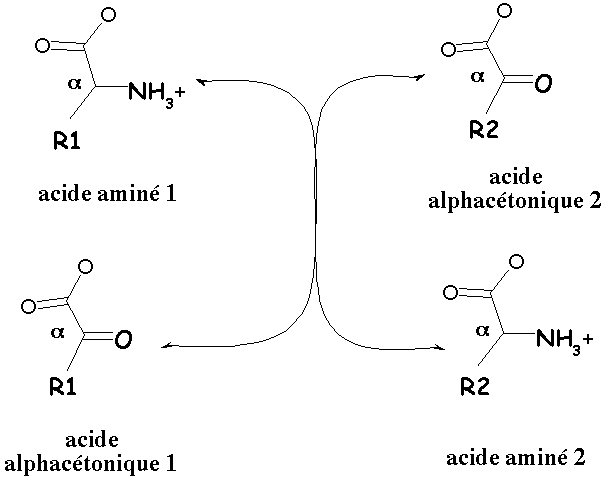
3. Rôle prépondérant du glutamate dans les réactions de transamination Le point de départ est la synthèse du glutamate car :
Le coenzyme de toutes les transaminases est le phosphate de pyridoxal qui forme une base de Schiff avec une lysine du site actif. Ces réactions de transamination permettent donc de fabriquer un grand nombre d'acide aminés courants. Cependant, dans le cas :
|
|
4. 2ème incorporation d'ammoniac : formation de la glutamine à partir du glutamate par la glutamine synthétase Une seconde réaction clé dans l'assimilation d'ammoniac correspond à la formation de glutamine à partir de glutamate et d'ammoniac, réaction catalysée par la glutamine synthétase. Cette réaction illustre le couplage de l'hydrolyse de l'ATP à la biosynthèse d'une molécule :
La glutamine synthétase de Escherichia Coli est un excellent exemple pour souligner la régulation à laquelle sont soumises la plupart des enzymes qui interviennent dans la synthèse des acides aminés. En effet cette enzyme est un oligomère de 12 sous-unités identiques qui porte chacune non seulement un site actif mais également des sites de fixation d'inhibiteurs allostériques :
|
|
5. La proline est synthétisée à partir du glutamate La synthèse de la proline est une voie qui tout en n'étant pas la plus complexe souligne le nombre important d'étapes que peut nécessiter la synthèse de certains acides aminés, à fortiori si ceux-ci sont cycliques :
|
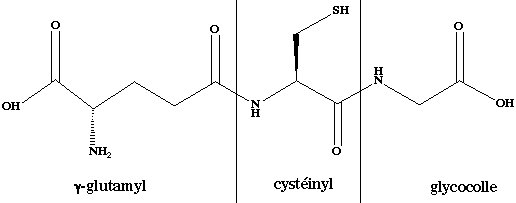
6. Le glutathion : cofacteur issu du glutamate a. Le glutathion est un tripeptide formé de glycine, de cystéine et de glutamate.
Cependant, il a une particularité : c'est le groupement carboxylique en position γ de la chaîne latérale du glutamate qui établit la liaison peptidique avec la cystéine. C'est donc le γ-glutamyl-cystéinyl-glycine. b. Le glutathion existe sous forme réduite ou oxydée. La forme oxydée correspond à l'association de 2 molécules de glutathion reliées par un pont disulfure (G-S-S-G). L'enzyme qui catalyse cette réaction est la glutathion réductase : G-S-S-G + NADPH + H+ <=> 2 G-SH + NADP+ c. L'une de ses fonctions biologiques est l'élimination de produits dérivés de l'oxygène qui sont hautement réactionnels et donc toxiques pour la cellule :
|
|
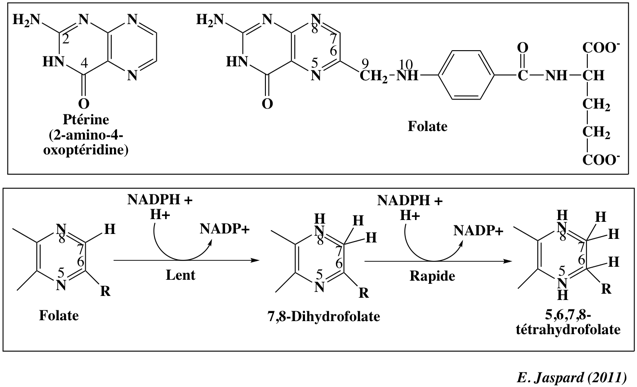
7. Synthèse de la glycine à partir de la sérine : rôle du tétrahydrofolate La glycine est synthétisée à partir du 3-phosphoglycérate, lui-même issu de la voie de la glycolyse. Le groupe hydroxyle est transformé en cétone pour donner le 3-phospho-hydroxypyruvate qui à son tour subit une transamination avec le glutamate pour former la 3-phosphosérine. Enfin la sérine est formée par la 3-phosphosérine phosphatase. La sérine est le précurseur direct de la glycine : celle-ci est formée via une réaction réversible catalysée par la sérine hydroxy-méthyltransférase. Cette enzyme ne fonctionne qu'en présence de deux cofacteurs : le phosphate de pyridoxal et un co-substrat, le tétrahydrofolate.
Le folate est composé de la ptérine, d'un résidu acide p-aminobenzoïque et d'un résidu glutamate (à droite dans la structure). Le tétrahydrofolate dérive du folate par addition d'atomes d'hydrogène en position 5, 6, 7 et 8 de la ptérine. On peut noter que la glycine est à l'origine de la chlorophylle et de l'hème. |
|
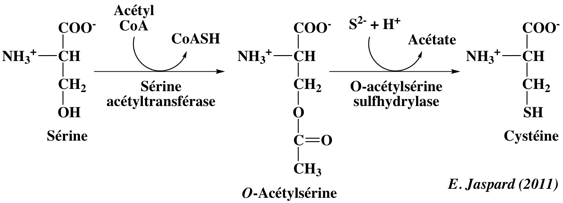
8. Synthèse de la cystéine à partir de la sérine ou à partir de la méthionine : rôle de la S-adénosylméthionine La sérine est le précurseur de la cystéine. Chez les plantes et les bactéries, la cystéine est formée en deux étapes :
Chez les autres organismes, la cystéine est synthétisée à partie d'un dérivé de la méthionine extrèmement important : la S-adénosylméthionine. La réaction est la suivante :
|
| 9.
Aperçu de la synthèse des acides aminés aromatiques
La synthèse des autres acides aminés, notamment celle des acides aminés aromatiques, est extrèmement complexe, entre autre du fait qu'à partir d'un squelette carboné linéaire, il y a cyclisation ou bien ramification. La première étape de la synthèse de ces acides aminés fait intervenir :
|
C. Catabolisme de la partie azotée des acides aminés : le cycle de l'urée
|
|
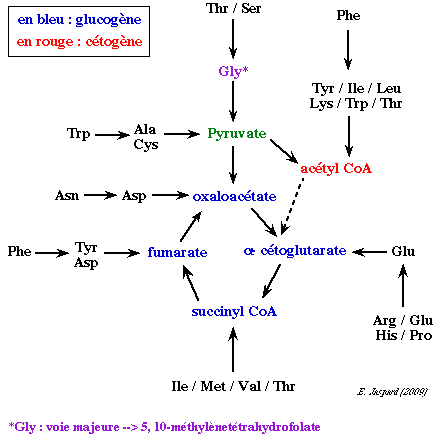
1. Aperçu du devenir de la chaîne carbonée Les caractéristiques principales de la dégradation du squelette carboné sont les suivantes :
a. Cette dégradation est un exemple remarquable d'économie des transformations métaboliques. En effet, le catabolisme de l'ensemble des squelettes carbonés des 20 acides aminés est canalisé en seulement 7 molécules :
b. Les acides aminés dégradés en acétyl CoA, en acéto-acétyl CoA ou en d'autres dérivés du coenzyme A, sont dits cétogènes puisqu'ils contribuent à former des corps cétoniques. Ceux qui aboutissent au pyruvate ou à un intermédiaire du cycle de Krebs sont dits glucogènes. En effet, ils peuvent être convertis en phosphoénolpyruvate et alimenter la voie de la néoglucogénèse. Cette classification n'est pas absolue puisqu'elle dépend du devenir réel d'un métabolite considéré. c. Quelques exemples caractéristiques du catabolisme du squelette carboné :
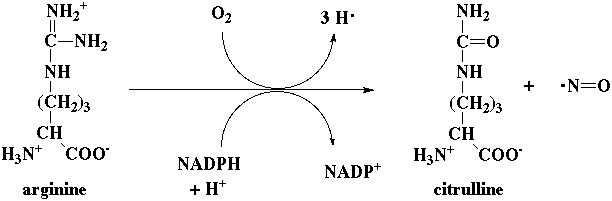
d. Le monoxyde d'azote Les mamifères possèdent une enzyme cytosolique, la NO synthase qui catalyse la formation de NO à partir de l'arginine :
|
|
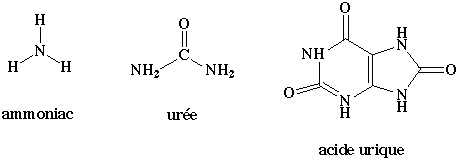
2. Formes d'excrétion des déchets azotés a. Du fait de la petite taille et de la grande solubilité de l'ammoniac, les poissons et beaucoup d'animaux aquatiques éliminent l'azote aminé sous cette forme, par diffusion au travers des membranes. Ils sont dits ammoniotéliques.
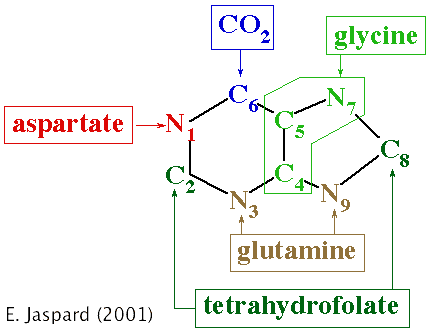
b. L'excrétion d'ammoniac pour un animal terrestre, donc ne baignant pas dans un grand volume d'eau, est extrèmement difficile car il lui faudrait uriner énormément pour arriver à diluer suffisamment l'ammoniac. En conséquence, l'homme et les mammifères éliminent l'azote sous forme d'urée, dont la toxicité est environ 100.000 fois plus faible que celle de l'ammoniac. Ils sont dits uréotéliques. L'urée est synthétisée dans le foie et le système circulatoire transporte l'urée jusqu'aux reins. Cependant, toute l'urée n'est pas éliminée par les reins : une partie, en effet, y est retenue où elle contribue à la régulation de l'osmolarité qui joue un rôle extrèmement important dans la réabsorption d'eau. c. Les oiseaux et les reptiles terrestres éliminent l'azote sous forme d'acide urique. Ils sont dits uricotéliques. L'acide urique est des milliers de fois moins soluble dans l'eau que ne le sont l'ammoniac et l'urée. Aussi, l'acide urique est-il éliminé sous forme de pâte (par exemple la fiente d'oiseau) après réabsorption totale de l'eau de l'urine. d. Des expériences utilisant des précurseurs marqués par des isotopes radioactifs ont permis de déterminer l'origine des atomes de carbone et d'azote constitutifs de l'acide urique. Ces expériences ont montré que l'acide urique et les purines qui entrent dans la composition des acides nucléiques sont issus des mêmes précurseurs et de la même voie métabolique.
|
|
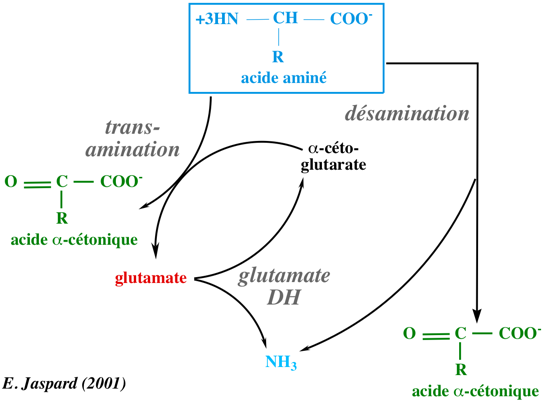
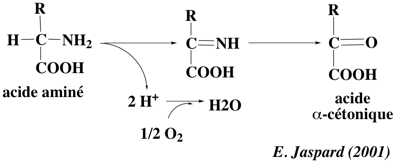
Entre 1932 et 1935, Hans Krebs et Kurt Henseleit observèrent qu'en incubant des acides aminés en présence d'un homogénat de foie et d'oxygène, ils obtenaient de l'ammoniac avec consommation de l'oxygène et disparition des acides aminés. Cette réaction est une désamiNation oxydative puisqu'il y a perte d'un groupement aminé accompagné d'un processus oxydatif. Ces réactions sont catalysées par des amino-acides oxydases.
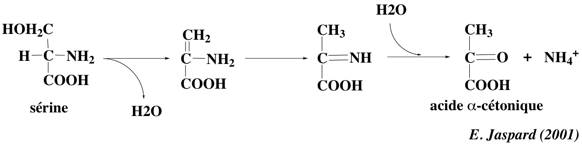
Il existe 2 cas particuliers : il s'agit des 2 acides aminés à fonction alcool (Ser et Thr), pour lesquels la perte du groupement aminé se fait par désamiNation non oxydative, catalysée par la sérine déshydratase et la thréonine déshydratase.
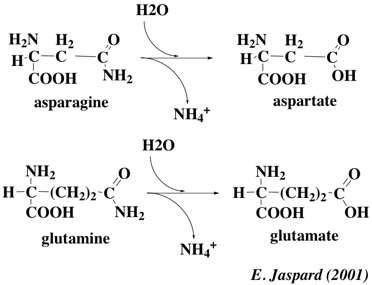
L'asparagine et la glutamine contiennent une fonction amide portée par leur chaîne latérale. Il existe 2 enzymes trés répandues, l'asparaginase et la glutaminase qui catalysent la réaction de désamiDation repectivement de l'asparagine et de la glutamine.
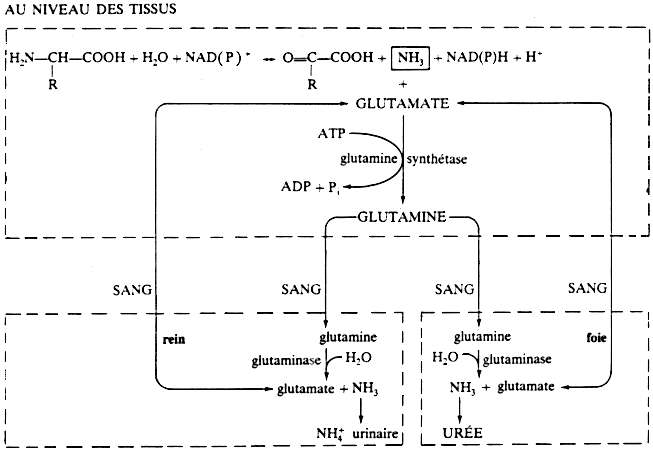
L'ammoniac est une molécule extrémement toxique pour la cellule.
En conséquence, quel que soit le mécanisme par lequel l'ammoniac est libéré (désamination directe, non oxydative et/ou désamidation), celui-ci se condense avec le glutamate pour former la glutamine (réaction catalysée par la glutamine synthétase). Puis la glutamine, l'acide aminé le plus concentré dans le sang (450 à 600 µM), sert de transporteur de l'ammoniac jusqu'au foie ou jusqu'aux reins. Dans chacun de ces organes, la glutaminase libére l'ammoniac de la glutamine par désamiDation. Au niveau des reins : l'ammoniac est éliminé dans l'urine sous forme d'ions ammonium. Cette élimination est d'autant plus importante qu'elle permet non seulement d'éliminer l'ammoniac mais aussi une grande quantité d'ions H+ formés au cours de diverses réactions métaboliques. Dans le foie : l'ammoniac va être transformé en urée : c'est ce que l'on appelle l'uréogénèse ou cycle de l'urée. L'urée est ensuite véhiculée par la circulation jusqu'aux reins d'où elle est éliminée par l'urine. |
a. formation de carbamylphosphate
5. Bilan de la synthèse de l'urée et conclusion CO2 + NH4+ + 3 ATP + Asp + 2 H2O ---> UREE + 2 ADP + 2 Pi + AMP + PPi + fumarate
|
![]()