| Les différents types de récepteurs - Les récepteurs couplés aux protéines G (RCPG) et les protéines G |
| Tweet |
|
|
1. Introduction. 2. Les principaux types de récepteurs
|
3. Les récepteurs couplés aux protéines G (récepteurs membranaires métabotropes)
4. Les protéines G 5. Exemples de transduction de signaux par des récepteurs couplés aux protéines G 6. Le récepteur de l'insuline 7. Liens Internet et références bibliographiques |
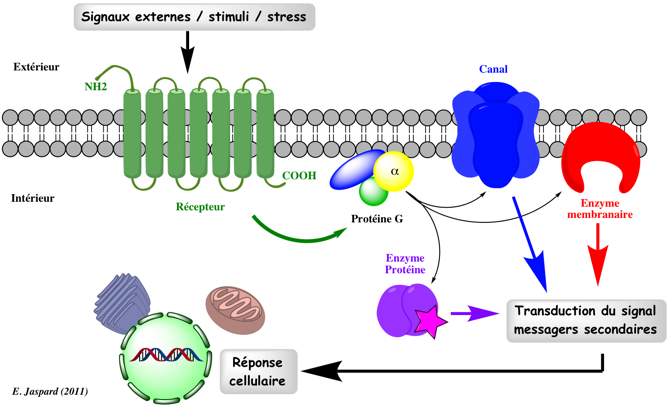
1. Introduction Toutes les cellules ont mis au point des mécanismes qui leur permettent de percevoir des signaux internes ou provenant de leur environnement et d'y apporter une réponse appropriée. Si les signaux sont extracellulaires, il existe des systèmes de transduction des signaux qui modifie leur nature de sorte que leur sens biologique soit "compris" à l'intérieur de la cellule. Cela implique l'existence de protéines particulières au niveau des membranes : les récepteurs. Pour survivre, les organismes unicellulaires doivent trouver et ingérer leur nourriture tout en évitant les substances toxiques. Les organismes supérieurs doivent répondre à une série de signaux encore plus importante et définir des systèmes de réponse pour les éléments nutritfs, les facteurs de croissance, les neurotransmetteurs, les stimuli sensoriels. Exemples de stimuli capables d'activer un récepteur couplé aux protéines G - RCPG (Source : J.M. Botto)
Dans tous ces cas :
La transduction des signaux se traduit par une série ou cascade de modifications des structures des molécules impliquées dans cette signalisation. Ces évènements ont une incidence sur d'autres processus biologiques ou voies métaboliques. La transduction du signal est extrêmement régulée. |
|
Exemple d'une cascade d'évènements L'activation des récepteurs couplés aux protéines G correspond à une cascade d'événements (figure ci-dessous) tels que : (i) l'activation du récepteur par la fixation de son ligand; (ii) l'activation d'une protéine G par hydrolyse du GTP; (iii) l'activation ou l'inhibition d'une protéine effectrice par une sous-unité de la protéine G.
Cette cascade d'événements inclut trés souvent la modulation de la concentration intracellulaire de molécules intermédiaires appelées messagers secondaires. Exemples :
|
|
2. Les principaux types de récepteurs A. Les récepteurs membranaires a. Les récepteurs métabotropes Ce sont des protéines trans-membranaires qui, après avoir fixé un ligand, changent de conformation et activent une cascade d'événements intracellulaires. Des récepteurs métabotropes ont été identifiés chez les animaux et les plantes. Il en existe plusieurs classes :
Il existe 3 grands types de récepteurs couplés à une enzyme intrinsèque :
|
|
Ce sont des protéines trans-membranaires qui, après avoir fixé un messager chimique, ouvrent un canal ionique : ces récepteurs - canaux sont généralement caractéristiques d'un ion (Na+, K+, Ca2+, Cl-). Il existe plusieurs classes de récepteurs ionotropes : α. Les récepteurs trimériques activés par l'ATP (récepteurs purinergiques) L'ATP est un intermédiaire dans la communication cellulaire (exemple : la transmission synaptique entre neurones). C'est un nucléotide qui peut être relargué en grandes quantités par des cellules en stress (inflammation, trauma, hypoxie). Dans ce cas, L’ATP est un signal de danger qui indique au système immunitaire que des tissus sont détruits. L’ATP extracellulaire est reconnu par des récepteurs purinergiques dont il existe 2 types :
Les récepteurs - canaux ioniques P2X contrôlent de manière non-sélective le passage de cations (Na+, K+ et Ca2+) au travers de la membrane plasmique de la cellule.
Source : Jiang et al. (2011) La fixation réversible de l'ATP sur le récepteur induit un changement de conformation du récepteur : ce canal ionique trans-membranaire est alors ouvert ce qui entraîne un influx de [Ca2+ / Na+] et un efflux de K+. Les récepteurs ATP2X sont structurés en 3 sous-unités homologues. Chaque sous-unité contient un domaine amino terminal extra-cellulaire, suivi par 2 segments transmembranaires séparés par un domaine extra-cellulaire. |
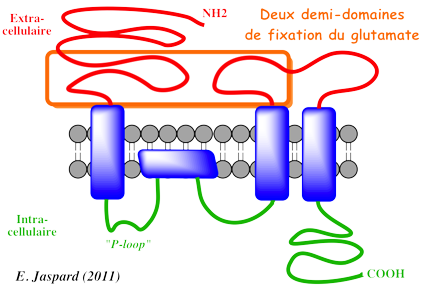
β. Les récepteurs tetramériques activés par le glutamate Ils sont spécifiques des cations. Ils sont structurés en 4 sous-unités homologues. Chaque sous-unité contient :
Exemple : le récepteur N-méthyl-D-aspartate (NMDA)
|
|
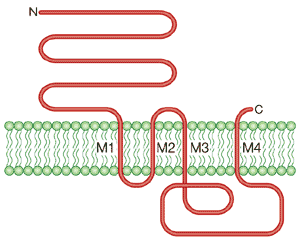
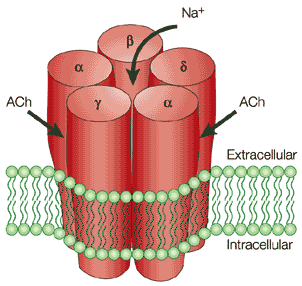
γ. Les récepteurs pentamériques ou "Cys-loop" Ce type de récepteurs constitue une superfamille ("Cys-loop ligand-gated ion channel superfamily"). Ils sont structurés en 5 sous-unités homologues. Exemple : le récepteur nicotinique de l'acétylcholine (figure ci-dessous).
Source : Karlin A. (2002) Chaque sous-unité contient 1 domaine amino terminal extra-cellulaire, 4 segments transmembranaires (M1 à M4) et la boucle localisée entre M3 and M4 constitue le domaine intracellulaire, de taille variable. Les récepteurs pentamériques ou "Cys-loop" peuvent être des canaux cationiques (exemples : le récepteur nicotinique, les récepteurs 5-HT3) ou des canaux anioniques (exemples : le récepteur GABA, le récepteur glycine). Figure ci-dessous : représentation de la structure quaternaire du récepteur nicotinique de l'acétylcholine dans laquelle on voit :
Source : Karlin A. (2002)
|
|
Des petites molécules lipophiles peuvent traverser la membrane plasmique ou entrer dans le noyau (via les pores nucléaires) et donc entrer dans les cellules. Les médiateurs des effets de ces petites molécules lipophiles sont les récepteurs nucléaires : ce sont dans la majorité des cas des facteurs de transcription activés par un ligand (les petites molécules lipophiles), qui régulent la transcription de réseaux complexes de gènes. Ces petites molécules lipophiles contrôlent donc indirectement de nombreux processus biologiques (reproduction, développement, métabolisme, inflammation, fonctions immunitaires, ...) chez les Eucaryotes. |
|
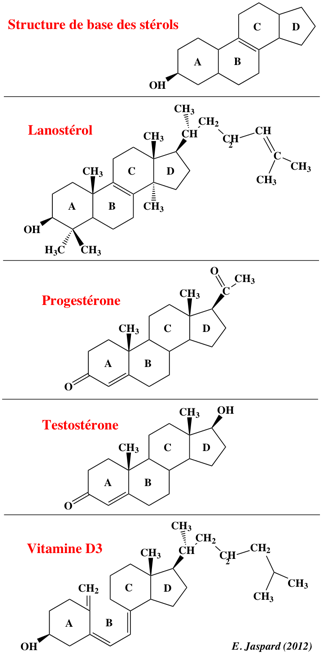
Ces petites molécules lipophiles sont essentiellement :
Voir une liste très complètes des récepteurs nucléaires connus chez l'homme. Certains récepteurs nucléaires n'ont pas de ligand identifié : ils sont dit "orphelins". |
|
α. Rôle et mode d'action des récepteurs nucléaires Il en existe plus de 150 repertoriés, dont 48 connus chez l'homme. Voir la base de données des récepteurs nucléaires : NURSA - Nuclear Receptor Signaling Atlas Un grand nombre de récepteurs nucléaires ont un rôle de facteur de transcription : ce sont des protéines qui agissent dans le noyau (en relayant des signaux hormonaux) et qui modulent la transcription des gènes. Ces récepteurs agissent en trans : ils induisent la transcription de gènes codant des protéines qui à leur tour activent de nombreux autres gènes. Certains récepteurs nucléaires ont un autre rôle en agissant sur d'autres voies de signalisation intracellulaire. Les récepteurs nucléaires peuvent être classés en 2 principales catégories selon leur mécanisme d'action et leur distribution sub-cellulaire en absence de leur ligand. Les récepteurs nucléaires de type I situés dans le cytosol puis délocalisés dans le noyau :
Les récepteurs nucléaires de type II :
Il existe 2 autres catégories "mineures" de récepteurs nucléaires :
|
|
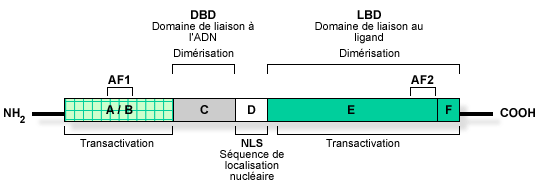
β. Structure des récepteurs nucléaires Ils ont une structure globale conservée. Ils sont constitués de 2 domaines :
Source : "Activation des récepteurs nucléaires"
Une fois activés, ces récepteurs se fixent sur l'ADN sous forme d'homo- ou d'hétérodimères. Les récepteurs nucléaires peuvent être classés en 4 catégories (voir ci-dessus) en fonction du type de dimérisation et des séquences d'ADN reconnues. Les acides aminés impliqués dans la dimérisation des récepteurs se trouvent dans les domaines C et [E / F]. La séquence en acides aminés appelée signal de localisation nucléaire ("Nuclear Localisation Signal" - NLS) permet au récepteur d'être adressé au noyau et d'y rester. |
|
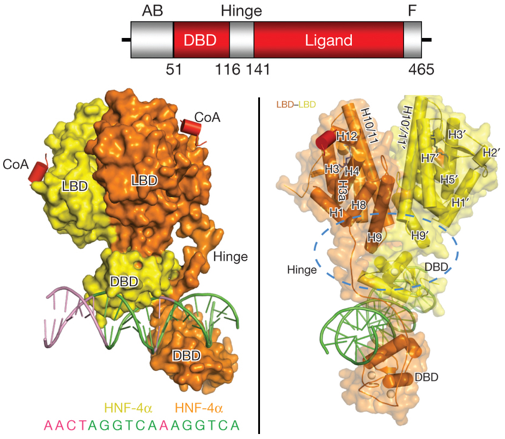
Illustration du récepteur nucléaire - facteur de transcription "Hepatocyte Nuclear Factor 4α" (HNF-4α ou NR2A1) HNF-4α est la protéine se fixant à l'ADN la plus abondante dans le foie où environ 40% des gènes transcrits possèdent un élément de réponse à HNF-4α. Ces gènes sont impliqués en grande partie dans la néoglucogénèse et le métabolisme des lipides. Les mutations de HNF-4α sont donc liées à l'hypoglycèmie hyperinsulinèmique et au diabète de type 1. Figure ci-dessous : structure cristalline (résolution 2.9 Å) des multiples domaines de HNF-4α humain (sous forme d'homodimère) fixé à son élément de réponse et à des peptides co-activateurs.
Source : Chandra et al. (2013) Une arginine cible de la méthylation par PRMT1 et une serine cible de la protéine kinase C contribuent au maintien des interactions entre domaines. Ces modifications post-traductionnelles induisent un changement de la fixation de l'ADN. |
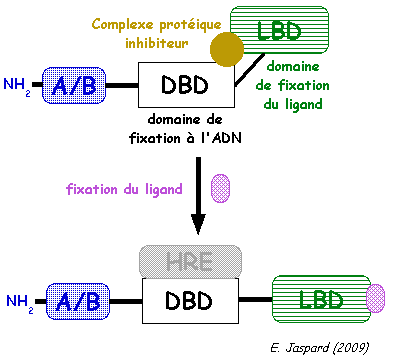
γ. Mode d'activation des récepteurs nucléaires Tant que le récepteur n'a pas fixé le ligand, il est dans une conformation inactive car le domaine DBD est bloqué par un complexe protéique inhibiteur.
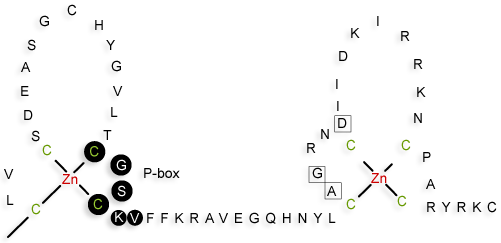
Les domaines de fixation de l'ADN des facteurs de transcription adoptent plusieurs types de structure très conservées. Par exemple, certains domaines DBD ont des séquences d'environ 66 à 68 acides aminés qui contiennent 9 cystéines conservées. Ces domaines DBD adoptent une structure dite "en doigt de zinc" de type C4 (4 cystéines). Cette structure est très rigide, et impose une topologie spécifique aux domaines DBD, qui permet à une hélice α de se positionner dans le grand sillon de l'ADN. Figure ci-dessous : les acides aminés de la "P-box" (cercles noirs) détermine la spécificité de reconnaissance de la séquence d'ADN. Les acides aminés entourés d'un carré sont impliqués dans la dimérisation du récepteur.
Source : "Activation des récepteurs nucléaires" |
|
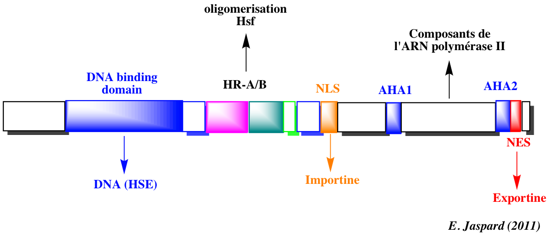
δ. Exemples de familles de facteurs de transcription Famille bZIP ("basic leucine zipper") : leur nom vient du fait qu'ils contiennent un domaine de fixation à l'ADN mixte constitué d' une séquence en acides aminés basiques et d'un motif riche en leucine à intervalles réguliers ("leucine zipper"). Les facteurs de transcription GBF1, GBF2 et GBF3 ("Arabidopsis bZlP family of G-box binding factors") intéragissent avec le motif palindrome "G-box" (CCACGTGG) trouvés dans de nombreux promoteurs de plantes. Famille HSF : ("heat shock transcription factor") : ils sont trimériques avec un domaine de fixation à l'ADN qui reconnaît la séquence répétée (nGAAn) et un domaine impliqué dans l'oligomérisation. Ils sont impliqués dans la réponse dite "de choc thermique" et la synthèse d'HSP ("heat shock proteins"). Il existe 3 classes (A, B et C). Ces facteurs de transcription sont organisés en modules fonctionnels comme le facteur de transcription AtHsfA2 de Arabidopsis :
Famille WRKY : ils sont ainsi nommés parce qu'ils possèdent un domaine de fixation à l'ADN (du côté N-terminal) qui contient, une ou deux fois, la séquence en acides aminés (quasi invariante) WRKY. Ils sont classés en fonction du nombre de motif WRKY et de leur motif "zinc-finger-like" : Cx[4,5]Cx[22,23]HxH ou Cx7Cx23HxC. |
|
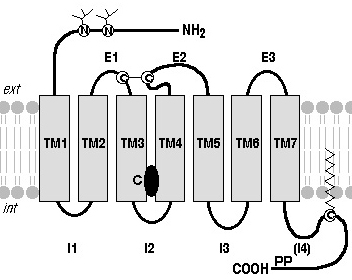
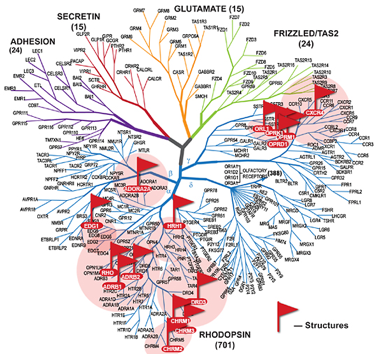
3. Les récepteurs couplés aux protéines G (RCPG) Robert Lefkowitz et Brian Kobilka ont reçu le prix Nobel de Chimie en 2012 pour leurs travaux sur les RCPG. Actuellement, on recense 826 RCPG chez l'homme. Les séquences codantes de leur gènes représentent plus de 1% du génome (voir Fredriksson et al., 2003). La structure de la grande majorité des RCPG est caractérisée par :
Source : "Récepteurs couplés aux protéines G"
|
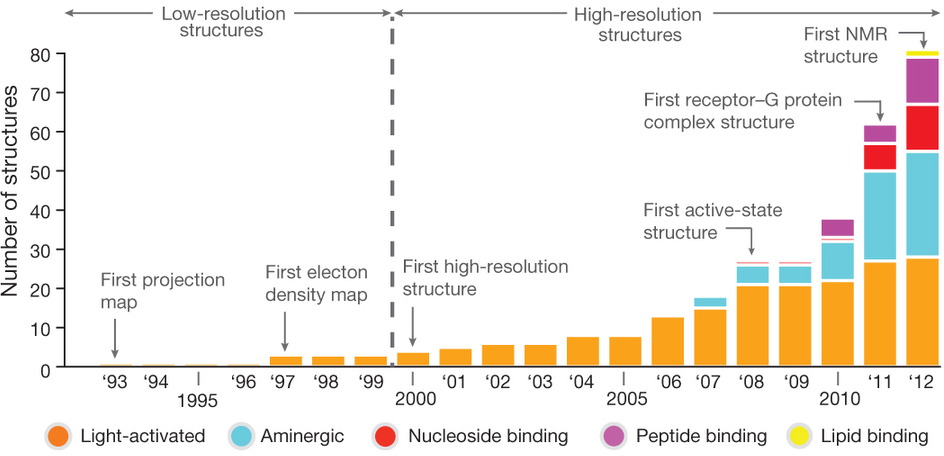
Figure ci-dessous : évolution du nombre et de la qualité des structures de RCPG déterminées par différentes techniques.
Source : Venkatakrishnan et al. (2013) L'obtention d'un nombre croissant de structures de RCPG permet d'affiner le mode de régulation de leur activation : elle est contrôlée par des mécanismes allostériques liés à la fixation de leur ligand mais aussi à la fixation d'un ion Na+, de lipides, du cholestérol et de molécules d'eau. La structure (résolution 1.8 Å) du récepteur de l'adénosine A2A a démontré la présence d'un groupe [Na+/molécules d'eau] au milieu du faisceau des 7 hélices transmembranaires. L'ion Na+ établit des liaisons de coordination avec les acides aminés conservés D2.50 et S3.39 (numérotation selon la nomenclature Ballesteros - Weinstein) et 3 molécules d'eau. Voir le mécanisme d'auto-activation du récepteur de la thrombine, impliqué dans la coagulation du sang. |
| Classe de RCPG | Famille de RCPG | Exemple de ligands |
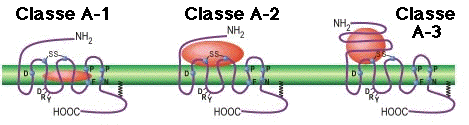
Classe A ("Rhodopsin like receptor family"). Ce sont de loin les plus nombreux : à ce jour plus de 700 gènes identifiés chez l'homme. Motifs fortement conservés dans certaines hélices transmembranaires : D(E)RY - TM2 / FxxCWxP - TM6 / NPxxY - TM7. Voir une analyse des relation phylogénétiques des sous-classes A (Pelé et al., 2011) |
||
| Classe A / sous-type 1 | Famille 1 / sous-groupe a | Rétinal, odorant, opiacés, β2-adrénergiques, dopaminergiques, histaminiques |
| Classe A / sous-type 2 | Famille 1 / sous-groupe b | bradykinine, angiotensine II, cytokines, thrombine |
| Classe A / sous-type 3 | Famille 1 / sous-groupe c | Hormones, glycoprotéiques |
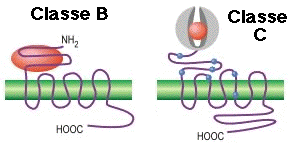
| Classe B ("Secretin like receptor family") | Famille 2 | Calcitonine, sécrétine |
| Classe C ("Metabotropic glutamate/pheromone receptor family") | Famille 3 | Glutamate, acide γ-aminobutyrique (GABA), calcium, phéromones |
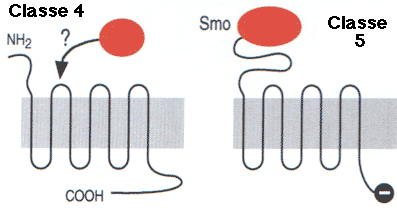
| Classe F ("Frizzled") | Familles 4 et 5 | Phéromones / développement embryonnaire |
| Les liens ci-dessus renvoient vers la base de données dédiée aux RCPG - GPCRdb. | ||
|
Source : S. Martin |
Source : P. Sarret |
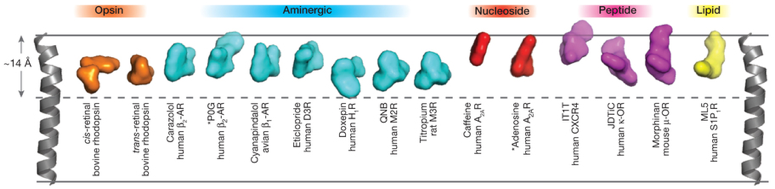
Figure ci-dessous : comparaison de la profondeur des poches de fixation des ligands des RCPG de la classe A. L'hélice transmembranaire TM4 est utilisée comme référence. Les étiquettes au dessus des structures indiquent la sous-classe des RCPG de la classe A.
Source : Venkatakrishnan et al. (2013) La profondeur de pénétration du ligand est la plus grande pour la doxépine pour le récepteur de l'histamine H1 et la moins grande pour la caféine pour le récepteur A2AR. |
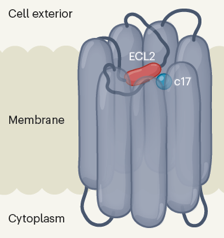
RCPG "orphelins" - Couples [RCPG - métabolites] Au début des années 2000, on a découvert que des métabolites du métabolisme énergétique sont les ligands de certains RCPG (interaction de faible affinité - RCPG 35, 40, 41, 43, 84, 91, 99, 120, ... chez l'homme) : lactate, succinate, β-hydroxybutyrate, acide kynurénique (acide lysophosphatidique), acide 3-hydroxy-octanoique , courtes, moyennes et longues chaînes d'acides gras libres, acides biliaires ... Ces récepteurs agissent comme des senseurs de l'activité métabolique ou du niveau énergétique de certains métabolites et utilisent cette information pour ajuster la sécretion d'hormones ou pour réguler l'activité métabolique de certaines cellules. Voir une revue : Smith N.J. (2012). Illustration du RCPG orphelin GPR52 auto-activant Le RCPG GPR52 est une cible potentielle pour le traitement de troubles neuropsychiatriques (par exemple, la maladie de Huntington et la schizophrénie). Lorsqu'il est activé (fixation de la réserpine par exemple), GPR52 se lie sélectivement à la famille Gs des protéines G (voir ci-dessous) et stimule la synthèse d'AMPc. GPR52 est le premier RCPG orphelin dont on ait déterminé la structure 3D (Lin et al., 2020) :
Source : Krumm & Roth (2020) Par ailleurs, le système cyclique aromatique de la molécule synthétique c17 (3-méthyle-5-hydroxyméthyle-1H-pyrazole, agoniste de substitution qui active GPR52) se lie (interactions π - π avec Phe 3007.36) à une région proche du site auquel se fixe ECL2. La molécule c17 pourrait donc être un modulateur allostérique qui potentialise l'activité du récepteur sans se lier au site de liaison de l'agoniste.
|
Source : Lin et al. (2013) |
Source : GPCR network |
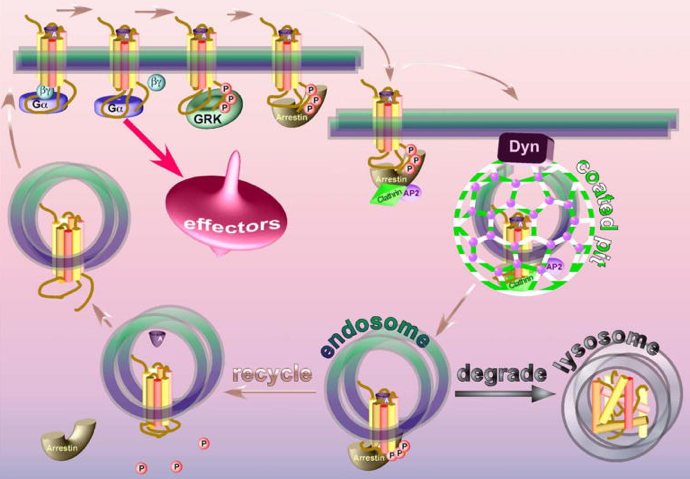
d. Traffic intracellulaire et recyclage des RCPG La phosphorylation de résidus Ser ou Thr de l'extrémité C-terminale par les GRK ("G-protein-coupled receptor kinases"), joue un rôle capital dans le traffic intracellulaire et le recyclage des RCPG. 1ère étape : cette phosphorylation augmente la fixation de protéines appelées β-arrestines sur la surface intracellulaire du récepteur. En masquant cette surface ("scaffolding proteins"), les β-arrestines empêchent ainsi le couplage du RCPG avec sa protéine G partenaire. C'est la désensibilisation ("desensitization") du RCPG.
Source : Gurevitch & Gurevitch (2006) 2ème étape : les β-arrestines fixent le RCPG aux éléments de la machinerie d'internalisation, la clathrine et l'adaptateur de la clathrine AP2 ("coated pit"). Il y a endocytose du RCPG (endosomes). 3ème étape : le RCPG est alors
|
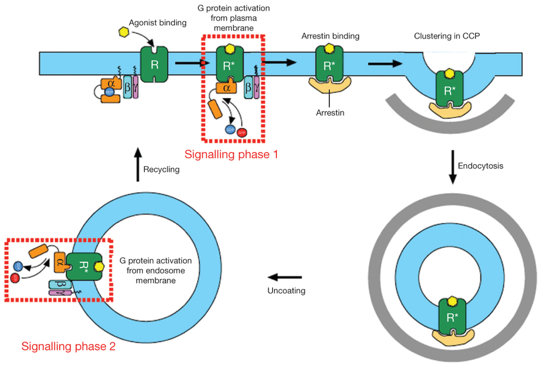
e. Voie non conventionnelle de la signalisation par les RCPG : "2ème vague" d'activation via les endosomes La voie classique ou principale de la transduction du signal médié par les RCPG est celle qui met en jeu les RCPG liés à la membrane plasmique.
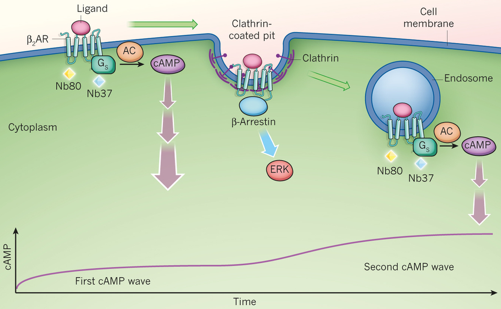
Source : Irannejad et al. (2013) La difficulté majeure pour étayer la ou les voie(s) moins conventionnelle(s), après internalisation de RCPG dans des endosomes, est l'absence de méthodes d'analyse qui fournissent des preuves expérimentales à l'échelle sub-cellulaire. En d'autres termes, il est difficile de prouver que les RCPG localisés dans les endosomes sont encore actifs. Une preuve expérimentale qui renforce cette hypothèse a récemment été apportée par l'utilisation de nano-anticorps ("nanobodies" - "conformation-specific single-domain antibodies"), qui sont les plus petits fragments d'anticorps entièrement fonctionnels, naturellement produits chez le chameau. Voir le cours : "Compléments sur les récepteurs couplés aux RCPG". Du fait de leur petite taille (15 kDa) et de leur flexibilité, ils accèdent à des régions de protéines que les anticorps ne peuvent pas atteindre. Ils ont notamment été employés pour stabiliser les formes actives des couples [RCPG - protéine G] afin de les cristalliser (Steyaert & Kobilka, 2011) . Cette technique a été employée pour sonder directement l'activation du récepteur β2-adrénergique (β2AR) associé à une protéine G de la classe Gs qui stimulent la production de l'AMPc dans des cellules de mammifère. Deux nano-anticorps ont été utilisés : Nb80 qui se fixe à β2AR et Nb37 qui se fixe à la forme Gs dépourvue du nucléotide. |
|
Modèle décrivant 2 phases d'activation de [β2AR - Gs] 1ère phase : au niveau de la membrane plasmique qui résulte en un premier accroissement du taux d'AMPc. il y aurait ensuite formation d'invaginations (recouvertes de clathrine) de la membrane : interaction du récepteur avec la β-arrestine, activation d'enzyme ERK ("Extracellular signal-Regulated Kinase") et autres signaux non conventionnels. 2ème phase : après internalisation du récepteur dans les vésicules sous forme d'endosomes. C'est la "2ème vague" impliquant la protéine Gs, qui résulte en un second accroissement du taux d'AMPc.
Source : Lohse & Calebiro (2013) L'agoniste adrénergique isoprénaline active le récepteur et la protéine G dans la membrane plasmique, comme prévu, mais aussi dans la membrane de l'endosome précoce. Les récepteurs internalisés dans des endosomes contribuent à la réponse AMPc globale quelques minutes après application de l'agoniste. |
|
Elles ont été découvertes par Alfred G. Gilman et Martin Rodbell (prix Nobel de médecine 1994). Les protéines G (G pour "Guanine nucleotide binding proteins") participent à la transduction du signal : ce sont des intermédiaires entre les récepteurs d'hormones situés dans la membrane plasmique et les systèmes réactionnels intracellulaires. Les protéines G appartiennent à la famille des enzymes GTPases (E.C. 3.6.5) qui hydrolysent la guanosine triphosphate (GTP). La famille des GTPases contient des petites protéines G monomèriques qui participent à d'autres processus cellulaires : la synthèse des protéines, la division cellulaire, la translocation des protéines au travers de la membrane, le transport des vésicules dans la cellule. Les protéines G amplifient le signal :
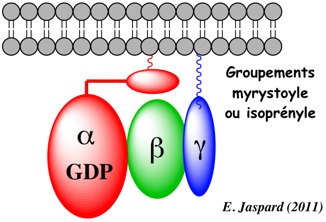
Les protéines G sont des protéines hétérotrimétriques liées à la membrane, constituées d'une sous-unités alpha (39 - 46 kDa), beta (35 - 39 kDa) et gamma (8 kDa).
Deux sous-unités sont directement liées à la membrane : l'extrémité N-terminale de la sous-unité α et l'extrémité C-terminale de la sous-unité γ sont modifiées par des groupements lipidiques myristoyle et isoprényle. |
Visualisation de la protéine G hétérotrimèrique complexée au GDP à une résolution de 2,4 Å Code PDB : 1GG2 Remarques :
|
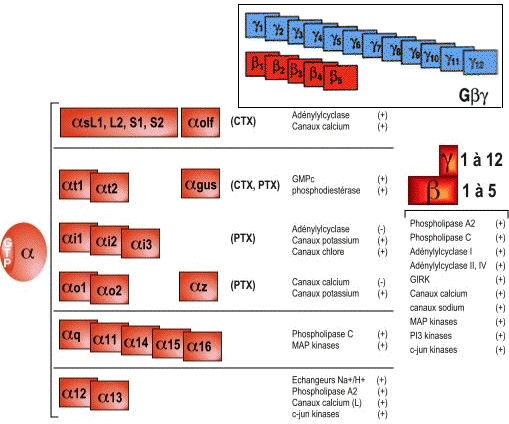
a. Diversité des sous-unités des protéines G La diversité des protéines G est moindre que celle des RCPG, mais permet de nombreuses combinaisons (environ 700 potentielles) entre les sous-unités Gα et [Gβ/Gγ]. En effet, il existe au moins (figure ci-dessous) : 16 gènes codant une sous-unité Gα, 5 gènes codant une sous-unité Gβ et 12 gènes codant une sous-unité Gγ. |
Les diverses classes de sous unités α :
Voir une revue sur l'action de ces différents types de sous unités et celles des sous-unités [β - γ]. |
Source : S. Martin |
|
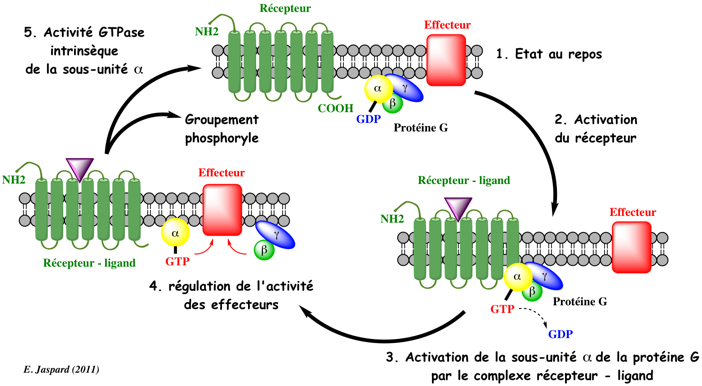
b. Activation - désactivation des protéines G La structure complète d'un complexe RCPG - protéine Gs (PDB : 3SN6) a été publiée en 2011 par l'équipe de B. Kobilka (Prix Nobel 2012) : Rasmussen et al. (2011) "Crystal structure of the β2 adrenergic receptor–Gs protein complex" Nature 477, 549 - 555 Le mécanisme est le suivant : quand une hormone se fixe à son récepteur sur la face externe de la membrane plasmique, celui-ci change de conformation (figure ci-dessous). Le récepteur peut alors se fixer à une protéine G (inactive - chargée en GDP) située sur la face interne de la membrane plasmique.
Une fois activée, la sous-unité α se dissocie du récepteur et se fixe à sa protéine cible appelée effecteur en l'activant.
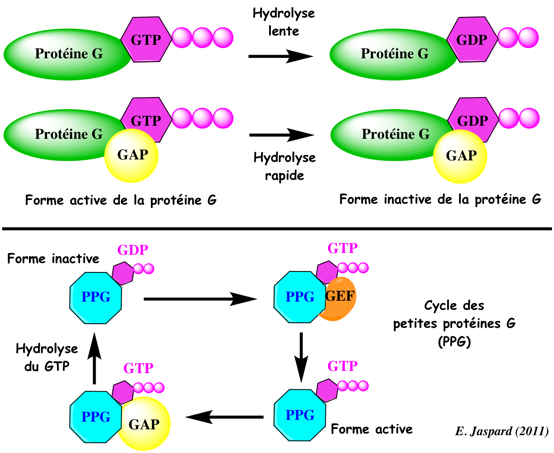
L'activité GTPase de la sous-unité α hydrolyse le GTP ce qui entraîne la réassociation des 3 types de sous-unités de la protéine G. La protéine G retrouve sa conformation liant le GDP et elle est de nouveau inactive. L'activité d'hydrolyse des groupements phosphates des protéines G est faible. Les protéines GAP ("GTPase-Activating Proteins" ou " GTPase-Accelerating Proteins") stimulent l'activité GTPase de la sous-unité α. Elles contribuent donc à la terminaison du cycle en accélérant l'hydrolyse du GTP (figure du haut ci-dessous).
Il existe des petites protéines G monomèriques (exemples : Ras, Rho, Rab, Ran, Arf, ...). Elles sont impliquées dans la régulation du cycle cellulaire, le transport, la polarité des cellules pendant leur développement, ... Le cycle GDP/GTP des petites protéines G monomèriques est régulé par :
|
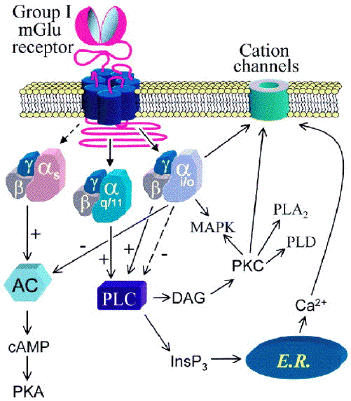
c. 1er facteur de la multiplicité des voies de signalisation Le couplage du récepteur à plusieurs sous-types de protéines G différentes. Le glutamate est un neurotransmetteur excitateur majeur dans les cellules du systéme nerveux central des mammifères. Son action est médiée par un ensemble de récepteurs métabotropes (8 récepteurs "mGlu" classés en 3 groupes ) et ionotropes ("glutamate-gated ion channels" - NMDA, AMPA, récepteurs kaïnate). Ces récepteurs couplent l'action de protéines G à différents messagers secondaires :
Figure ci-dessous : les multiples voies de signalisation activées par le récepteur mGlu1a via son con couplage à différents types de protéines G.
Source : Hermans & Challiss (2001) Légende : AC : adénylate cyclase - cAMP : AMP cyclique - PKA : protéine kinase A - PLC : phospholipase C - InsP3 : inositol 1,4,5-triphosphate - DAG : 1,2-diacylglycérol - PKC : protéine kinase C - MAPK : "mitogen-activated protein kinase" - E.R. : réticulum endoplasmique |
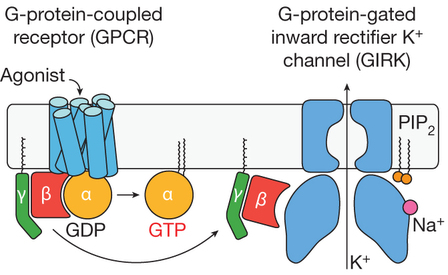
d. 2ème facteur de la multiplicité des voies de signalisation : les sous-unités [β - γ] Les sous-unités [β - γ] peuvent agir sur des effecteurs différents de ceux reconnus par la sous-unité α, après l'activation et la dissociation des sous-unités. Les sous-unités β et γ ne sont pas des enzymes, elles n'ont donc pas de site catalytique : elles agissent donc comme des modulateurs dans la signalisation liée aux protéines G via des interactions protéine - protéine qu'elles régulent. En conséquence, le complexe [β - γ] agit aussi comme une molécule signal. Voir une liste de quelques cibles des sous-unités [β - γ]. C'est le cas par exemple du récepteur [muscarinique / acétylcholine] impliqué dans l'ouverture des canaux à courant potassique GIRK ("G protein-regulated inward rectifier potassium channel").
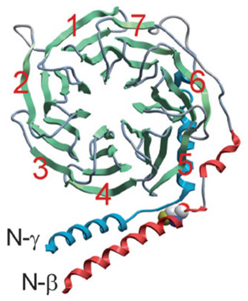
Source : Whorton & MacKinnon (2013) On dénombre 5 gènes codant la sous-unité β et 12 gènes codant la sous-unité γ chez l'homme et la souris. Il y a une plus grande identité de séquences en acides aminés pour les sous-unités β (80% entre β1 et β4) que pour la sous-unité γ. Ces différentes isoformes de sous-unités β et γ peuvent s'assembler en différentes combinaisons [βx/γy]. Figure ci-dessous : structure des sous-unités β et γ du dimère β1/γ1 (PDB 1TBG). Les numéros correspondent aux différents parties des hélices (en rouge : hélice N-terminale de la sous-unité β / en bleu : hélice N-terminale de la sous-unité γ).
Source : Wall et al. (1995) Le complexe [β/γ ] est membranaire : l'extrémité C-terminale de la sous-unité γ est isoprénylée par des groupements farnesyle ou géranyle-géranyle liés à une Cys. Ainsi modifié, le complexe [β/γ ] est adressé au réticulum endoplasmique où une protéase ("Ras converting enzyme" - Rce1) hydrolyse les 3 acides aminés C-terminaux de la sous-unité γ. Enfin l'isoprényle cystéine carboxy-méthyl transférase catalyse la carboxy-méthylation de l'acide aminé C-terminal de la sous-unité γ et celle-ci est pleinement modifiée post-traductionnellement. |
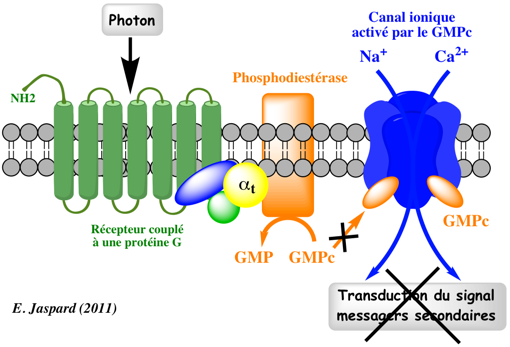
5. Exemples de transduction de signaux par des récepteurs couplés aux protéines G Dans le système de la vision, la rhodopsine (pour les bâtonnets) et 3 opsines distinctes (pour les cônes sensibles aux couleurs) servent de récepteurs pour la lumière. La molécule qui absorbe le photon est le 11-cis-rétinal lié de manière covalente à une lysine dans la 7è hélice trans-membranaire. Le récepteur s'associe à une protéine G appelée transducine (αt1, αt2) qui active la phosphodiéstérase (PDE). Or la PDE hydrolyse le GMP cyclique (GMPc) et la baisse de concentration de GMPc provoque la fermeture d'un canal ionique et l'hyperpolarisation de la cellule. Des mutations affectant les gènes codant pour les opsines conduisent à ne plus pouvoir distinguer les couleurs (daltonisme).
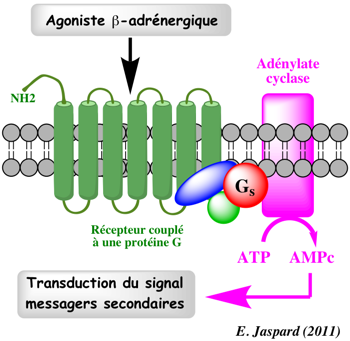
Le récepteur β-adrénergique s'associe avec la protéine Gs qui le couple à l'adénylate cyclase (AC). L'AC produit l'AMP cyclique (AMPc) qui exerce ses effets par l'intermédiaire d'une protéine kinase AMPc dépendante.
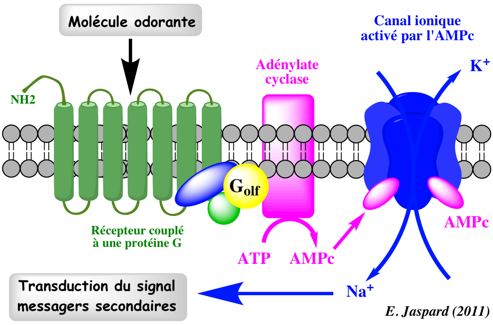
Dans le système olfactif, les récepteurs des odeurs s'associent à une unique protéine G appelée Golf. Cette protéine G couple le récepteur à une forme d'adénylate cyclase propre aux neurones olfactifs. La formation d'AMPc ouvre un canal ionique Na+ - K+.
|
|
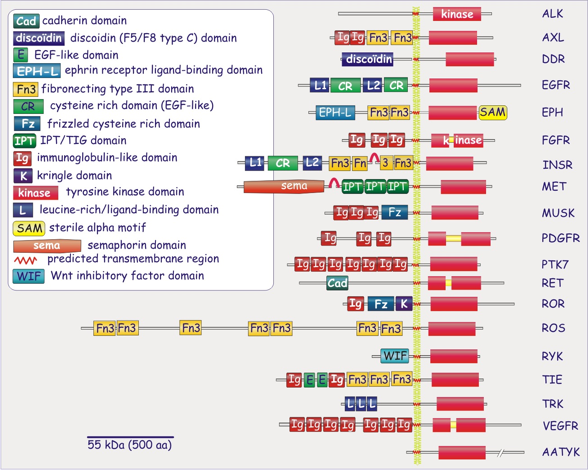
a. Généralités Il a été découvert dans les années 70 [Freychet et al. (1971) Proc Natl Acad Sci 68, 1833]. Le récepteur de l'insuline joue un rôle central dans l'homéostasie du glucose. Il fait partie de la super-famille des récepteurs à activité tyrosine kinase, famille subdivisée en une vingtaine de classes (59 récepteurs de ce type codés par le génome de l'homme). Les récepteurs à activité tyrosine kinase médient le signal perçu en phosphorylant des tyrosines des protéines cibles. Parmi les autres membres de cette famille, on peut citer "insulin-like growth factor receptor" (IGF1R) et "insulin receptor related receptor" (IRR).
Source : "Cell Biology promotion" - INSR = récepteur de l'insuline; VEGFR" = "Vascular Endothelial cell Growth Factor Receptors". La teneur en récepteur de l'insuline est variable. La concentration la plus élevé est dans les cellules impliquées dans le métabolisme du glucose, des lipides et des protéines sous contrôle de l'insuline : les tissus adipeux, les muscles squelettiques et le foie. |
b. Maturation du récepteur de l'insuline La forme non maturée, le pré-pro-récepteur, est une séquence linéaire α - β. La séquence signal de 27 acides aminés hydrophobes en position N-terminale adresse le pré-pro-récépteur au réticulum endoplasmique au sein duquel le peptide signal est hydrolysé. Le pro-récépteur est ensuite protéolysé par la furine (ou "Paired basic Amino acid Cleaving Enzyme" - PACE) dans l'appareil de Golgi en 2 chaînes polypeptidiques distinctes α et β, après que les deux sous-unités aient été reliées par des ponts disulfure. La furine est une protéase à sérine calcium-dépendante de la famille de la subtilisine. Le site de protéolyse est constitué de 4 acides aminés basiques (Arg-Lys-Arg-Arg). Chaque dimère est donc constitué d'une sous-unité α (partie N-terminale) et d'une sous-unité β transmembranaire et intracellulaire. Le récepteur de l'insuline mature est une glycoprotéine sous forme de dimère de dimère α2β2 (structure PDB 2DTG). Remarque : l'insuline elle-même est une protéine biosynthétisée sous forme de pré-pro-insuline qui subit de profondes modifications post-traductionnelles pour aboutir à sa forme mature fonctionnelle. |
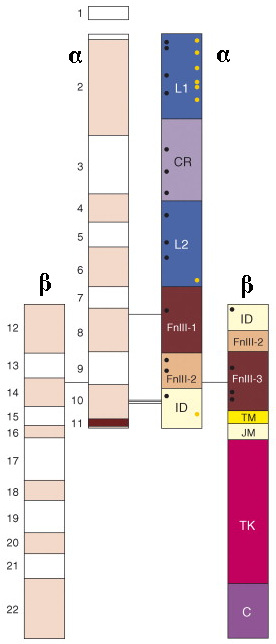
c. Structure du récepteur α2β2 Cette structure est particulièrement complexe (figure et description ci-dessous).
Source : De Meyts (2008)
Remarque : on trouve d'autres nomenclatures dans la littérature, mais l'ordre est bien évidemment le même. La sous-unité α extracellulaire
La sous-unité β transmembranaire et intracellulaire Le domaine transmembranaire (riche en hélice) est suivi par un domaine adjacent à la membrane. Le domaine tyrosine kinase et la queue C-terminale sont intracellulaires. Les ponts disulfures
|
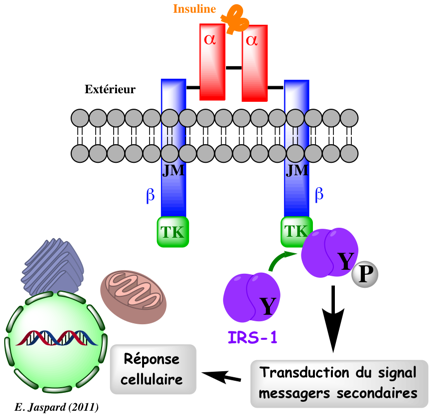
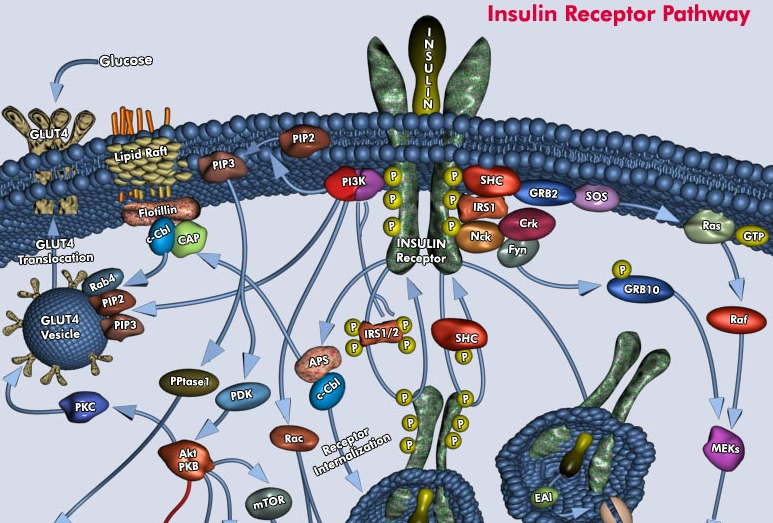
d. Exemples de voies de signalisation liées au récepteur de l'insuline La fixation de l'insuline sur le récepteur de l'insuline induit une autophosphorylation de plusieurs tyrosines de la sous-unité β intracellulaire : tyrosines 965 et 972 (domaine JM), tyrosines 1158, 1162 et 1163 (domaine TK), tyrosines 1328 et 1334 (domaine C). Cette autophosphorylation induit des changements de conformation qui, à leur tour, induisent l'activation de l'activité tyrosine kinase. Le récepteur de l'insuline peut alors phosphoryler différentes protéines cibles effectrices. Bien que liées, l'autophosphorylation et l'activation du domaine tyrosine kinase sont deux aspects distincts du fonctionnement du récepteur de l'insuline : ils peuvent être découplés et sont soumis à différents mécanismes de régulation.
Parmi les protéines cibles, on peut citer "insulin receptor substrate" (IRS-1 et IRS-2), "Src homology collagen" (Shc) et APS ("adaptor protein" avec un domaine SH2). L'activité tyrosine kinase du récepteur de l'insuline stimule, entre autres, l'activité kinase de la phosphatidylinositol 3-kinase (PI3K) . L'augmentation rapide de la concentration de l'inositol triphosphorylé (PIP3) déclenche une cascade de Ser/Thr protéines kinases PIP3-dépendantes.
Source : Qiagen Parmi ces dernières, Akt (ou Ser/Thr protéine kinase B) et des isoformes atypiques de la protéine kinase C semblent impliquées dans :
L'existence d'un grand nombre de [Ser/Thr protéines kinases PIP3-dépendantes / insulino-dépendantes] renforce l'hypothèse que différentes voies métaboliques sont nécessaires pour réguler les différentes actions biologiques de l'insuline. La déphosphorylation du récepteur de l'insuline par les protéines tyrosine phosphatases (PTP) est responsable de la désactivation de son activité tyrosine kinase. Exemples de PTP dans le muscle squelettique : les "leukocyte common antigen-related phosphatases" (LAR) et la PTP-1B. |
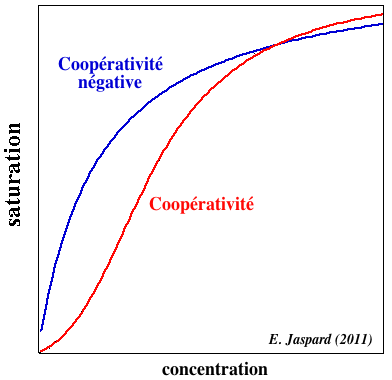
e. Coopérativité négative de la fixation de l'insuline à son récepteur Selon les études et/ ou les modèles théoriques, 1 ou 2 molécules d'insuline se fixe(nt) par récepteur.
La coopérativité négative semble liée à certaines protéines qui sont des points de branchement dans le réseau de régulation métabolique : ce concept s'applique à certains couples [ligand - récepteur] comme le couple [insuline - récepteur de l'insuline]. Par exemple, la cytidine triphosphate (CTP) est à la fois le produit final d'une série de réactions dont le point de départ est catalysé par la carbamyl-phosphate synthétase et un inhibiteur allostérique négatif de cette enzyme. Or cette enzyme est un carrefour à partir duquel un grand nombre de métabolites autres que la CTP sont synthétisés. Il n'est pas concevable qu'une inhibition de cette enzyme par la CTP soit totale, car ce serait au détriment de la synthèse des autres métabolites. La coopérativité négative de la fixation du CTP permet donc que, même en trés large excès, l'inhibition de la carbamyl-phosphate synthétase ne soit pas totale. |
| 7. Liens Internet et références bibliographiques |
|
Base de données : "GPCRDB: Information system for G protein-coupled receptors (GPCRs)" Base de données : "Orientation of Proteins in Membranes" Base de données des récepteurs nucléaires : "The Nuclear Receptor Signaling Atlas" Nobel Prize in Chemistry 2012 : Robert J. Lefkowitz and Brian K. Kobilka |
|
|
Gether, U.(2000) "Uncovering Molecular Mechanisms Involved in Activation of G Protein-Coupled Receptors" Endocrine Rev. 21,90 - 113 Kido et al. (2001) "The Insulin Receptor and Its Cellular Targets" J. Clin. Endocrin. Metab. 86, 972 Fredriksson et al. (2003) "The G-Protein-Coupled Receptors in the Human Genome Form Five Main Families. Phylogenetic Analysis, Paralogon Groups, and Fingerprints" Mol. Pharmacol. 63, 1256 - 1272 J. Cherfils & P. Pacaud (2004) "L'activation des protéines G en 3 dimensions : un pas vers l'inhibition thérapeutique des facteurs d'échange nucléotidique" M/S : médecine sciences 20 De Meyts (2008) "The insulin receptor: a prototype for dimeric, allosteric membrane receptors ?" Trends Biochem. Sci. 33, 376 - 384 Jiang et al. (2011) "Agonist trapped in ATP-binding sites of the P2X2 receptor" PNAS 108, 9066 - 9071 |
|
|
"Kyoto Encyclopedia of Genes and Genomes" "Cell signaling" |
|
|
Koshland & Hamadani (2002) "Proteomics and Models for Enzyme Cooperativity" J. Biol. Chem. 277, 46841 - 46844 Chapitre 34 - "Biochemistry" - 3rd Edition (2005) Reginald H. Garrett & Charles M. Grisham - University of Virginia : "An interactive biochemistry book developped with the support of WWW . Web." Virginia. Ed. Wall et al. (1995) "The structure of the G protein heterotrimer Giα1β1γ2" Cell 83, 1047 - 1058 Prasobh & Manoj (2009) "The Repertoire of Heterotrimeric G Proteins and RGS Proteins in Ciona intestinalis" PLoS ONE 4, e7349 |
|
|
Ensemble de superbes cours (en Français) sur les RCPG et les protéines G Silva & Marcey : "An introduction to Jmol Scripting" |
|
|
Protéine G - Wikipédia Récepteurs - "Phamacorama" "Cell Biology promotion" : Des dizaines de trés beaux schémas de récepteurs avec la description de leur mécanisme. Wettschureck & Offermanns (2005) "Mammalian G Proteins and Their Cell Type Specific Functions" Physiol Rev 85, 1159 - 1204 |
|
|
Maruyama et al. (2002) "Identification of membrane-type receptor for bile acids (M-BAR)" Biochem. Biophys. Res. Commun. 298, 714 - 719 He et al. (2004) "Citric acid cycle intermediates as ligands for orphan G-protein-coupled receptors" Nature 429, 188 - 193 Kimura et al. (2011) "Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41)" Proc. Natl. Acad. Sci. U S A. 108, 8030 - 8035 Smith, NJ. (2012) "Low Affinity GPCRs for Metabolic Intermediates: Challenges for Pharmacologists" Front. Endocrinol. 3, 1 |
|
|
Hermans and Challiss (2001) "Structural, signalling and regulatory properties of the group I metabotropic glutamate receptors: prototypic family C G-protein-coupled receptors" Biochemical J. 359, 465 - 484 Karlin A. (2002) "Emerging structure of the Nicotinic Acetylcholine receptors" Nature Rev. Neuroscience 3, 102 - 114 Rasmussen et al. (2011) "Crystal structure of the β2 adrenergic receptor–Gs protein complex" Nature 477, 549 - 555 Chandra et al. (2013) "Multidomain integration in the structure of the HNF-4α nuclear receptor complex" Nature |
|
|
Gurevitch & Gurevitch (2006) "The structural basis of arrestin-mediated regulation of G-protein-coupled receptors" Pharmacol. Ther. 110, 465 - 502 Smrcka, AV. (2009) "G protein βγ subunits: Central mediators of G protein-coupled receptor signaling" Cell Mol. Life Sci. 65, 2191 - 2214 Pelé et al. (2011) "Multidimensional Scaling Reveals the Main Evolutionary Pathways of Class A G-Protein-Coupled Receptors" PLoS ONE 6(4): e19094 Zheng et al. (2012) "Palmitoylation and membrane cholesterol stabilize μ-opioid receptor homodimerization and G protein coupling" BMC Cell Biol. 13, 6 Lin et al. (2013) "A pharmacological organization of G protein–coupled receptors" Nature Methods 10, 140 - 146 |
|
|
Steyaert & Kobilka (2011) "Nanobody stabilization of G protein-coupled receptor conformational states" Curr. Opin. Struct. Biol. 21, 567 - 572 Irannejad et al. (2013) "Conformational biosensors reveal GPCR signalling from endosomes" Nature 495, 534 - 538 Lohse & Calebiro (2013) "Cell biology : receptor signals come in waves" Nature 495, 457 - 458 Venkatakrishnan et al. (2013) "Molecular signatures of G-protein-coupled receptors" Nature 494, 185 - 194 Whorton & MacKinnon (2013) "X-ray structure of the mammalian GIRK2–βγ G-protein complex" Nature 498, 190 - 197 |
|
|
Ballesteros & Weinstein (1995) "Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors" Meth. Neurosciences 25, 366 - 428 Isberg et al. (2015) "Generic GPCR Residue Numbers - Aligning Topology Maps Minding The Gaps" Trends Pharmacol. Sci. 36, 22 - 31 |
|
|
Krumm & Roth (2020) "A self-activating orphan receptor" Nature - News and views Lin et al. (2020) "Structural basis of ligand recognition and self-activation of orphan GPR52" Nature |
![]()